
Abstract
We compared two methods to stain apoptotic cells, one using terminal deoxynucleotidyl transferase (TDT), the other DNA polymerase I, using leukemia cell lines treated with anti-Fas monoclonal antibody (MAb). Both TDT and polymerase I strongly reacted with fragmented nuclei of apoptotic MOLT-16 and Jurkat cells, but only polymerase I strongly reacted with nonfragmented nuclei of early apoptotic cells. Anti-Fas MAb-treated MOLT-4 cells showed morphological changes corresponding to early apoptosis and were strongly positive for polymerase I only. MOLT-16 and Jurkat cells treated with anti-Fas MAb and inhibitors of endonuclease and poly(ADP-ribose) polymerase showed the morphology of early apoptosis but were not strongly stained by TDT. Because DNA polymerase I has nick-translation activity, it is possible that DNA polymerase I reaction is positive in early apoptotic cells by detecting single-strand DNA cleavage, which occurs before extensive oligonucleosomal DNA cleavage and late morphological changes of apoptosis in leukemia cell lines. Although TDT is widely used to stain apoptotic cells, DNA polymerase I may be more applicable in special cases of apoptosis, in which cells undergo single-strand rather than double-strand DNA breaks. However, the procedure has limitations, such as the necessity to use cell smears for comparison with the TDT reaction.
Keywords
A
Many lymphoma/leukemia cell lines undergo apoptosis in the presence of anti-Fas monoclonal antibody (MAb) (Kondo et al. 1994), and inhibitors of endonuclease and poly(ADP-ribose) polymerase (PARP) have been known to modify the process of apoptosis (Cohen et al. 1992; Sumimoto et al. 1994; Yoon et al. 1996). Using these cell lines, anti-Fas MAb, and inhibitors, the apoptotic process was examined with TDT and DNA polymerase I staining and the results were compared.
Materials and Methods
Cells
The human leukemia cell lines, Jurkat, MOLT-16, and MOLT-4 have been maintained at Fujisaki Cell Center, Hayashibara Biochemical Laboratories (Okayama, Japan) and well characterized (Minowada 1988). For induction of apoptosis, anti-Fas mouse IgM MAb (CH-11; MBL, Nagoya, Japan) was added to cell suspensions of 5.0 × 105 viable cells/ml in culture flasks at concentrations of 25, 50, and 100 ng/ml. The concentrations of inhibitors of Ca2+/Mg2+-dependent endonuclease and PARP were chosen on the basis of earlier reports (Durkacz et al. 1980; Creissen and Shall 1982; Seto et al. 1985; Cohen et al. 1992; Sumimoto et al. 1994): 0.1, 0.25, 0.5, and 1.0 mM for Zn2+ (ZnSO4); 3 and 5 mM for 3-aminobenzamide (Sigma Chemical; St Louis, MO); 1, 2.5, 5, and 10 mM for benzamide (Aldrich Chemical; Milwaukee, WI); and 5 and 10 mM for nicotinamide (Sigma Chemical). For experiments with inhibitors, anti-Fas MAb at the concentration of 100 ng/ml and inhibitors were added simultaneously to the cell suspension.
Morphological Studies
For light microscopic observation, cells were collected at 1, 3, 6, 12, and 24 hr after the start of the treatment, smeared on glass slides, air-dried, and stained with May–Grünwald–Giemsa staining. Several hundred cells on each smear were counted to evaluate apoptotic reactions. For electron microscopy, cell pellets were fixed in 3% glutaraldehyde and 1% osmium tetroxide and embedded in Epon resin. Ultrathin sections were stained with uranyl acetate and lead citrate and observed with a transmission electron microscope (JEM-1200 EX II; Japan Electron Optics Laboratory, Tokyo, Japan).
Staining of Apoptotic Cells
The TDT and DNA polymerase I reactions were performed as previously reported (Gavrieli et al. 1992; Gold et al. 1993; Wijsman et al. 1993) with some modifications (Aftabuddin et al. 1995). In brief, air-dried cell smears were pretreated with 1 M NaSCN at 70C for 20 min and with 20 μg/ml proteinase K at room temperature for 20 min. For the TDT reaction, 140 U/ml TDT (TOYOBO; Osaka, Japan) was incubated with 0.01 mM biotin-16-labeled dUTP (Boehringer Mannheim; Mannheim, Germany) at 37C for 30 min. For the DNA polymerase I reaction, 20 U/ml DNA polymerase I (TOYOBO) was incubated with 0.01 mM dATP, dCTP, dGTP, and biotin-16-labeled dUTP at 37C for 90 min. After incubation with peroxidase-conjugated streptavidin (Scytek; Logan, UT), the reactions were developed with diaminobenzidine and H2O2. Smears were counterstained with methyl green.
