
Abstract
Doppel (Dpl) protein shares some structural features with prion protein (PrP), whose pathologic isoform (PrPsc) is considered to be the causative agent of transmissible spongiform encephalopathies. Dpl is mainly expressed in testes but, when ectopically expressed in the central nervous system, is neurotoxic. We have examined the expression pattern of Dpl and PrP on bovine lymphoid tissues and circulating leukocytes. A polyclonal anti-Dpl antibody along with a panel of monoclonal antibodies specific for leukocyte membrane antigens or PrP were used to examine frozen sections from spleen, lymph nodes, and bone marrow by immunohistochemistry. Blood was analyzed by flow cytometry. Double staining was used to study the possible coexpression of the two proteins and to characterize cells expressing Dpl and/or PrP. Dpl was expressed in B-cells, in dendritic cells within lymphoid follicles, bone marrow, circulating myeloid cells, and circulating B-cells. The distribution of Dpl was quite similar to that of PrP. The only differences in expression observed concerned the low number of Dpl + cells in lymph nodes and the strong Dpl positivity of circulating granulocytes. The two proteins were rarely co-expressed, suggesting an independent expression mechanism in resting cells. The role of Dpl+ leukocytes in the pathogenesis of Dpl- or PrP-induced diseases merits further investigation.
D
Studies using Western blot and RT-PCR have shown that the tissue distribution of Dpl in both humans and bovine is similar to that reported in mice (Sakaguchi et al. 1996; Moore et al. 1999; Tranulis et al. 2001; Rondena et al. 2004). Interestingly, bovine Dpl has also been visualized by immunohistochemistry on the membrane of scattered cells in the spleen and lymph nodes (Rondena et al. 2004). Previous studies have shown that PrP is also expressed in follicular dendritic cells (FDCs) (Thielen et al. 2001b) and in several types of blood cells in various species (Herrmann et al. 2001; Barclay et al. 2002). The expression of PrP in FDCs is essential for neuroinvasion (Brown et al. 1999; Mabbot et al. 2000), while the role of PrP-expressing leukocytes is still debated (Klein et al. 1998; Li et al. 2001).
Because of the above-mentioned similarities between PrP and Dpl and the possible role of Dpl in the pathogenesis of experimentally induced (PrP deletion) or spontaneous (infertility, brain tumor) diseases, it is important to understand more about Dpl biology. This article describes the results of a study focused on the expression of Dpl in both circulating and tissue leukocytes and on the relationship between Dpl and PrP expression. For these purposes, bovine cells and tissues were used owing to the relevance of cattle in prion-related diseases.
Materials and Methods
Biological Samples
Jejunal and tracheobronchial lymph nodes, spleen, and bone marrow were collected at the public slaughterhouse of Milan from 10 routinely slaughtered calves of both sexes aged 12-24 months. Tissue samples, including bone marrow obtained by vertebral squeezing, were immediately frozen in liquid nitrogen and stored at −80C. From each sample, 6-μm-thick cryostat sections were prepared and routinely stained with hematoxylin and eosin or used for immunohistochemistry.
Blood was collected by venipuncture from the coccygeal vein of six adult Holstein-Fresian cattle. Animals came from a dairy farm and were clinically healthy. Blood was collected in EDTA-coated tubes. A routine complete cell blood count using an automated analyzer (Hemat 8; SEAC, Firenze, Italy) followed by microscopic evaluation of differential leukocyte count on May-Grünwald-Giemsa-stained smears was performed to exclude the presence of subclinical hematological abnormalities. Blood was then used to perform flow cytometry.
Bovine semen was used as a positive control for flow cytometric detection of Dpl due to the established expression of the protein in spermatozoa (Rondena et al. 2004). Frozen bovine semen, routinely stored for bovine artificial insemination at the Department of Clinical Sciences of the Veterinary School of Milan, was used. The semen was thawed and washed with PBS to remove cryopreservatives. Spermatozoa were then counted, resuspended in RPMI supplemented with 5% fetal calf serum (FCS) and 0.2% sodium azide to a final dilution of 1 × 106 cells/ml, and analyzed by flow cytometry.
