
Abstract
Prostaglandin endoperoxide synthase (PGHS) is a key rate-limiting enzyme in prostaglandin biosynthesis. PGHS has recently been shown to be expressed in human colorectal cancers and in experimental cutaneous papillomas and carcinomas. However, PGHS expression has not been investigated in ovarian cancers. The objectives of this study were to determine whether PGHS isoenzymes are expressed in human ovarian cancer and, if so, to identify which isoform is involved (PGHS-1 and/or PGHS-2) and to characterize its cellular localization. Sixteen human ovarian adenocarcinomas were studied by immunohistochemistry using specific antibodies recognizing PGHS-1 or PGHS-2. Results showed that all adenocarcinomas demonstrated the presence of tumor cells expressing PGHS-1 but not PGHS-2. Patterns of staining of tumor cells varied among different types of adenocarcinomas, with cells presenting either a mostly diffuse cytoplasmic immunoreactivity or, alternatively, a staining mainly concentrated around the nucleus. No correlation between the intensity of the immunostaining and the degree of malignancy of tumors could be established (r −0.03: p>0.05). Immunoblot analysis with PGHS-1-selective antibodies of cell extracts from adenocarcinomas revealed the presence of a characteristic 72,000 Mr immunoreactive band. Therefore, these results show for the first time that PGHS-1 is expressed in human ovarian adenocarcinomas.
P
Prostaglandin endoperoxide synthase (PGHS) is the first rate-limiting enzyme in the biosynthetic pathway of prostaglandins from arachidonic acid (DeWitt et al. 1993; Funk 1993). It is now well established that two different isoforms of PGHS exist in tissues (Miyamoto et al. 1976; Simmons et al. 1989; Sirois and Richards 1992), and that these isoforms are differently regulated. PGHS-1 (also known as COX-1) is constitutively expressed in various tissues such as brain, kidney, uterus, and fibroblasts, and appears to be involved in the synthesis of prostaglandins necessary for normal cellular processes (Kujubu et al. 1991; Wong and Richards 1991; Kawasaki et al. 1993). In contrast, PGHS-2 (COX-2) is transiently induced by different agonists, including growth factors, phorbol esters, interleukin-1, endotoxins, and hormones (Kujubu et al. 1993; Lee et al. 1993; Lyons-Giordano et al. 1993), and is associated with prostaglandin production during inflammation (Herschman et al. 1995) and ovulation (Sirois et al. 1992).
Recent studies have shown that PGHS-2 is elevated in colon cancers and might be responsible for the elevated levels of prostaglandins in these tumors (Eber-hart et al. 1994; Kargman et al. 1995; Sano et al. 1995; Kutchera et al. 1996). Overexpression of PGHS-2 has also been demonstrated in cutaneous papillomas and carcinomas induced using a mouse skin model of tumor development (Müller-Decker et al. 1995). Tumors of the ovary are common neoplasms, being the second most common gynecological malignancy in the United States (Wingo et al. 1995). The majority of primary ovarian tumors are epithelial tumors derived from the surface epithelium covering the ovary. The expression of PGHS in ovarian tumors has not yet been investigated. Therefore, the objectives of the present study were to determine whether PGHS isoenzymes are expressed in human ovarian cancer and, if so, to identify which isoform is involved (PGHS-1 and/or PGHS-2) and to determine its cellular localization.
Materials and Methods
Tissue Samples
Sixteen cases of ovarian tumors surgically resected from women were studied. The patients' ages ranged from 22 to 82 years, and all tumors included in the study were diagnosed as either malignant tumors or as tumors of borderline malignancy (carcinomas of low malignant potential) (Table 1) according to the “Histologic Classification of Ovarian Tumors by the World Health Organization” (Serov et al. 1973). One benign tumor (a serous cystadenoma) was also included in the study. Control normal ovaries were obtained from three women who had undergone oophorectomy for reasons unrelated to ovarian neoplasia. All tissues were fixed in 10% formalin. Fresh tissues from two tumors were frozen at −70C for immunoblot analysis.
