
Abstract
Squamous cell carcinoma is one of the most common cancers in humans and is also a frequently diagnosed neoplasm in dogs. Induction of cyclo-oxygenase-2 (COX-2), a key rate-limiting enzyme in prostaglandin biosynthesis, has been implicated in the oncogenesis of various cancers in humans, including squamous cell carcinomas. However, expression of COX-2 has not been reported in spontaneous squamous cell carcinomas of non-human species. Canine squamous cell carcinomas share several similarities with the human disease. Therefore, the objective of this study was to determine whether COX isoenzymes were expressed in naturally occurring cases of squamous cell carcinomas in dogs. Canine normal skin (n=4) and squamous cell carcinomas (n=40) were studied by immunohistochemistry and immunoblotting analysis using polyclonal antibodies selective for COX-1 or COX-2. COX-2 was strongly expressed by neoplastic keratinocytes in all cases of squamous cell carcinomas, whereas no COX-2 was detected in normal skin and in the nonneoplastic skin and oral mucosa included in the tumor tissue samples (p < 0.01). Immunoblotting analysis confirmed the restricted expression of COX-2 (72,000–74,000 molecular weight doublet) in squamous cell carcinomas only. In contrast, faint COX-1 staining was found in normal skin and in squamous cell carcinomas. This study demonstrates for the first time that COX-2 is induced in canine squamous cell carcinomas, and provides a new model to investigate the role and regulation of COX-2 gene expression in naturally occurring squamous cell carcinomas.
Keywords
S
Prostaglandins are believed to play a role in the development and progression of some forms of cancer (Herschman et al. 1995; Subbaramaiah et al. 1997; DuBois et al. 1998; Taketo 1998; Rustgi 1998; Lipsky 1999; Ziegler 1999). The first rate-limiting step in the synthesis of all prostaglandins from arachidonic acid is controlled by the enzyme prostaglandin G/H synthase, also known as cyclo-oxygenase (COX) (Smith 1992; Funk 1993). COX is a homodimer composed of two subunits of about 70,000 Daltons and one heme group (van der Ouderaa et al. 1979; Kulmacz and Lands 1984). Two forms of cyclo-oxygenase have been identified, i.e., COX-1 and COX-2. COX-1 is constitutively expressed in many tissues and is believed to mediate physiological responses requiring a rapid and/or constant biosynthesis of prostaglandins. COX-2 is normally absent from cells but can be induced in response to inflammatory reactions, growth factors, and tumor promoters (Herschman 1996; Williams and DuBois 1996). The two isoforms share important similarities at the protein level, with about 60% identity in their amino acid sequences (Subbaramaiah et al. 1997). However, COX-1 and −2 are derived from distinct genes located on two different chromosomes and are encoded by messenger RNA (mRNA) of different sizes (about 2.8 vs 4.0 kb for COX-1 and −2, respectively) (Funk et al. 1991; Kujubu et al. 1991; Kosaka et al. 1994). The different responses of COX-1 and COX-2 reflect, in part, differences in the regulatory elements in the promoter of the two genes (Mestre et al. 1997).
Expression of COX-2 has been demonstrated in various human neoplasms, including colorectal, lung, gastric, pancreatic, prostate, breast, and hepatocellular adenocarcinomas, transitional cell bladder carcinomas, and adenocarcinomas and squamous cell carcinomas of the esophagus (Kargman et al. 1995; Parrett et al. 1997; Ristimaki et al. 1997; Wilson et al. 1998; Ratnasinghe et al. 1999; Tucker et al. 1999; Fosslien 2000; Gupta et al. 2000; Kömhoff et al. 2000). Recent studies have found that COX-2 is upregulated in vivo in human precancerous skin lesions such as actinic keratoses and in squamous cell carcinomas (Buckman et al. 1998; Müller-Decker et al. 1999). Chan et al. (1999) also reported that COX-2 mRNA and protein are increased in squamous cell carcinomas of the head and neck in humans. In vitro, ultraviolet (UV) B irradiation of human keratinocytes induced upregulation of COX-2 expression, suggesting the involvement of COX-2 in skin tumor development after prolonged sunlight exposure (Buckman et al. 1998; Isoherranen et al. 1999). Experimental administration of a selective inhibitor of COX-2 [celecoxib (SC-58635)] to hairless mice irradiated with UV light reduced tumor yield if the COX-2 inhibitor was administered during the irradiation period or prevented new tumor formation if the compound was administered after the onset of photocarcinogenesis (Fischer et al. 1999; Pentland et al. 1999). However, in contrast to experimental models of chemical- or radiation-induced skin tumors in rodents, the implication of COX-2 in spontaneous squamous cell carcinomas in animals has never been reported. Therefore, the objectives of the present study were to determine whether COX isoenzymes are expressed in canine squamous cell carcinoma and, if so, to identify which isoform is involved (COX-1 and/or COX-2) and to determine its cellular localization.