DNA Fragmentation Assay
DNA was extracted from cells by the phenol/chloroform method after digestion with proteinase K and was then electrophoresed on a 1.2% SeaKem ME agarose gel (FMC Bio-Product; Rockland, ME).
Statistical Analysis
The mode of apoptosis was compared based on the ratio of apoptotic cells with fragmented nuclei to apoptotic cells with nonfragmented nuclei. The difference was tested by the χ2 test.
Results
Light Microscopy
Anti-Fas MAb induced apoptosis of typical morphology (Kerr et al. 1972; Arends et al. 1990) in Jurkat and MOLT-16 cell lines (Figure 1A). At 12–24 hr of MAb treatment, most cells were either apoptotic or dead. Most apoptotic cells during this period had the morphology of late apoptosis, with condensed cytoplasm and a fragmented nucleus completely composed of condensed chromatin. Apoptotic bodies, i.e., small cell fragments containing fragmented nuclei, were also observed. At 3–12 hr of treatment, cells at various stages of apoptosis were detected, with cells in the early stage of apoptosis exhibiting condensed chromatin in the periphery of nonfragmented nuclei and lucent areas around condensed chromatin. At 1–3 hr, a few cells in the early stage of apoptosis were scattered among cells of normal morphology (Figure 1B). The time course of apoptosis is shown in Figure 2. Apoptosis progressed faster in MOLT-16 cells than in Jurkat cells.
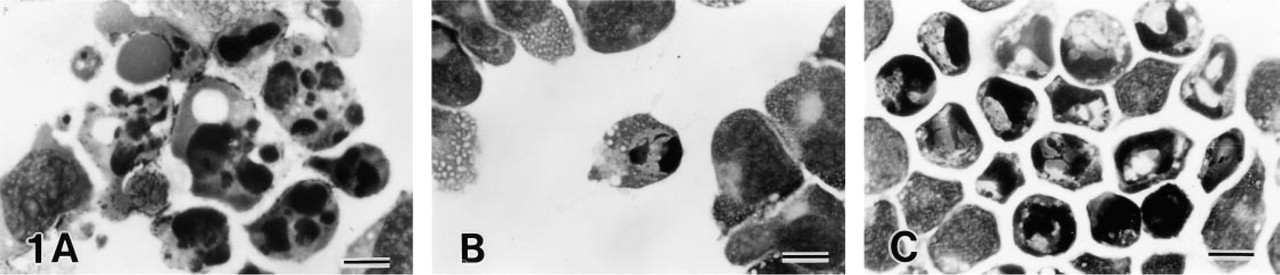
Light microscopy of leukemia cell lines treated with 100 ng/ml of anti-Fas MAb. (