Patient data and PGHS-1 expression in ovarian adenocarcinomas
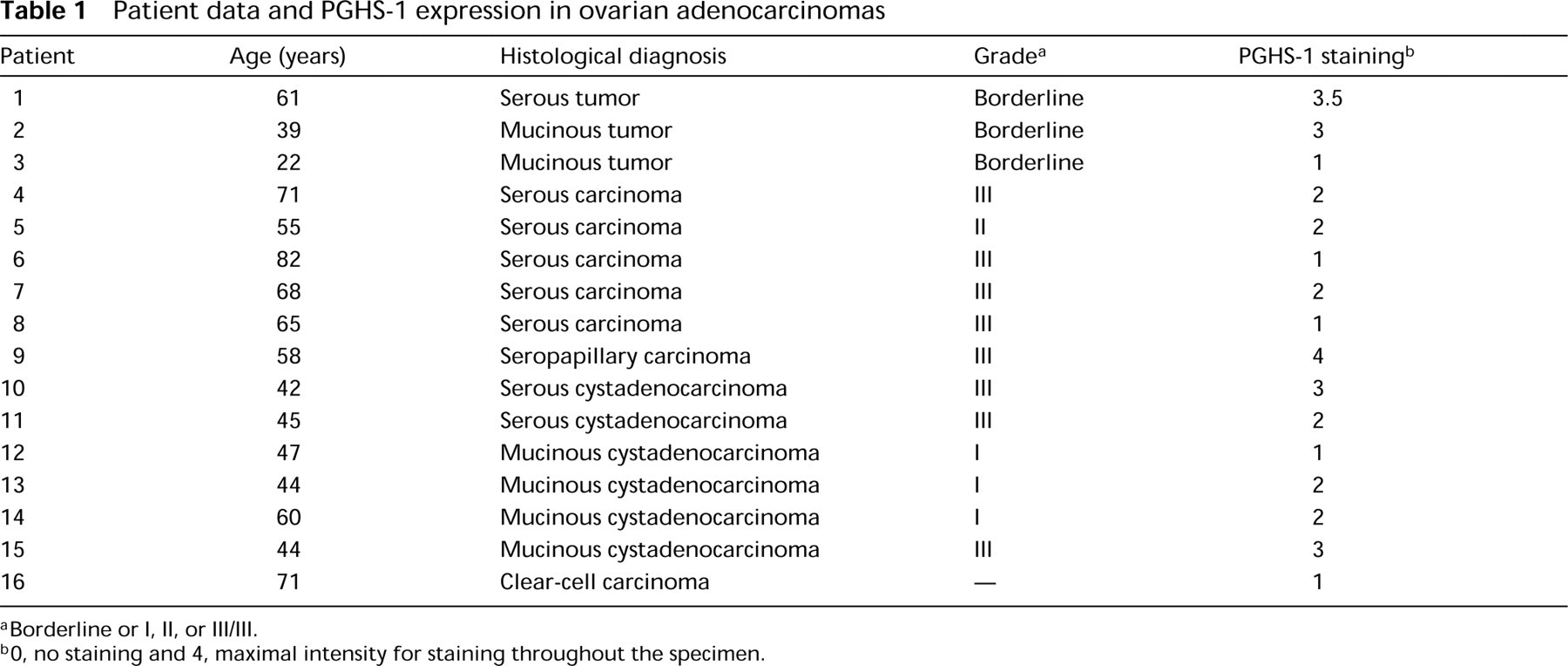
Borderline or I, II, or III/III.
O, no staining and 4, maximal intensity for staining throughout the specimen.