Materials and Methods
Materials
Diethyldithiocarbamic acid (DEDTC), octyl β-
Tissue Samples and Platelet Isolation
Forty cases of canine squamous cell carcinomas submitted to the Departement of Pathology and Microbiology of the Faculty of Veterinary Medicine (Université de Montréal) were included in the study. All cases were confirmed as squamous cell carcinomas by examination of hematoxylin–eosin–saffran-stained (HES) sections by a veterinary pathologist. The malignant nature of the squamous cell carcinomas was defined using criteria such as anaplasia (anisocytosis, pleomorphism, anisokaryosis), loss of normal polarity, anarchic growth, atypical mitotic figures, and local or vascular invasion. The histological classification used was the World Health Organization International Histological Classification of Tumors of Domestic Animals (Hall et al. 1976). Normal skin was obtained from four adult beagle dogs (aged 3–4 years old) euthanized for reasons unrelated to health problems. All tissues studied were fixed in 10% neutral buffered formalin, whereas samples from normal skin and a squamous cell carcinoma were frozen at –70C for immunoblotting analysis. Canine platelets were isolated as previously described (Tremblay et al. 1999).
Anti-COX Antibodies
Two anti-COX antibodies (antibodies 8223 and MF243) were used. Affinity-purified polyclonal antibody 8223 was raised in rabbits against ovine COX-1 and was shown to be selective for COX-1 in various species (Wimsatt et al. 1993; Sirois 1994; Sirois and Doré 1997; Doré et al. 1998; Tremblay et al. 1999). Antibody MF243 was generously provided by Drs. Jilly F. Evans and Stacia Kargman (Merck Frosst Centre for Therapeutic Research; Pointe-Claire-Dorval, PQ, Canada). MF243 was raised in rabbits against ovine placental COX-2 and its selectivity for COX-2 has previously been characterized (Kargman et al. 1995; Tremblay et al. 1999).
Immunohistochemistry
Immunohistochemical staining was performed using the Vectastain ABC kit as previously described (Doré et al. 1993). Briefly, formalin-fixed tissues were paraffin-embedded and 3 μm-thick sections were prepared and deparaffinized through graded alcohol series. Endogenous peroxidase was quenched by incubating the slides in 0.3% hydrogen peroxide in methanol for 30 min. After rinsing in PBS for 15 min, sections were incubated with diluted normal goat serum for 20 min at room temperature (RT). Primary antibodies diluted in PBS were applied (8223 at 1:100 dilution and MF243 at 1:7500 dilution) and sections were incubated overnight at 4C. Control sections were incubated with PBS or with non-immune rabbit serum. After rinsing in PBS for 10 min, a biotinylated goat anti-rabbit antibody (1:222 dilution; Vector) was applied, and sections were incubated for 45 min at RT. Sections were washed in PBS for 10 min and incubated with the avidin DH–biotinylated horseradish peroxidase H reagents for 45 min at RT. After a PBS wash for 10 min, the reaction was revealed using diaminobenzidine tetrahydrochloride (DAB) as the peroxidase substrate. Sections were counterstained with Gill's hematoxylin stain and mounted. Immunoreactivity was evaluated in a blinded fashion by two independent observers, using a grading system where 0 = 0%, 1 = <10%, 2 = 10–30%, 3 = 31–60%, and 4 = >60% of positive cells. The intensity of COX-2 immunoreactivity was graded from — = no, + = weak, + + = moderate, and + + + = strong staining.
Solubilized Cell Extracts and Immunoblotting Analysis
Solubilized cell extracts were prepared as previously described (Sirois and Doré 1997). Briefly, tissues were homogenized on ice in TED buffer [50 mM Tris (pH 8.0), 10 mM ethylenediamine tetra-acetic acid (EDTA), and 1 mM diethyldithiocarbamic acid (DEDTC)] containing 2 mM octyl glucoside and were centrifuged at 30,000 × g for 1 hr at 4C. The crude pellets (membranes, nuclei, and mitochondria) were sonicated (8 sec/cycle; three cycles) in TED sonication buffer [20 mM Tris (pH 8.0), 50 mM EDTA, and 0.1 mM DEDTC] containing 32 mM octyl glucoside. The sonicates were centrifuged at 16,000 × g for 15 min at 4C. The recovered supernatant (solubilized cell extract) was stored at –70C until electrophoretic analyses were performed. The protein concentration was determined by the method of Bradford (Bio-Rad Protein Assay). Proteins were resolved by one-dimensional SDS-PAGE and electrophoretically transferred to nitrocellulose filters, as described (Sirois and Doré 1997). Filters were incubated with anti-COX antibodies diluted in TBS (10 mM Tris-buffered saline, pH 7.5) containing 2% non-fat dry milk. [125I]-protein-A (1 × 106 cpm/ml TBS–2% milk) was used to visualize immunopositive proteins. Filters were washed three times (10 min/wash) in TBS–0.05% Tween-20 and exposed to film at –70C.