Immunohistochemistry
Immunohistochemistry was performed on cryostat sections using the avidin-biotin complex (ABC) technique using a commercially available kit (Vector Laboratories; Burlingame, CA). Endogenous peroxidases were inhibited with H2O2 (1%) in 0.1% sodium azide. The blocking solution was prepared by mixing 2% milk powder in Tris-buffered saline (TBS), pH 7.6, with normal goat (for the polyclonal antibody) or horse (for monoclonal antibodies) serum. The blocking solution was then applied (30 min at room temperature) to the slides. The primary antibodies listed in Table 1 were diluted in blocking solution and applied to the slides for 1 hr at 37C. The optimal working dilution for each antibody was established by preliminary tests with serial dilution. After three washes in TBS, the biotinylated secondary antibody (30 min at room temperature) and the ABC complex (30 min at room temperature) were added sequentially. Either diaminobenzidine tetrahydrochloride (DAB; Vector Laboratories) or 3-amino-9-ethyl-carbazole (AEC; Vector Laboratories) was used as chromogen for visualization. The reaction was blocked by rinsing in running tap water and the slides were counterstained with Mayer's hematoxylin and coverslipped using non-aqueous mounting medium (Eukitt, Kindler; Freiburg, Germany). Selected histological sections were used as negative controls using normal rabbit serum (DAKO; Glostrup Denmark) as the primary antibody. As positive controls, a section of ovary and a section of testis from adult bovine, for which the distribution of Dpl-positive cells has already been standardized (Rondena et al. 2004), was also included.
Antibodies used in the study
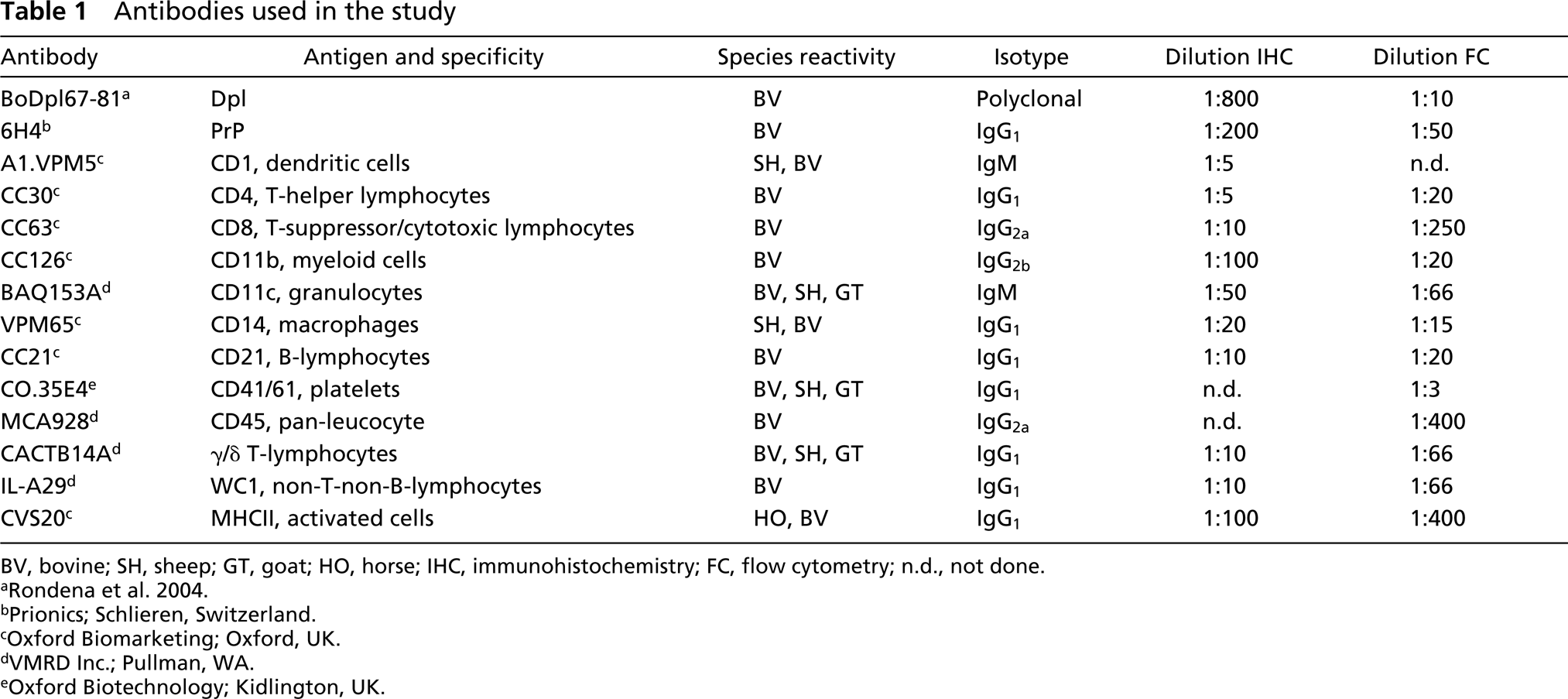
BV, bovine; SH, sheep; GT, goat; HO, horse; IHC, immunohistochemistry; FC, flow cytometry; n.d., not done.