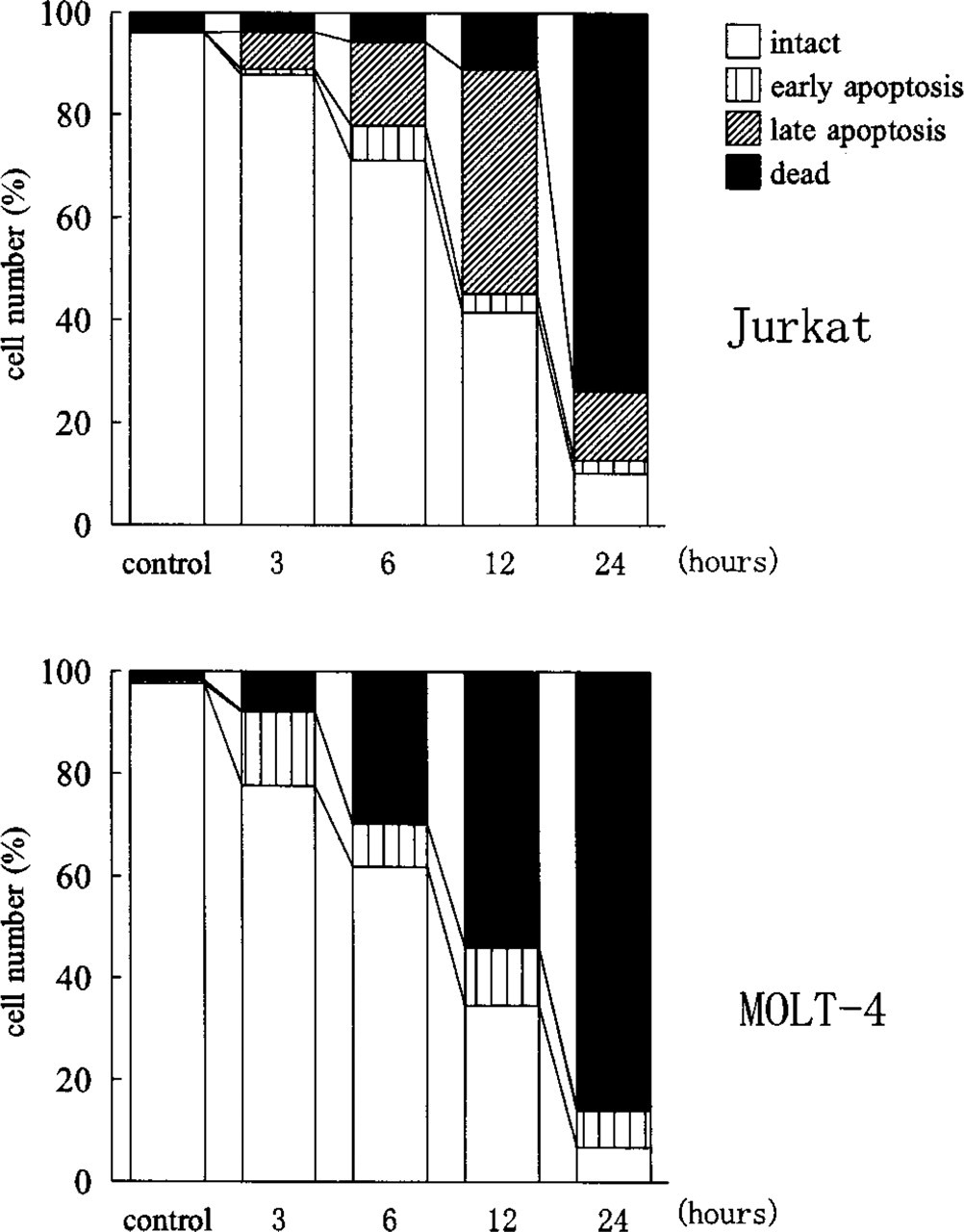
Representative time courses of the apoptosis induced by 100 ng/ml anti-Fas MAb in Jurkat and MOLT-4 cells in stained smears. The numbers of intact cells, early apoptotic cells with non-fragmented nuclei, late apoptotic cells with fragmented nuclei, and dead cells are presented as percentages.
MOLT-4 cells reacted differently to anti-Fas MAb. Only a few typical apoptotic cells with fragmented nuclei appeared. Most reacting cells had condensed chromatin, but nuclei had lucent areas and did not become small (Figure 1C). The morphology of these cells was similar to that of Jurkat or MOLT-16 cells in the early stage of apoptosis. At the same time, many dead cells appeared early (Figure 2). The percentage of early apoptotic cells was much higher than in MOLT-16 or Jurkat cells (p<0.01). The concentration of anti-Fas MAb affected the number of apoptotic cells, but the mode of apoptosis was the same in all three cell lines.
Electron Microscopy
Anti-Fas MAb-treated Jurkat and MOLT-16 cells showed cytoplasmic and nuclear condensation; nuclei were fragmented and filled with compacted chromatin (Figure 3A). Apoptotic bodies were also observed, discrete cellular fragments containing one or more round small nuclear fragments filled with condensed chromatin. MOLT-4 cells treated with anti-Fas MAb also showed cytoplasmic and nuclear condensation, but nuclear fragmentation was minimal, and dense masses of condensed chromatin were often observed around the circumference of the nuclear envelope (Figure 3B).