Statistical Analysis
The Fisher's exact test was used to compare the frequency of COX-2 expression between normal skin and skin squamous cell carcinomas. Statistical analyses were performed using the JMP Software (SAS Institute; Cary, NC).
Results
Characteristics of Dogs with Squamous Cell Carcinomas
The age of dogs with squamous cell carcinomas ranged from 3.5 to 12.8 years (mean 8.4 years). Eighty-five percent of dogs (34/40 cases) were more than 6 years old at the time of cancer diagnosis, which is more than 40 years old when expressed in equivalent human years (Reeve and DeForest 1988). Of the 40 cases, 25 animals (62.5%) were large-breed dogs (Table 1). There was no sex predilection, as 22 animals (55%) were females and 18 (45%) were males (Table 1). The majority of tumors (24/40, 60%) were located on the feet, whereas nine cases (22.5%) originated in the skin and seven (17.5%) were located in the oral cavity.
COX Expression in Normal Canine Skin and Oral Mucosa
To determine whether COX-1 and/or COX-2 was expressed under physiological conditions, immunohistochemical staining was performed on normal canine skin (n=4). Results showed that no COX-1 was present in the epidermis or in the follicle walls (Figure 1A), whereas some COX-1 staining was found in smooth muscle cells of arrector pili muscles (Figure 1C). However, no COX-2 expression was detected in normal canine skin (Figure 1B). Moreover, no COX-2 immunostaining was found in the non-neoplastic skin included in the tumor tissue samples (26/33 cases) and in the non-neoplastic oral mucosa included in the tumor samples (5/7 cases) (Figure 1D).
Induction of COX-2 in Canine Squamous Cell Carcinomas
In contrast to normal skin and oral mucosa in which no COX-2 expression was found, all cases of squamous cell carcinomas (100%) displayed immunoreactivity for COX-2. The extent and intensity of the COX-2 staining varied among the tumors (Table 1). There was no correlation between site, grade, and/or COX-2 intensity. COX-2 immunostaining was predominantly localized in the cytoplasm of neoplastic keratinocytes (Figures 2A-2C), often being concentrated around the nuclear membrane (Figure 2C). In a small number of tumors (n=3/40), fusiform fibroblast-like cells located in the stroma were COX-2-positive (Figure 2D). In the majority of carcinomas (n=35/40), neoplastic keratinocytes also displayed very weak positive staining for COX-1 (Figure 2E).
Characteristics of specimens from dogs with squamous cell carcinoma
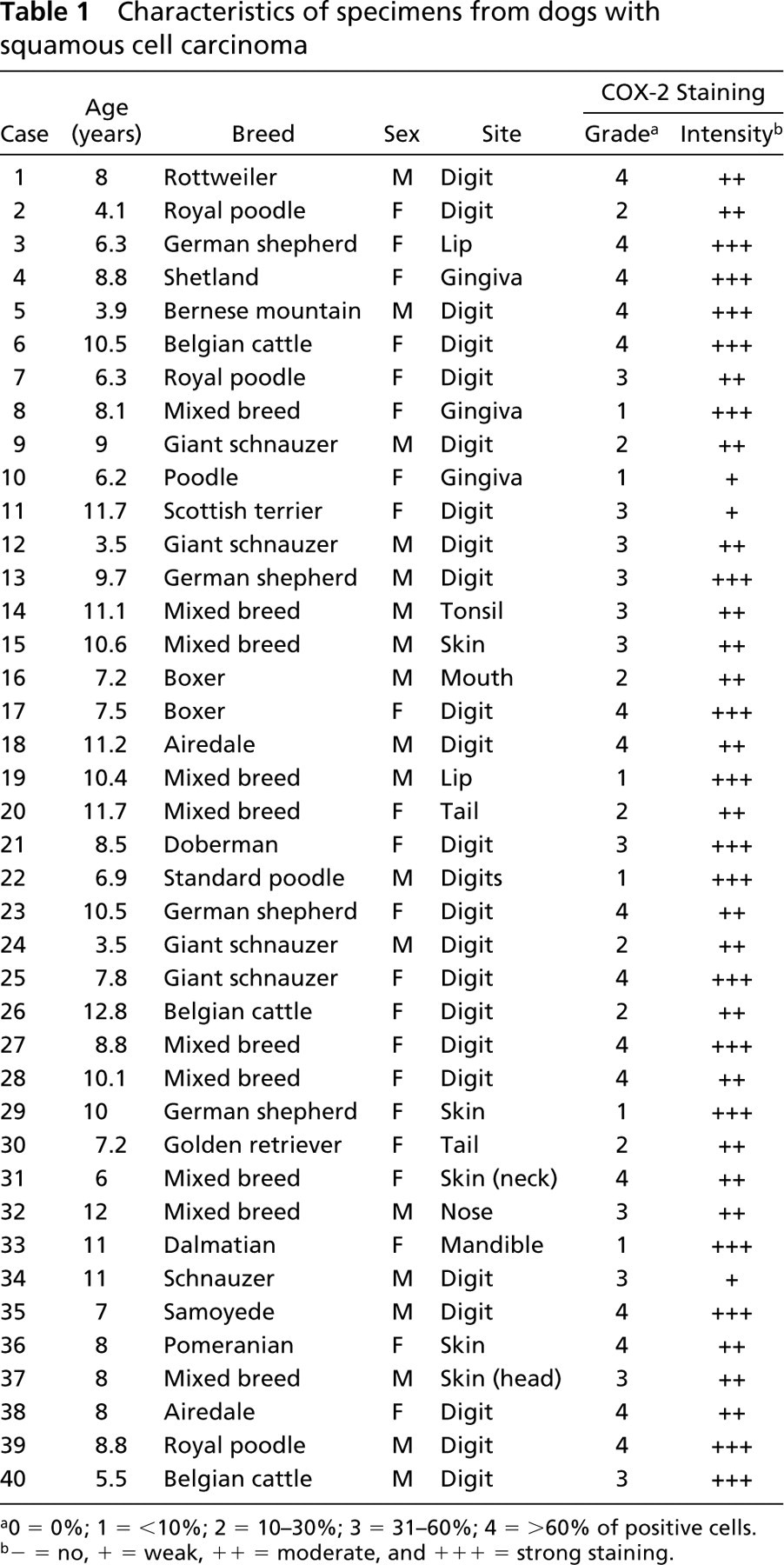
a0 = 0%; 1 = <10%; 2 = 10–30%; 3 = 31–60%; 4 = >60% of positive cells.