bPrionics; Schlieren, Switzerland.
cOxford Biomarketing; Oxford, UK.
dVMRD Inc.; Pullman, WA.
eOxford Biotechnology; Kidlington, UK.
To examine the coexpression of Dpl, PrP, and leukocyte membrane antigens, double immunohistochemical tests were carried out on serial sections from spleen and lymph nodes. Specifically, the first step of the immunohistochemical staining was done as described above, using anti-CD antibodies or normal mouse serum (DAKO) as a negative control. After development of the immunohistochemical reaction with DAB or AEC, the slides were washed in Tris, blocked with normal goat serum, and the anti-Dpl or the anti-prion antibody was applied overnight at 4C. After washing, the secondary biotinylated antibody and the ABC complex were applied as specified above. This second staining reaction was revealed using AEC or DAB, depending on the chromogen used for the first staining step. The slides were then counterstained with Mayer's hematoxylin and coverslipped using Kaiser's glycerin. The same double immunohistochemical staining protocol was used to study possible coexpression of Dpl and PrP.
Flow Cytometry
Spermatozoa were incubated with the polyclonal anti-Dpl antibody or with a negative rabbit IgG (DAKO) as a control, for 20 min at 4C. They were washed twice, resuspended, and incubated with the secondary antibody (phycoerythrin-conjugated anti-rabbit IgG; Serotec, Oxford, UK) for 20 min at 4C. Blood samples (0.5 ml) were lysed in 7 ml of RBC lysing solution (0.15 M ammonium chloride, 10 mM potassium bicarbonate, and 0.1 mM tetrasodium EDTA in distilled water). After washing, leukocytes were diluted in RPMI containing 5% FCS and 0.2% sodium azide at a final concentration of 1 × 106 cells/ml. Cells were first incubated with the polyclonal anti-Dpl antibody at the dilution established in a previous work (Rondena et al. 2004) or with a negative rabbit IgG (DAKO) as a control, for 20 min at 4C. Cells were then incubated with the secondary antibody (phycoerythrin-conjugated anti-rabbit IgG) for 20 min at 4C. Double staining was performed in a second step, using monoclonal antibodies or isotype-matched murine control sera (Serotec). To establish the best working dilution of each monoclonal antibody, titration of each antibody was performed starting from the dilution suggested by the manufacturers. Negative controls were used at the same IgG concentration. After incubation and washing, the secondary antibody (fluorescein-conjugated rabbit anti-mouse IgG; Serotec) was added.
Both spermatozoa and leukocytes were resuspended in 0.5 ml PBS. Fluorescence was measured on a minimum of 10,000 cells using a FACSort flow cytometer (Becton Dickinson; San Jose, CA). Data were analyzed using CELL Quest software (Becton Dickinson).