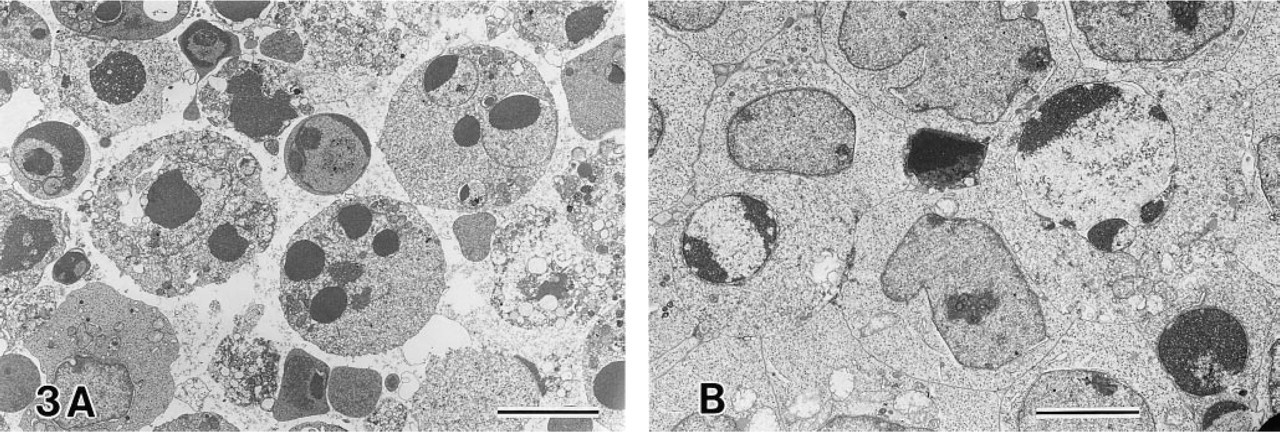
Electron microscopy of anti-Fas MAb-treated MOLT-16 (
DNA Fragmentation Assay
Anti-Fas MAb-treated MOLT-16 and Jurkat cells showed a clear ladder pattern of DNA fragmentation, but anti-Fas MAb-treated MOLT-4 cells showed minimal DNA fragmentation and no ladder pattern (Figure 4).
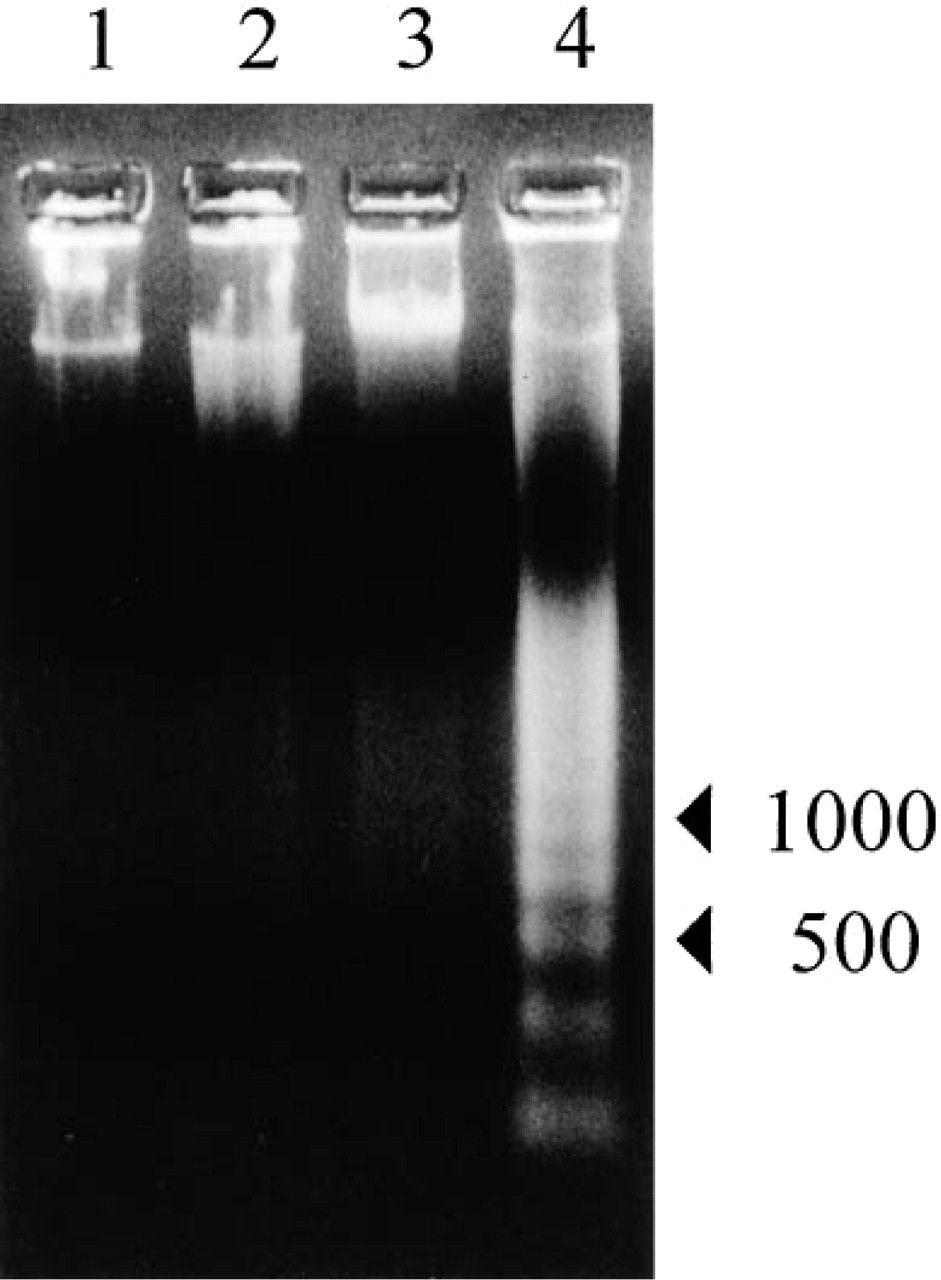
DNA fragmentation assay. 1, Control MOLT-4 cells; 2, anti-Fas MAb-treated MOLT-4 cells; 3, control MOLT-16 cells; 4, anti-Fas MAb-treated MOLT-16 cells. Anti-Fas MAb was added at a concentration of 100 ng/ml for 12 hr. DNA fragmentation was of a ladder pattern in anti-Fas MAb-treated MOLT-16 cells, but was minimal in anti-Fas MAb-treated MOLT-4 cells.
TDT and DNA Polymerase I Reactions
Jurkat and MOLT-16 cells showing typical apoptotic morphology with fragmented nuclei were strongly positive for both TDT and DNA polymerase I (Figures 5A and 6A). Cells at the early stage of apoptosis without nuclear fragmentation were weakly positive for TDT but clearly positive for DNA polymerase I (Figures 5A and 6A). Apoptotic MOLT-4 cells were strongly positive for DNA polymerase I but only weakly positive for TDT (Figures 5B and 6B).
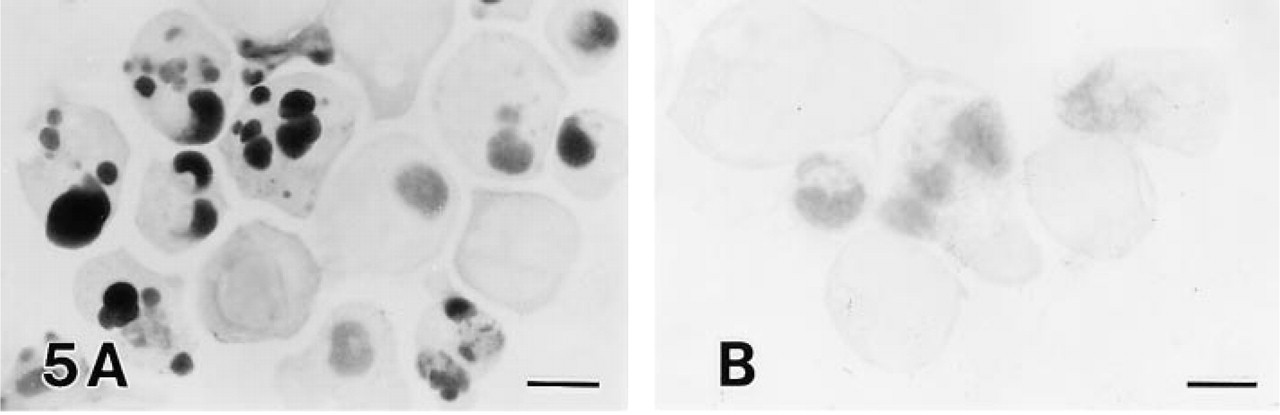