b− = no, + = weak, ++ = moderate, and + + + = strong staining.
Immunoblotting of COX Isoforms in Canine Tissues
To confirm the specificity of each antibody and to characterize each COX isoform, solubilized cell extracts were prepared from normal skin, a squamous cell carcinoma, and canine platelets, and proteins were analyzed by Western blotting. When a selective anti-ovine COX-1 antibody was used, a very faint 69,000 Mr band was detected in the squamous cell carcinoma (Figure 3A). A band of identical molecular weight was detected in canine platelets (Figure 3A) and therefore corresponded to canine COX-1. When a selective anti-ovine COX-2 antibody was used, no signal was detected in normal skin but strong COX immunoreactivity was observed in the squamous cell carcinoma (Figure 3B). Canine COX-2 appeared as a 72,000–74,000 Mr doublet and a small 62,000 Mr band (Figure 3B) believed to correspond to a proteolytic fragment, as previously observed in other species (Sirois and Richards 1992; Sirois 1994; Sirois and Doré 1997). The absence of detectable COX-2 in canine platelets is in keeping with reports in other species (DuBois et al. 1998).
Discussion
This study is the first to document the expression of COX-2 in naturally occurring squamous cell carcinomas in animals. Interestingly, whereas no or very low expression of the enzyme was observed in normal skin, all cases of canine squamous cell carcinomas examined expressed COX-2. COX-2 was abundantly present in neoplastic keratinocytes and in stromal cells within some tumors. Tumors originated from different anatomic locations (i.e. mouth, feet, or skin), with no predilection of COX-2 expression for a specific site because all cases contained immunoreactive cells. In humans, squamous cell carcinomas from different anatomic sites, including the skin, head, and neck, the lung, and the esophagus have also been shown to overexpress COX-2 (Wolff et al. 1998; Chan et al. 1999; Müller-Decker et al. 1999; Ratnasinghe et al. 1999). The high prevalence of COX-2-positive tumors in dogs is also in agreement with a recent report in humans in which all tumors (10/10 cases) were shown to express COX-2 (Chan et al. 1999). The perinuclear concentration of the enzyme in some cancer cells is in keeping with the phenotype reported in previous studies (Morita et al. 1995; Subbaramaiah et al. 1997). The mean age of 8.4 years of affected dogs in this study corresponds to approximately 50 years for humans, suggesting that, as in humans, spontaneous squamous cell carcinoma in dogs is influenced by age.