Positivity for CD45 was used to distinguish leukocytes from debris and to backgate each population (Figure 1). The percentage of positive events for Dpl in each leukocyte subset or in PrP-expressing cells was then counted and compared with that recorded in control cell population by means of the Kolmogorov Smirnov test. Statistical significance was established at p<0.001.
Results
Immunohistochemistry
Prion protein was expressed in the large majority of cells composing germinal centers of lymphoid follicles, while bovine Dpl was detectable on scattered cells in germinal centers of lymphoid follicles in both spleen and lymph nodes (Table 2). The majority of these cells were morphologically identifiable as lymphocytes (Figure 2A), although some had irregular shapes and/or dendritic processes. Moreover, a diffuse intracytoplasmic positivity for Dpl was detectable on rare myeloid precursors (Figure 2B) and megakaryocytes in bone marrow. Erythroid precursors did not express PrP or Dpl.
Double immunohistochemical staining revealed that both Dpl and PrP were expressed on CD21+ B-cells (Figures 2C and 2D, respectively). Rare CD1+ FDCs showed multifocal cytoplasmic immunoreactivity for Dpl (Figure 2E). Conversely, PrP was strongly expressed on FDCs (Figure 2F). Some cells in the germinal centers of lymphoid follicles co-expressed Dpl and MHCII, whereas coexpression of Dpl and PrP was rare. CD4+ and CD8+ T-cells, γ/δ T-cells, WC1+ lymphocytes, and myeloid cells (CD11b+, CD11c+, or CD14+) were normally distributed in lymphoid organs but never expressed Dpl or PrP.
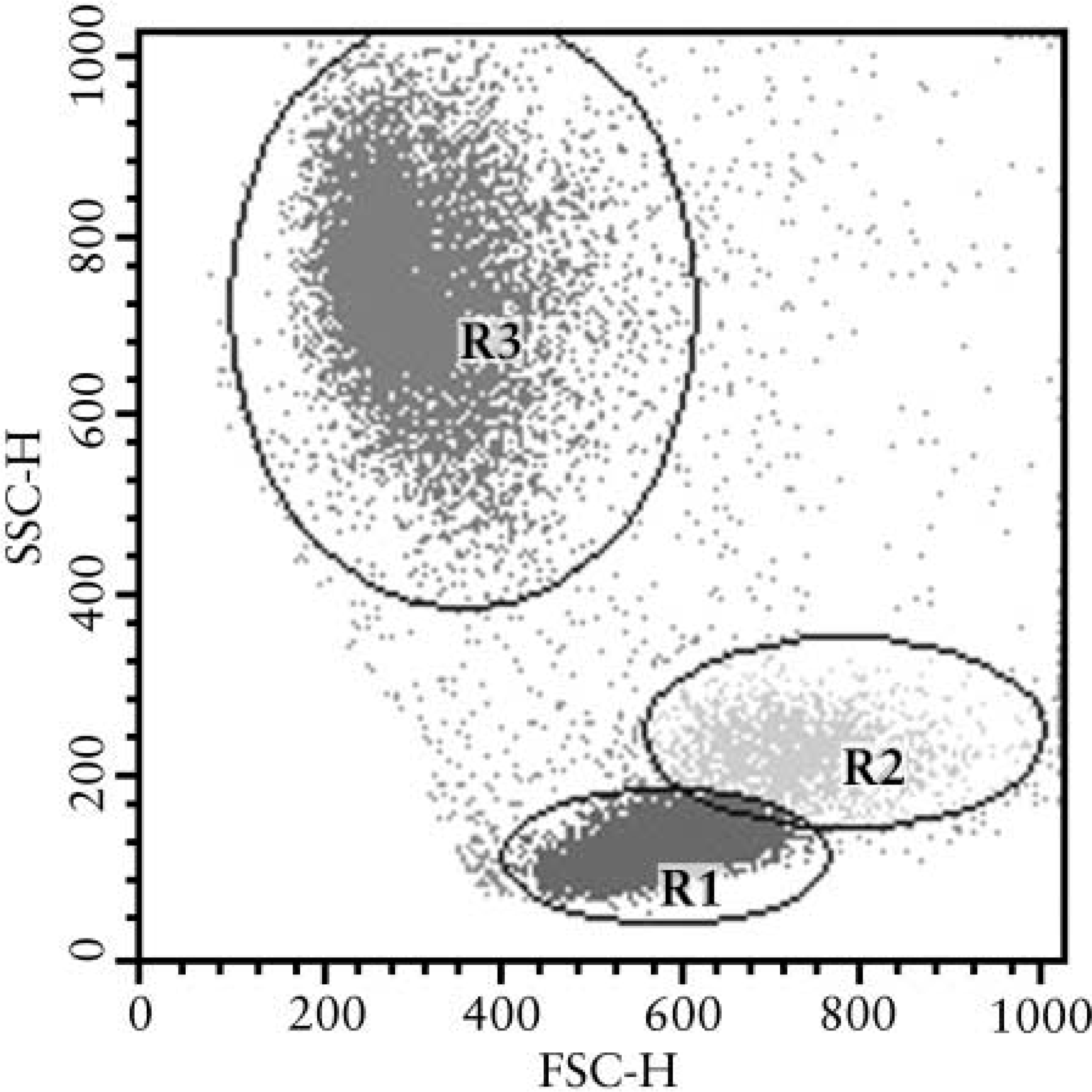
Example of a forward and side scatter plot of bovine blood. CD45+ cells were distributed in three different areas, lymphocytes (R1), monocytes (R2), and neutrophils (R3). Each of these populations has been gated and was analyzed in double staining tests.
Summary of the results of double immunohistochemical (IHC) or flow cytometric (FC) staining for leukocyte subsets membrane antigens and Dpl a
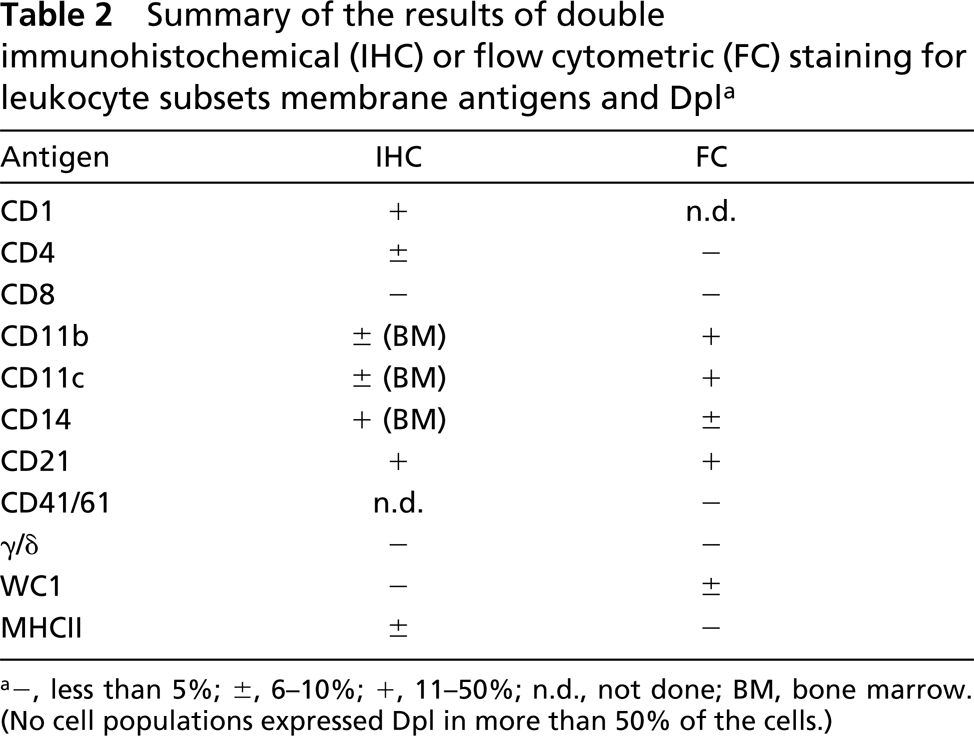
a−, less than 5%; ±, 6-10%; +, 11-50%; n.d., not done; BM, bone marrow (No cell populations expressed Dpl in more than 50% of the cells.)