Antibodies
Affinity-purified polyclonal antibodies 9181 and 8223 were raised in rabbits against ovine PGHS-1 (Wong and Richards 1991; Sirois et al. 1992). Their specificity has previously been characterized. Antibody 9181 has been shown to recognize both PGHS isoforms (PGHS-1 and PGHS-2), whereas antibody 8223 has been shown to be selective for PGHS-1. Two additional anti-PGHS antibodies were used to determine which isoform was present. These antibodies (MF241 and MF243) were generously provided by Drs. Jilly F. Evans and Stacia Kargman (Merck Frosst Centre for Therapeutic Research; Pointe-Claire-Dorval, Québec, Canada). MF241 and MF243 were raised in rabbits against full-length sheep seminal vesicle PGHS-1 (for MF241) and placental PGHS-2 (for MF243) purified proteins. The specificity of these anti-PGHS antibodies has previously been characterized (Kargman et al. 1995). They recognize the homologous human PGHS isoforms with approximately 1000-fold selectivity for the appropriate isoform. The specificity of antibody MF243 was tested and confirmed in our hands by immunohistochemistry on samples known to selectively express PGHS-2 (bovine and equine preovulatory follicles; data not shown).
Immunohistochemistry
Immunohistochemical staining was performed using the Vectastain ABC kit (Vector Laboratories; Burlingame, CA), as previously described (Doré et al. 1993). Briefly, formalin-fixed tissues were paraffin-embedded and 3-μm-thick sections were prepared, and deparaffinized through graded alcohol series. Endogenous peroxidase was quenched by incubating the slides in 0.3% hydrogen peroxide in methanol for 30 min. After rinsing in PBS for 15 min, sections were incubated with diluted normal goat serum for 20 min at room temperature (RT). Primary antibodies diluted in PBS were applied (9181 and 8223 at 1:100 dilution; MF241 and MF243 at 1:5000 and 1:7500 dilution) and sections were incubated overnight at 4C. Control sections were incubated with PBS only or with nonimmune rabbit serum. After rinsing in PBS for 10 min, a biotinylated goat anti-rabbit antibody (1:222 dilution) was applied and sections were incubated for 45 min at RT. Sections were washed in PBS for 10 min and incubated with the avidin DH-biotinylated horseradish peroxidase H reagents for 45 min at RT. After a PBS wash for 10 min, the reaction was revealed using diami-nobenzidine tetrahydrochloride (DAB) as the peroxidase substrate. Sections were counterstained with Gill's hematoxylin stain and mounted. Immunoreactivity was evaluated in a blinded fashion using a grading system ranging from 0 to 4, in which 0 = no staining and 4 = maximal intensity of staining throughout the specimen.
Solubilized Cell Extracts and Immunoblot Analysis
Tissues from ovarian carcinomas were homogenized on ice in TED buffer [50 mM Tris (pH 8.0), 10 mM EDTA, and 1 mM DEDTC] containing 0.1% Tween-20 and centrifuged at 30,000 × g for 1 hr at 4C. The crude pellets (membranes, nuclei, and mitochondria) were sonicated (8 sec/cycle; three cycles) in TED sonication buffer [20 mM Tris (pH 8.0), 50 mM EDTA, and 0.1 mM DEDTC] containing 1% Tween-20. The sonicates were centrifuged at 16,000 × g for 15 min at 4C. The recovered supernatant (solubilized cell extract) was stored at −70C until electrophoretic analyses were performed. The protein concentration was determined by the method of Bradford (Bio-Rad Protein Assay; Richmond, CA). Proteins (100 μg) were resolved by one-dimensional SDS-PAGE and electrophoretically transferred to nitrocellulose filters, as previously described (Sirois and Richards 1992). Filters were incubated with anti-PGHS antibodies diluted in TBS (10 mM Tris-buffered saline, pH 7.5) containing 2% nonfat dry milk. [125I]-Protein A (1 × 106 cpm/ml TBS-2% milk) was used to visualize immunopositive proteins. Filters were washed three times (20 min/wash) in TBS-0.05% Tween and exposed to film at −70C.
Statistical Analysis
The coefficient of correlation between the intensity of PGHS-1 staining and the degree of malignancy (grade of adenocarcinomas) was determined to measure the strength of the relationship between the two variables. The coefficient was considered significant when the null hypothesis was rejected at p<0.05 (Student's t distribution). In addition, an analysis of variance was used to test the effect of grade of adenocarcinoma on PGHS-1 staining intensity. All values are given as mean ± SEM.