Among the etiologic factors contributing to skin cancer, ultraviolet (UV) irradiation plays a central role in the development of cutaneous squamous cell carcinomas in humans, in part through its ability to induce prostaglandin synthesis (Grewe et al. 1993; Buckman et al. 1998). An association between UV, solar dermatosis, and squamous cell carcinomas has also been suggested in dogs in a study in which animals exposed to the long-term effects of low-level ionizing radiation displayed an increased risk for development of solar dermatosis and squamous cell carcinomas (Nikula et al. 1992). Two recent studies have demonstrated that irradiation of cultured human keratinocytes or of human subjects causes upregulation of COX-2 protein, suggesting a role for COX-2 in prostaglandin synthesis in response to acute UVB irradiation (Buckman et al. 1998; Isoherranen et al. 1999). Specific inhibitors of COX-2 have also been shown to reduce UV-induced skin tumor development in hairless mice (Fischer et al. 1999; Pentland et al. 1999). It is possible that UVB irradiation contributes to the increased COX-2 expression through its action on the tumor suppressor gene p53. Indeed, p53 has recently been shown to cause a marked decrease in the expression of COX-2 in mouse fibroblasts (Subbaramaiah et al. 1999). Because UVB irradiation of mouse skin induces mutations of the p53 gene, the presence of mutant forms of p53 in malignant tissues could contribute to the high levels of COX-2. Interestingly, mutations of the p53 gene have been reported in natural cases of squamous cell carcinomas in dogs (Gamblin et al. 1997; Mayr et al. 1999).
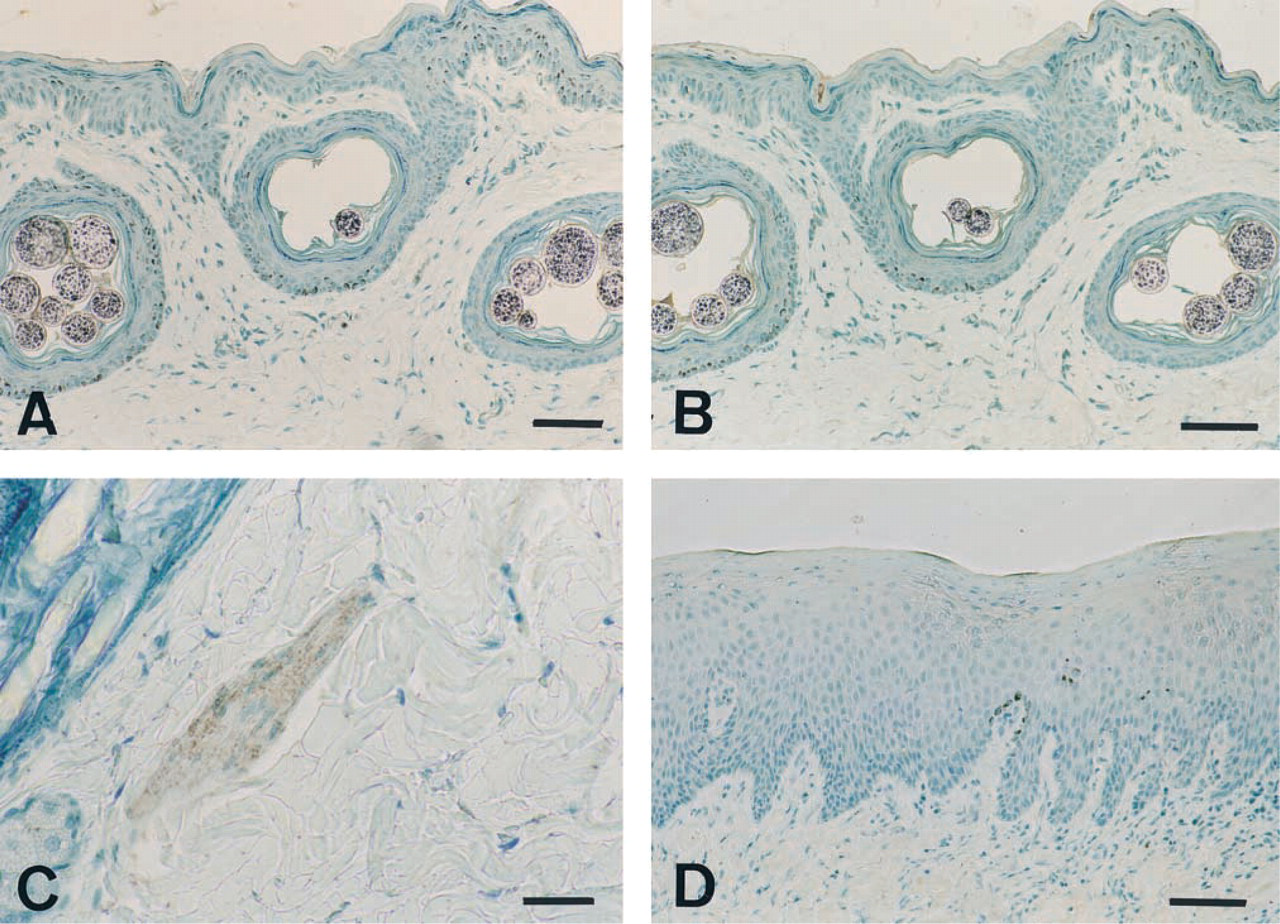
Expression of COX-1 and COX-2 by normal canine skin and oral mucosa. Immunohistochemistry was performed on formalin-fixed sections of normal canine skin and oral mucosa as described in Materials and Methods. Immunostaining with antibody 8223 (selective for COX-1) demonstrated no COX-1 in the epidermis or in follicle walls (