Flow Cytometry
The flow cytometric protocol to detect Dpl was preliminarily assessed on ejaculated spermatozoa, in which the expression of Dpl was already documented by flow cytometry in humans (Peoc'h et al. 2002) and bovine (Rondena et al. 2004). Using ejaculated spermatozoa as a positive control, the expression of Dpl and its possible relationship with PrP expression was then investigated in blood cells. The results are summarized in Table 2 and shown in Figure 3, which clearly demonstrates the presence of Dpl-expressing cells in different leukocyte subpopulations. Specifically, a positive signal was detected in CD11b+, CD11c+ polymorphonuclear granulocytes (PMNs), and in CD21+ B-lymphocytes, while CD14+ monocytes and other lymphocyte subsets and platelets had weak to absent Dpl positivity (Table 2).
PrP was expressed in lymphocytes and, to a lesser extent, in monocytes and PMNs. Dpl was detected in 18.6% of PrP-expressing cells (Table 2).
Discussion
The detection of Dpl on scattered cells in both spleen and lymph nodes confirmed the results of previous studies in bovine (Rondena et al. 2004). Compared with the PrP, which was widely distributed in lymphoid germinal centers, the distribution of Dpl was limited to scattered cells within germinal centers. Based on their location in lymphoid areas in which a large part of the cells express B-cell antigens and on the results of double immunostaining, Dpl-expressing cells can be identified as B-lymphocytes. PrPc has been reported to be expressed in B-cells and in FDCs within the germinal centers (Klein et al. 1998; Li et al., 2001; Thielen et al. 2001a). PrPsc expression in FDCs is thought to be involved in the pathogenesis of scrapie, the ovine TSE (Mabbot et al. 2000). Our results suggest that Dpl may also be occasionally expressed in FDCs, based on the morphology of some Dpl-expressing cells. Unfortunately, the double immunohistochemical staining using anti-Dpl and anti-CD1 antibodies did not support this hypothesis. Experiments based on FDC isolation, already established for PrP (Thielen et al. 2001b), followed by flow cytometric analysis of Dpl expression will be required to confirm this finding.
Dpl and PrP appear to be expressed in different cell populations in blood. PrP in ruminants has been reported to be expressed in B-cells and, to a lesser extent, in other lymphocyte subsets and monocytes (Herrmann et al. 2001; Barclay et al. 2002). PrP expression in platelets and PMNs has been reported to be low (Herrmann et al. 2001) or absent (Barclay et al. 2002). Our flow cytometric results basically confirm these findings with regard to PrP. Similarly to PrP, Dpl is not expressed on platelets and its expression in B-cells is higher than in other lymphocyte subsets. The strongest Dpl expression, however, was found in circulating PMNs. This finding is in agreement with the immunohistochemical detection of rare positive myeloid cells in bone marrow. This suggests that, contrary to what has been demonstrated for PrP (Dodelet and Cashman 1998), the expression of Dpl increases during PMN maturation and Dpl appears to be downregulated during platelet maturation. Interestingly, previous immunohistochemical investigations did not detect Dpl in resident macrophages or scattered phagocytes that can be occasionally found in bovine tissues (Rondena et al. 2004). Moreover, in the present study myeloid cells identified by antibodies directed against CD11b, CD11c, or CD14 did not stain for Dpl in lymphoid tissues. Resting and/or non-circulating phagocytes therefore appear to downregulate Dpl, suggesting that the activation state might influence Dpl expression, as already demonstrated for PrP, which is differentially expressed during maturation of leukocytes or platelets (Holada et al. 1998; Durig et al. 2000). This possibility is supported by our observation that many Dpl-expressing lymphocytes co-express MHCII. The possible expression of Dpl in activated cells merits further investigation, because it cannot be excluded that phagocytes recruited to inflamed sites might express Dpl.