TDT reaction in anti-Fas MAb-treated Jurkat (
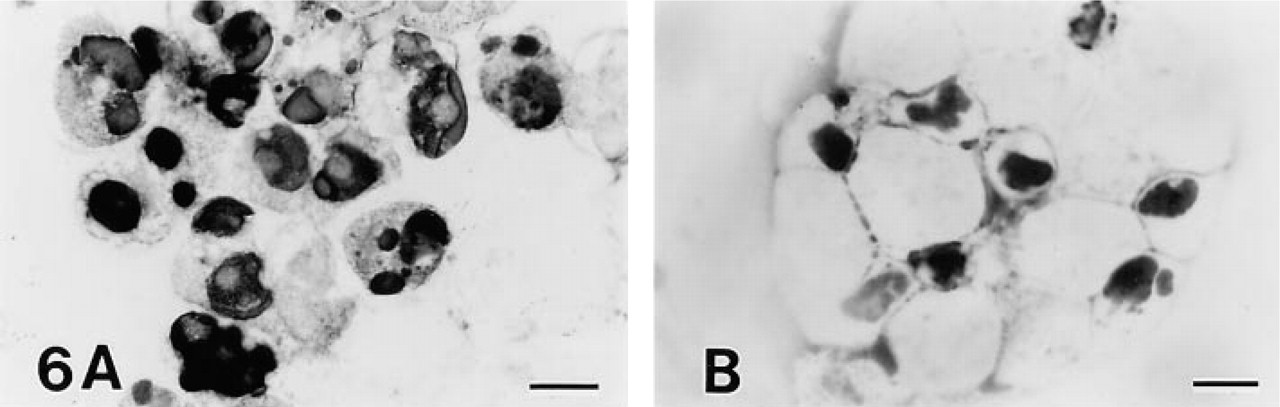
DNA polymerase I reaction in anti-Fas MAb-treated Jurkat (
Effect of Inhibitors of Endonuclease and PARP
The addition of Zn2+, an inhibitor of Ca2+/Mg2+-de-pendent endonuclease, to the cell suspension caused some degree of cytoplasmic condensation and chromatin aggregation even without treatment with anti-Fas MAb. None of the usual apoptotic changes induced by anti-Fas MAb were observed in Jurkat and MOLT-16 cells on addition of 1 mM Zn2+. Very few of the apoptotic cells appeared typical, with most showing nuclear morphology consistent with early apoptosis (lacking nuclear fragmentation) (Figures 7A and 8). The percentage of early apoptotic cells was much higher than in cells treated only with anti-Fas MAb (p<0.01). The addition of Zn2+ did not prevent cell death. In all three cell lines treated with anti-Fas MAb and Zn2+, very few apoptotic cells were positive for TDT. Zn2+ was not effective in anti-Fas MAb-treated MOLT-4 cells.
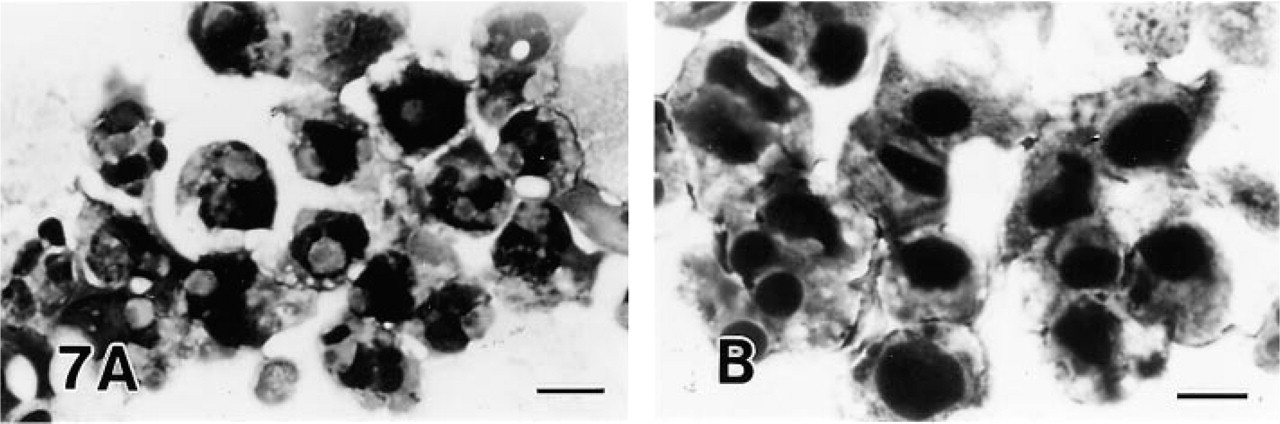
The effect of inhibitors of endonuclease and poly(ADP-ribose) polymerase. (
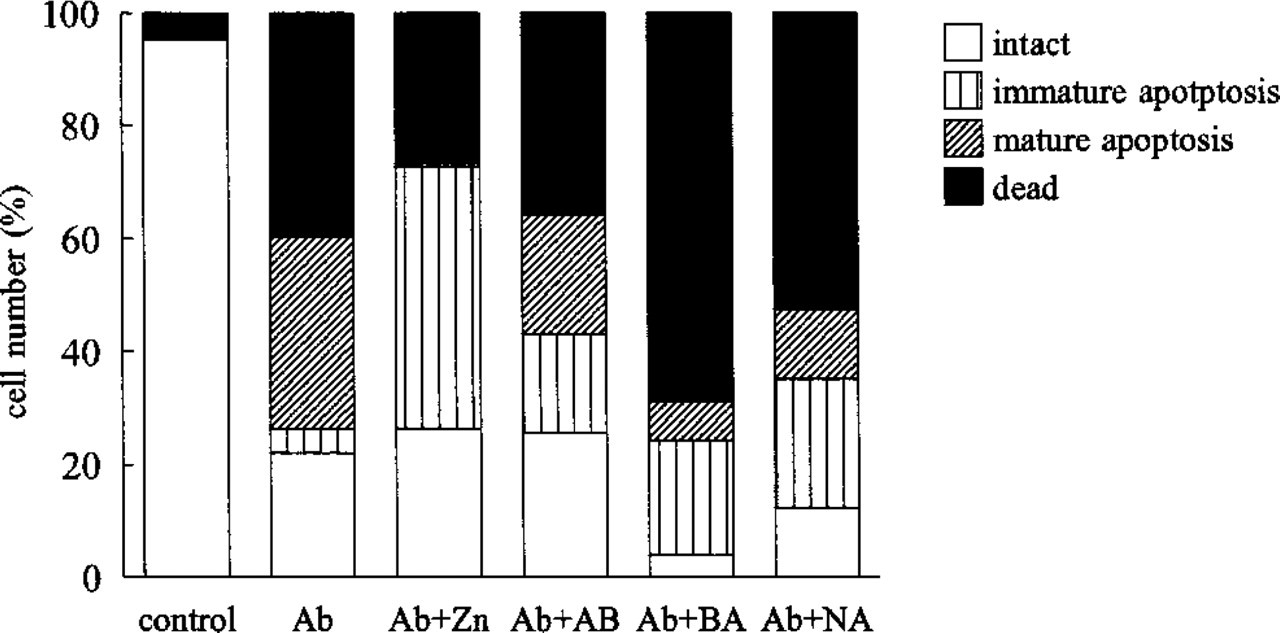