Results
Expression of PGHS Isoforms in Normal Human Ovaries
To determine if PGHS (PGHS-1 and/or PGHS-2) is expressed under physiological conditions, immunohis-tochemical staining was performed on normal human ovaries (n = 3). Results of the immunostaining showed that PGHS expression was restricted to two cell types in normal human ovaries. The first type was the surface epithelial cells covering the ovary, which displayed cytoplasmic staining (Figure 1A). Epithelial cells lining crypt-like structures also showed strong PGHS expression (Figure 1C). The second type of cells immunore-active for PGHS were endothelial cells of some blood vessels that showed cytoplasmic staining. The isoform expressed in these two cell types was determined to be PGHS-1. No PGHS-2 expression was detected in normal human ovaries (Figures 1B and 1D).
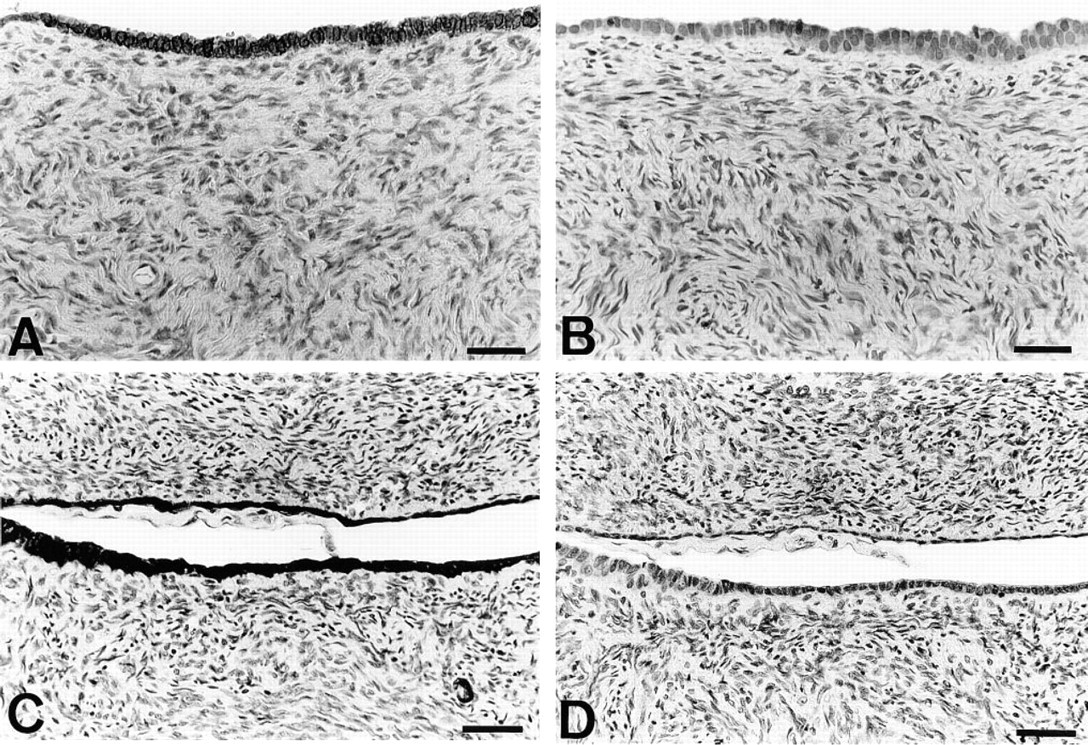
Expression of PGHS-1 by normal ovarian surface epithelium. Immunohistochemistry was performed on formalin-fixed sections of normal human ovaries as described in Materials and Methods. Epithelial cells covering the surface of the ovary (A) or lining crypt-like structures (C) were shown to express PGHS-1, because they displayed staining with anti-PGHS antibody 9181 but showed no immunoreactivity with PGHS-2-selective antibody MF243 (B.D). Counterstaining with Gill's hematoxylin. Ovaries from two different subjects are shown. Bars = 25 μm.