It has been demonstrated that PrP and Dpl are expressed on different membrane rafts (Shaked et al. 2002), and the observed coexpression of Dpl and PrP in blood and tissues confirms this evidence. However, the number of cells expressing both proteins was very low, suggesting that the mechanisms involved in Dpl expression in resting blood cells are probably different from and independent of those responsible for PrP expression. Likewise, the possible coexpression of PrP and Dpl and the influence of PrP on Dpl expression (and vice versa) would be of interest to investigate in blood and tissues from animals affected by TSE and/or in an experimental model based on PrP/Dpl blocking or deletion.
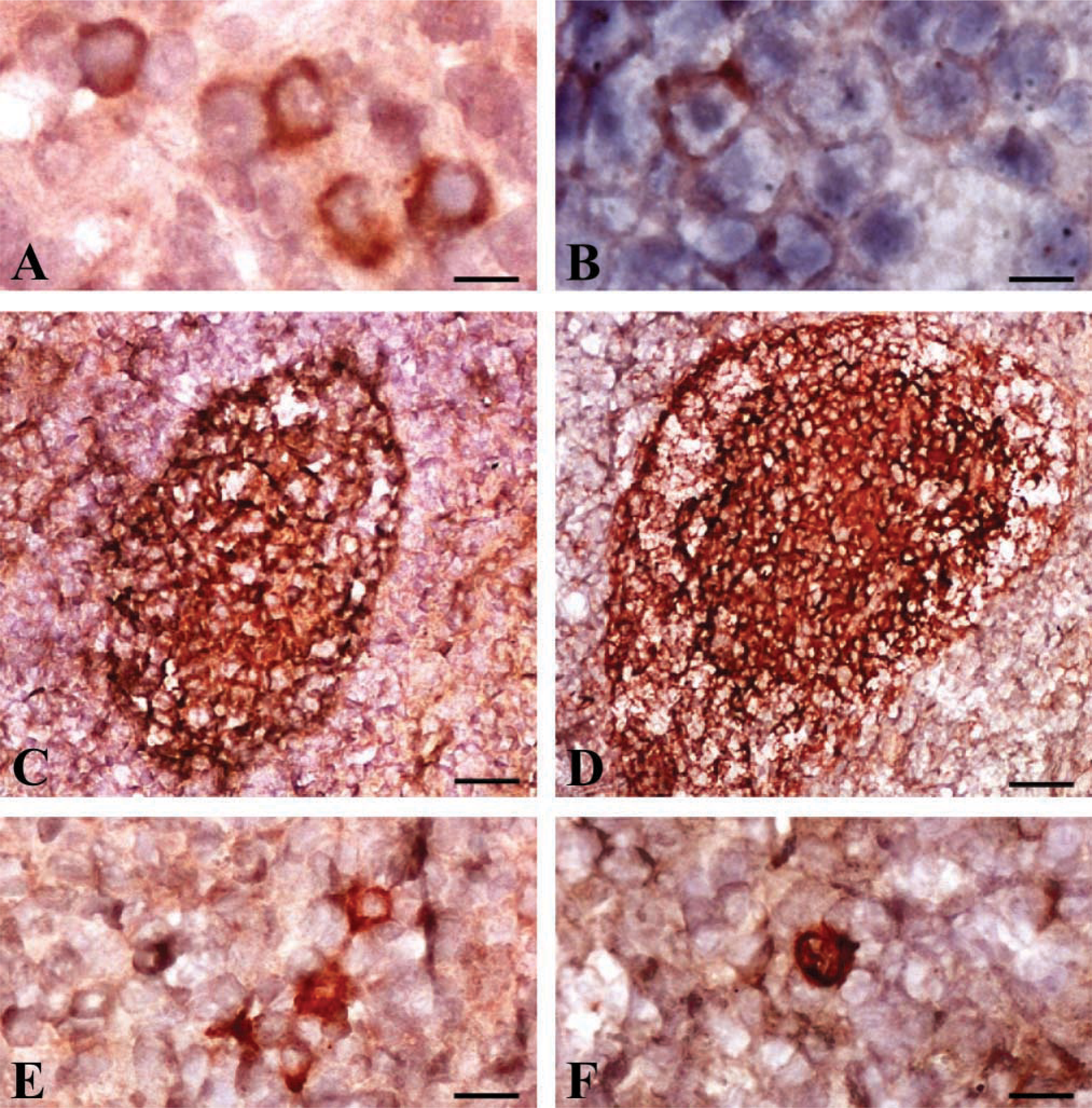
Immunohistochemical distribution of bovine Dpl (
Other factors that might influence the expression of Dpl in blood and lymphoid cells remain to be elucidated. On the basis of the results of previous reports (Rondena et al. 2004) that did not show any agedependent differential expression of bovine Dpl, it might reasonably be excluded that age might influence Dpl expression in blood. However, the possible influence exerted by host genetic factors in conditioning different expression levels of both Dpl and PrP in bovine leukocytes must also be examined because prnd, the Dpl-encoding gene in bovine, has been shown to be polymorphic (Comincini et al. 2001).