The effect of inhibitors of endonuclease and poly(ADP-ribose) polymerase on anti-Fas MAb treatment of Jurkat cells. Shown are cell counts in representative experiments with various treatments done in the same cell suspension on the same day. Ab, 100 ng/ml anti-Fas MAb; Zn, 1 mM Zn2+; AB, 3 mM aminobenzamide; BA, 10 mM benzamide; NA, 10 mM nicotinamide.
Treatment with inhibitors of PARP, aminobenzamide, benzamide, or nicotinamide, in addition to anti-Fas MAb, in MOLT-16 and Jurkat cells remarkably reduced the percentage of typical apoptotic cells (p< 0.01), and produced many early apoptotic cells without nuclear fragmentation (Figures 7B and 8). Many dead cells also appeared. No such effect occurred in MOLT-4 cells treated with anti-Fas MAb. Cells treated only with PARP inhibitors were morphologically unchanged. No cells positive for TDT were detected on combined treatment with anti-Fas MAb and PARP inhibitors.
The effect of inhibitors of endonuclease and PARP on anti-Fas MAb-treated cells was basically the same as the effect of anti-Fas MAb and Zn2+ treatment. The cells were shrunken as with Zn2+ treatment, the nuclear morphology was basically that of early apoptosis, and the appearance of dead cells was not prevented.
DNA polymerase I did not react well in experiments with inhibitors. The reaction was weak overall and could not be judged as positive or negative.
Discussion
The TUNEL method is widely used to demonstrate apoptosis in histology sections, but we showed in this experiment that TDT and DNA polymerase I reactions give different staining results in leukemia cell lines. These two procedures utilize different enzymatic reactions detecting different kinds of DNA breaks.
In addition to oligonucleosomal cleavage by endonuclease, other forms of DNA cleavage have been observed during apoptosis (Bortner et al. 1995). One is the generation of 300- or 50-KBP DNA fragments detected by field gel electrophoresis (Oberhammer et al. 1993). Another is single-strand breaks (Sumimoto et al. 1994). In general, single-strand breaks in dsDNA (nicks) are quickly repaired in relation to the activation of PARP (Durkacz et al. 1980; Creissen and Shall 1982), resulting in the synthesis of poly(ADP-ribose), a polymer produced from NAD. Consumption and severe depletion of NAD is followed by decreased ATP generation and is incompatible with cell survival (Seto et al. 1985). Generation of single-strand DNA breaks is presumed to be one of the major causes of cell death by apoptosis in leukemia cell lines.