Immunohistochemical Expression of PGHS Isoforms in Human Ovarian Carcinomas
Of the 16 cases of malignant epithelial tumors studied, all showed some immunoreactivity for PGHS (Table 1). Different cell types in the tumors expressed PGHS, including epithelial tumor cells, fibroblasts, vascular endothelial cells, and vascular smooth muscle cells. Immunostaining with anti-PGHS-1 and anti-PGHS-2 antibodies showed that PGHS-1 was the only isoform expressed by the epithelial tumor cells (Figures 2A and 2B) as well as by the other cell types. Patterns of staining of tumor cells varied among different types of adenocarcinomas, with cells presenting either a staining mainly concentrated around the nucleus (Figure 2C) or, alternatively, a mostly diffuse cytoplasmic immunoreactivity (Figures 2A and 2D). Cytoplasmic staining was the most common pattern observed in adenocarcinomas of the serous type (Figure 2D), although some tumor cells also exhibited perinuclear immunoreactivity (Figure 2C). Intense staining concentrated around the nuclear membrane was commonly observed in the mucinous type of adenocarcinoma (Figure 2E). Occasionally, some epithelial cells of mucinous adenocarcinomas also presented a coarsly granular staining distributed throughout their cytoplasm (Figure 2A). The intensity of PGHS expression was heterogeneous (Table 1), being very light in some tumors and strong in others. No significant difference in PGHS-1 staining intensity was present (p > 0.05) among the tumors of different grades, with mean PGHS-1 staining of 2.5 ± 1.3 for tumors of borderline malignancy, 1.8 ± 0.3 for tumors of Grades I and II, and 2.3 ± 1.0 for tumors of Grade III. In addition, no correlation between the intensity of the immunostaining and the degree of malignancy (r = −0.030) of tumors could be established. Interestingly, immunostaining of one benign epithelial ovarian tumor (a serous cystadenoma) showed that the tumor epithelial cells strongly expressed PGHS-1 (Figure 2F). Control staining with normal rabbit serum was always negative (data not shown).
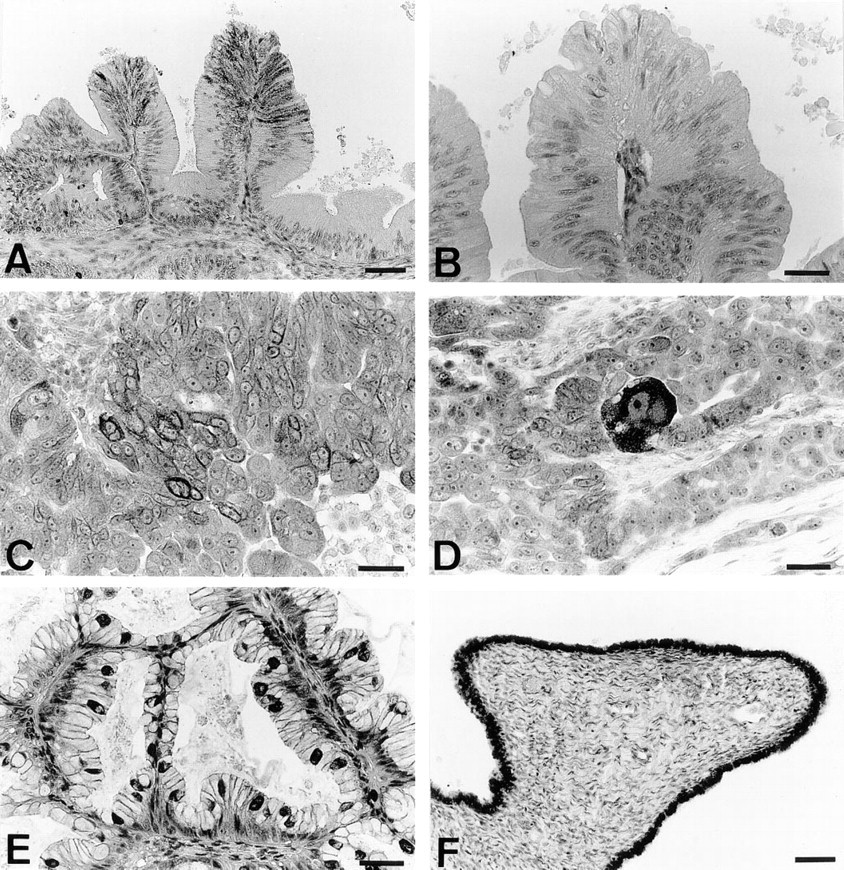
Expression of PGHS-1 by human ovarian adenocarcinomas. Immunohistochemistry was performed on formalin-fixed sections of 16 human ovarian adenocarcinomas as described in Materials and Methods. Immunostaining with anti-PGHS antibody 9181 revealed the presence of many PGHS-immunoreactive tumor cells (deep brown granular cytoplasmic staining) (A), whereas immunostaining with the anti-PGHS-2-selective antibody MF243 showed no reactivity (B). Different patterns of staining were observed among the tumors, including staining mostly concentrated around the nucleus (C,E) and diffuse cytoplasmic staining (A,D). A benign ovarian tumor (serous cystadenoma) exhibited intense expression of PGHS-1 (F). Bars = 25 μm.