Overexpression of COX-2 has also been reported in various experimental models of skin carcinogenesis (Müller-Decker et al. 1995, 1998; Maldve and Fischer 1996; Scioscia et al. 1997; Matsuura et al. 1999; Nishimura et al. 1999). Papillomas and carcinomas induced in mice by a procedure of initiation–promotion were found to overexpress COX-2, and administration of a specific COX-2 inhibitor suppressed papilloma development and retarded malignant progression in the same murine model of skin carcinogenesis (Müller-Decker et al. 1995, 1998). COX inhibitors such as piroxicam and nabumetone have also been reported to inhibit tumor growth and decrease tumor weight in mice inoculated SC with cells from squamous cell carcinomas of the head and neck (Scioscia et al. 1997). In addition, a selective COX-2 inhibitor (JTE-522) was shown to suppress tumor growth in nude mice injected with human head and neck squamous cell carcinoma cells (Nishimura et al. 1999). Stimulation of cultures of human or murine keratinocytes by agonists such as interferon-γ, transforming growth factor-α, or 12–0-tetradecanoylphorbol-13-acetate (TPA) resulted in the induction of COX-2 mRNA or protein (Maldve and Fischer 1996; Matsuura et al. 1999). In contrast, interferon-γ was ineffective in inducing COX-2 expression in squamous cell carcinoma cell lines, suggesting alterations in the IFN-γ response pathway in cancerous keratinocytes (Matsuura et al. 1999).
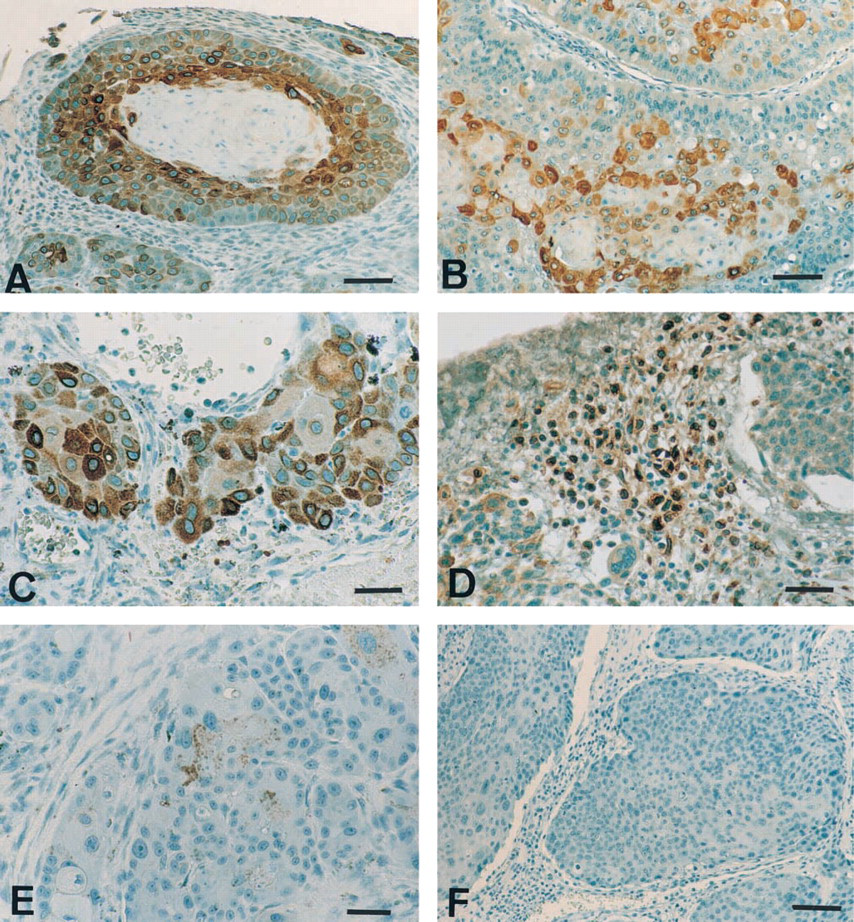
Expression of COX-1 and COX-2 by canine squamous cell carcinomas. Immunohistochemistry was performed on formalin-fixed sections of canine squamous cell carcinomas as described in Materials and Methods. Immunostaining with antibody MF243 (selective for COX-2) revealed the presence of COX-2 in neoplastic keratinocytes (
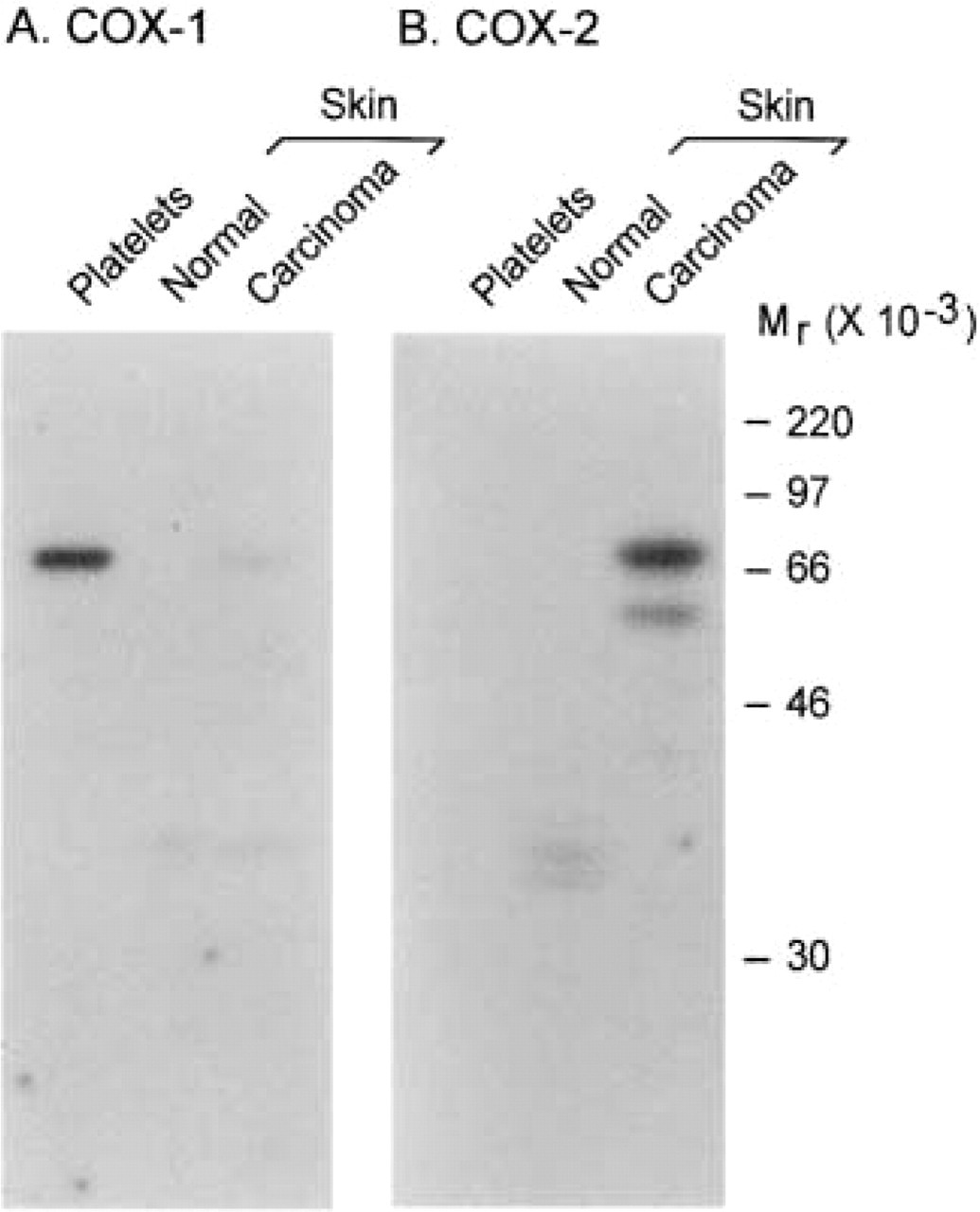
Immunoblotting analysis of COX-1 and COX-2 isoforms in canine skin tissues. Solubilized cell extracts were prepared from normal skin, a squamous cell carcinoma (Case 39; Table 1), and canine platelets, and were analyzed by one-dimensional SDS-PAGE and immunoblotting as described in Materials and Methods. Duplicate blots were probed with antibody 8223 (selective for COX-1,