It has been reported that thymocytes undergo apoptosis when incubated with dexamethasone, but in the presence of Zn2+ cell death morphologically resembles early apoptosis but without the DNA laddering (Cohen et al. 1992). In apoptosis of a human leukemia cell line caused by anti-Fas MAb, Zn2+ did not prevent cell death and single-strand breaks were increased, as determined by the alkaline DNA unwinding assay (Sumimoto et al. 1994). It has been proposed that poly(ADP-ribosyl)ation of nuclear histone protein in the early stage of apoptosis increases the accessibility to chromatin of cellular endonuclease and facilitates internucleosomal DNA fragmentation (Yoon et al. 1996).
Because DNA polymerase I has nick-translation activity, it is possible that DNA polymerase I reaction is positive in early apoptotic cells by detecting single-strand DNA cleavage, which occurs before extensive oligonucleosomal DNA cleavage and late morphological changes of apoptosis in leukemia cell lines. Our staining results were consistent with the idea that activation of endonuclease is a later event in the apoptotic process (Cohen et al. 1992; Bortner et al. 1995). The results with inhibitors of endonuclease and PARP were also consistent with this presumption.
The MOLT-4 cell line responded to anti-Fas MAb in a manner different from the other leukemia cell lines examined. Cells died with the morphology of early stage apoptosis. Apoptotic cells were weakly positive for TDT but strongly positive for DNA polymerase I, and did not show a ladder pattern of DNA fragmentation on gel electrophoresis. Similar examples of cell death have been documented in the literature. Although surface IgM crosslinking or treatment with anti-Fas MAb induced apoptosis in several IgM+ B-leukemia and Fas antigen-positive leukemia cell lines, the cell death in B104 cells treated with anti-IgM or anti-Fas MAb more closely resembles necrosis than apoptosis (Ishigami et al. 1992; Sumimoto et al. 1994). Cell death in the regression stage of human lactating breast, in which single-strand DNA breaks accumulate, has recently been reported to be neither necrotic nor apoptotic (Uemura et al. 1996). This type of programmed cell death probably involves a deficiency in endonuclease activation, as in the MOLT-4 cells in the present study. It is interesting that irradiation caused programmed cell death with features of both apoptosis and necrosis in MOLT-4 cells, but with features of apoptosis only in other leukemia cell lines (Akagi et al. 1993). There might be certain inherent characteristics in MOLT-4 cells whereby stimuli that induce typical apoptosis in other cell lines produce cell death deficient in endonuclease activation. The extent to which anti-Fas MAb treatment activates endonuclease may cause the different morphology of apoptosis between MOLT-4 and other cell lines, because differences among leukemia cell lines have been reported in the participation of intracellular NAD, activation of PARP, and subsequent NAD depletion in apoptosis (Wright et al. 1996).
As pointed out by Duke et al. (1995), TDT may fail to stain apoptotic cells that do not undergo typical double-strand DNA breaks. In special cases of apoptosis, such as in MOLT-4 cells treated with anti-Fas MAb, DNA polymerase I is better suited to detecting DNA cleavage. Moreover, a discrepancy between TDT and DNA polymerase I staining can suggest single-strand DNA cleavage. It should be noted, however, that different results in TDT and DNA polymerase I staining can be obtained only in cell smears. In our preliminary experiments using paraffin sections of cell pellets, both reactions yielded the same results, presumably because DNA degradation during processing involved single- to double-strand breaks. Furthermore, staining of apoptotic cells is more difficult in smears than in paraffin-embedded tissue sections, probably because of the poor accessibility of the enzymes to nuclear DNA. Pretreatment of smears with NaSCN, proteinase K, or other chemicals is crucial to ensure good results. In addition, DNA polymerase I staining appeared to be more readily influenced by residual chemicals on cell smears, because the reaction did not proceed in experiments with inhibitors. Although the use of DNA polymerase I has such limitations, its applicability to different mechanisms of apoptosis should not be ignored.
Footnotes
Acknowledgements
We thank Tadao Zouda for technical assistance with electron microscopy and Yoshiko Senda for secretarial work.