Immunoblotting of PGHS Isoforms in Human Ovarian Carcinomas
Solubilized cell extracts were prepared from a serous adenocarcinoma and a mucinous adenocarcinoma (Patients 8 and 15, Table 1) and proteins analyzed by Western blotting using anti-PGHS antibodies. In both tumors, a 72,000 Mr band was visualized using antibody 9181 (recognizes both PGHS-1 and PGHS-2) and antibodies 8223 and MF241 (selective for PGHS-1) (Figure 3). No immunoreactive PGHS was detected with antibody MF243 (selective for PGHS-2) (Figure 3). The precise nature of the lower band (Mr 50,000) detected in Figure 3C (and to a lesser extent in Figure 3D) is unknown.
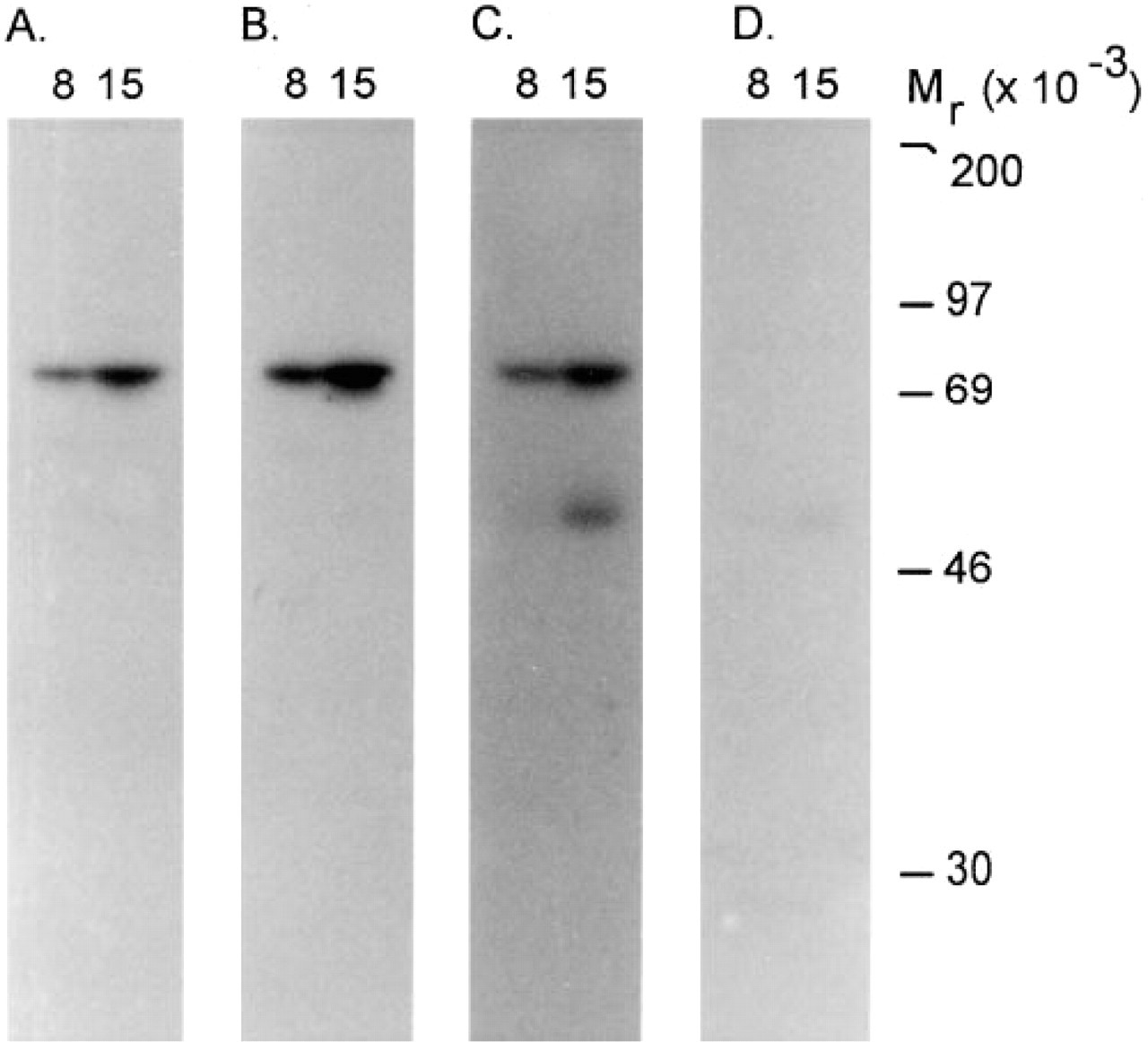
Expression of PGHS-1 by human ovarian adenocarcinomas. Solubilized cell extracts were prepared from a serous carcinoma (Patient 8, Table 1) and a mucinous cystadenocarcinoma (Patient 15, Table 1) and analyzed by one-dimensional SDS-PAGE and immunoblotting using specific anti-PGHS antibodies as described in Materials and Methods. Quadruplicate blots were probed with antibody 9181 (
Discussion
Our study demonstrates for the first time that PGHS, a key enzyme in the prostaglandin synthesis pathway, is expressed in human ovarian adenocarcinomas. Using antibodies specific to each isoform of PGHS (PGHS-1 or PGHS-2), we showed that PGHS-1 is present in ovarian adenocarcinomas. By immunohistochemistry, we determined that the enzyme was localized to the tumor epithelial cells and, in some tumors, to fibroblasts, vascular endothelial cells, and vascular