The precise mechanisms by which COX-2 expression contribute to carcinogenesis are beginning to unravel. These include effects on cell proliferation, apoptosis, cell invasion, and immune response (Fischer 1997). In humans and rats, prostaglandins appear to be required for normal cell proliferation of skin keratinocytes (Fischer 1997). A recent study reported that the enhanced expression of COX-2 in a human skin epidermal cancer cell line compared with a normal keratinocyte cell line played a role in cell growth (Higashi et al. 2000). In the latter study, suppression of COX-2 expression with antisense oligonucleotides resulted in significant inhibition of cell growth. Prostaglandins have also been reported to contribute to the growth of some but not all colon cancers (Fischer 1997). Inhibition of apoptosis also appears to contribute to the carcinogenesis of COX-2 expressing cancers (Fischer 1997). Indeed, Tsujii and DuBois (1995) have shown that intestinal epithelial cells permanently transfected with a COX-2 expression vector were resistant to apoptosis. Subbaramaiah et al. (1999) speculated that the mechanisms underlying this inhibition of apoptosis could include an effect on nucleobindin, an apoptosis-associated protein, or the modulation of intracellular arachidonic acid. Enhanced angiogenesis and increased invasiveness have also been observed in cancer cells overexpressing COX-2 (Fosslien 2000). Williams et al. (2000) recently showed that tumor growth was markedly attenuated when Lewis lung carcinoma cells were implanted into mice genetically deficient in COX-2. Moreover, a decrease in vascular density was observed in tumors grown in these COX-2-deficient mice, suggesting that host-derived COX-2 could regulate intratumor vascular density (Williams et al. 2000). High levels of some prostaglandins (PGE2) are also known to suppress immune surveillance and to impair killing of malignant cells (Fischer 1997). Administration of indomethacin, a nonselective COX inhibitor, to patients with squamous cell carcinomas of the head and neck resulted in a significant increase in the number of infiltrating CD2+ lymphocytes in the tumors (Cross et al. 1992). These results suggest that production of prostaglandins contributes to inhibition of cellular immunity in this type of cancer.