The information obtained from this study might therefore be translated to murine models of PrP-induced diseases or to the analysis of human PrP and Dpl biology because additional homologies between the two proteins have been identified. However, in spite of a similar distribution in both lymphoid cells and tissues, PrP and Dpl are rarely co-expressed, and the following differences in the expression pattern of the two proteins were detected: (a) the number of cells expressing Dpl is lower than that expressing PrP; (b) PMNs express Dpl but not PrP; and (c) Dpl expression appears to vary with the activation state of the cells. Our results further support the hypothesis that Dpl is not involved in spontaneous TSE. Nevertheless, blood might be easily used to design in vitro tests to differentially enhance or suppress the expression of the two proteins to gain useful information about the pathobiology of Dpl and PrP.
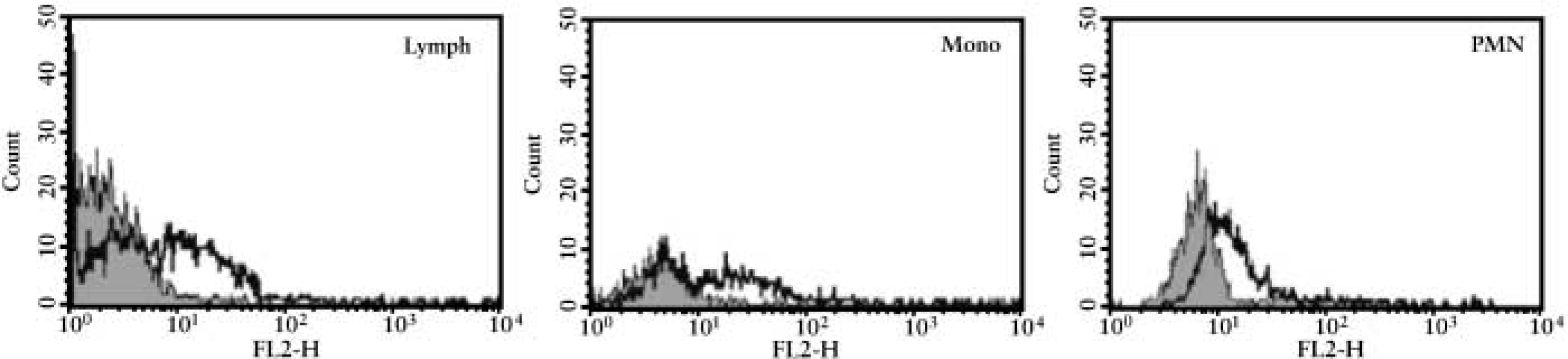
Flow cytometric evaluation of Dpl expression (black line) on gated circulating lymphocytes (Lymph), mono-cytes (Mono), and neutrophils (PMN) compared with cells stained with an irrelevant isotype-matched antibody (gray area). Dpl expression is detectable on gated PMNs and on some lymphocyte and monocyte subpopulations.
Footnotes
Acknowledgements
Supported by grant COFIN 2001 from the Italian Government.
We are very grateful to Prof Giorgio Cammarata, Prof Cecilia Luvoni, and Dr Sara Chigioni (University of Milan) and to Prof Luca Ferretti (University of Pavia).