smooth muscle cells. Interestingly, this study is first to document that the epithelium covering the surface of the ovary (from which ovarian adenocarcinomas are believed to be derived) expresses abundant amounts of PGHS-1. The physiopathological significance of PGHS-1 expression in normal surface epithelial cells and in ovarian adenocarcinomas is presently unknown.
PGHS-1 and PGHS-2 are two distinct but related enzymes possessing similar activities and generating similar eicosanoid products. Their primary structure is approximately 60% identical, as deduced from amino acid sequences of complementary DNAs (cDNAs) (Funk et al. 1991; Hla and Neilson 1992). PGHS-2 can be induced in a variety of cell types by a wide range of stimuli (Herschman 1994). Induction of PGHS-1 can also occur, but is not as extensive as PGHS-2 induction and not as widely observed (Herschman 1994). Development of PGHS-1- or PGHS-2-deficient mice by targeted gene disruption has resulted in different phenotypic changes (Dinchuk et al. 1995; Langenbach et al. 1995; Morham et al. 1995). PGHS-1-deficient mice showed no gastric pathology but had reduced platelet aggregation and a decreased inflammatory response to arachidonic acid (Langenbach et al. 1995). PGHS-2-deficient mice exhibited severe renal dysplasia, cardiac fibrosis, and infertility, as well as an altered inflammatory response (Dinchuk et al. 1995; Morham et al. 1995). These differences suggest that these two enzymes could play distinct roles in the organism (Williams and Dubois 1996).