Evidence for the role of COX-2 in tumor progression in humans also comes from promising results obtained with trials of therapy aimed at the specific inhibition of COX-2 expression in colon, bladder, and skin cancer (Ziegler 1999). Interestingly, a Phase I trial of piroxicam in dogs with naturally occurring tumors resulted in a partial remission of the disease in three of five dogs with squamous cell carcinoma (Knapp et al. 1992). Piroxicam has also been shown to be effective in the treatment of transitional cell carcinoma of the urinary bladder in dogs, a form of cancer in which overexpression of COX-2 by tumor cells has been reported in both humans and dogs (Knapp et al. 1994; Mohammed et al. 1999; Khan et al. 2000). These results suggest that strategies directed at the inhibition of COX-2 represent a potentially effective means to treat squamous cell carcinomas.
In summary, squamous cell carcinomas share many similarities between humans and dogs, including an increased prevalence with advancing age, an absence of breed or race predilection, and the presence of tumors arising from a variety of anatomic sites. The present study further strengthens the parallel between the two species by establishing that, as in humans, induction of COX-2 occurs in most if not all squamous cell carcinomas in dogs. It should also be considered that, as companion animals, dogs share with humans a similar environment and may be exposed to the same external factors that contribute to cancer. In view of all these similarities, we propose the dog as a valuable model to study the role and regulation of COX-2 gene expression in naturally occurring cases of squamous cell carcinoma.
Footnotes
Acknowledgements
Supported by grants from the Morris Animal Foundation (MD and JS), the Natural Sciences and Engineering Council of Canada (MD), and the Canadian Institutes for Health Research (CIHR) (JS). JS is the recipient of a CIHR Investigator Award.
We thank Drs Jilly F. Evans and Stacia Kargman (Merck Frosst Centre for Therapeutic Research; Pointe-Claire-Dorval, Québec) for kindly providing antibody MF243.