In rat and bovine ovarian follicles, there is a selective induction of PGHS-2 after the endogenous surge of luteinizing hormone (Sirois and Richards 1992; Sirois 1994). The induction is granulosa cell-specific and occurs only in preovulatory follicles and not in follicles of a less advanced developmental stage. In this study, no PGHS-2 staining was observed in follicles present in normal ovarian samples (data not shown). This finding was expected because our samples contained no large preovulatory follicles.
Breast cancer as well as head and neck tumors have been shown to produce PGE2 (Watson and Chuah 1992; Klapan et al. 1993). Production of prostaglandins (PGE2) by colorectal cancers has also been reported (Rigas et al. 1993). The production by colorectal cancers probably results from PGHS-2 activity, because increased expression of PGHS-2 has recently been shown in human colorectal adenomas and adenocarcinomas (Eberhart et al. 1994; Kargman et al. 1995; Sano et al. 1995; Kutchera et al. 1996). Under in vitro conditions, intestinal epithelial cells overexpressing PGHS-2 demonstrate increased adhesion to the extracellular matrix and are resistant to induced apoptosis, two phenotypic changes that could enhance their tumorigenic potential (Tsuji and DuBois 1995). Experimental administration of sulindac sulfide, a nonsteroidal anti-inflammatory drug, to the Min strain of mice (mice with a mutation in the APC gene leading to the development of gastrointestinal adenomas) inhibits tumor formation and decreases small bowel PGHS-2 and prostaglandin E2 (Boolbol et al. 1996).
In addition to their effects on cellular adhesion and inhibition of apoptosis, prostaglandins have been implicated as possible modulators of the immune defenses against cancer. Suppression of immune functions by tumor-derived PGE2 and PGE2-producing suppressor macrophages may impair the destruction of tumor cells by the host defense (Young 1994). For example, PGE2 has been shown to block stimulation of T-cells to produce tumor necrosis factor-α (TNF-α) and lymphotoxin (TNF-β) (Ferreri et al. 1992) and to be suppressive to the cytotoxic activity of natural killer cells (Vaillier et al. 1992). Finally, eicosanoids might also contribute to carcinogenesis by increasing the proliferation rate of tumor cells, as recently shown with human colon carcinoma cells in vitro (Qiao et al. 1995), and by inducing angiogenesis (Hla et al. 1993).
Conversely, some prostaglandins are able to inhibit proliferation of tumor cells (Kikuchi et al. 1986, 1994). These prostaglandins have been shown to bind to nuclear target molecules, and to lead to cell cycle arrest of tumor cells at the G1-phase (Kikuchi et al. 1992). Interestingly, one of these prostaglandins, PGD2, can inhibit tumor growth and prolong the survival time of nude mice bearing human ovarian cancer cells (Kikuchi et al. 1986). Moreover, some of the antineoplastic prostaglandins have been shown to demonstrate adjuvant effects to cisplatin on human ovarian cancer cell growth, and their clinical use has been proposed as new treatment strategies for ovarian carcinoma (Kikuchi et al. 1992).
The significance of PGHS-1 expression by ovarian cancer cells can only be speculative at this time. However, the present study clearly establishes that the surface epithelium of the normal ovary and tumor cells in ovarian adenocarcinomas express large amounts of PGHS-1, a key rate-limiting enzyme in the prostaglandin biosynthetic pathway. Future studies will be needed to unravel the potential role(s) of prostaglandin synthesis in ovarian carcinogenesis.
Footnotes
Acknowledgements
Supported by grants from the Medical Research Council of Canada (JS) and the Fonds pour la Formation de Chercheurs et l'Aide à la Recherche (FCAR) (JS and MD).
We thank Drs Jilly F. Evans and Stacia Kargman (Merck Frosst Centre for Therapeutic Research; Pointe-Claire-Dorval, Québec) for kindly supplying antibodies MF241 and MF243.