
Abstract
The relaxed conformation of plasminogen activator inhibitor-2 (PAIr) is formed during inactivation of the matrix-degrading enzyme urokinase plasminogen activator (uPA). The presence of PAIr in tissues, therefore, indicates the in situ inhibition of uPA-mediated proteolysis. In addition, PAIr functions as a ligand for the clearance receptor low-density lipoprotein receptor-related protein (LRP), thereby promoting internalization of receptor-bound uPA-PAIr complexes from the cell surface. The rapid internalization of receptor-bound, inactivated uPA has been suggested to be characteristic of invasive cell phenotypes. The aims of this study were to characterize the immunohistochemical localization of PAIr in human term gestational tissues (amnion, choriodecidua, and placenta) and to establish its co-expression with other components of the uPA cascade. The results obtained indicate that PAIr immunoreactivity was exclusively localized to amnion epithelial cells, with only minimal staining in the underlying chorion. PAIr immunoreactivity was not detectable in any of the trophoblastic tissues examined (villous and extravillous). The tissue-specific expression of PAIr immunoreactivity was not significantly altered in association with labor onset. uPA and PAI-2 staining was localized predominantly to amnion epithelial cells, underlying chorion, and trophoblast cells of villous and extravillous tissue. Amnion and trophoblasts of extravillous and chorionic tissue showed uPAR immunoreactivity, whereas staining in placenta was absent. Immunoreactive LRP was confined to trophoblasts of the chorion, and the villous and extravillous tissue. For the first time, localization of PAIr at the tissue level has been identified. The data obtained are consistent with the hypothesis that cells of invasive phenotype, although expressing all components of the uPA cascade, do not accumulate immunoreactive PAIr, because it is rapidly internalized from the cell surface. Conversely, cells of noninvasive phenotype will accumulate PAIr immunoreactivity only in the absence of LRP expression. We propose that the presence of PAIr and the absence of LRP at the cell surface are putative markers of noninvasive phenotypes.
One of the principal regulators of extracellular uPA activity is plasminogen activator inhibitor-2 (PAI-2) (Gleeson et al. 1992; Blasi 1993). This inhibitor complexes with and inactivates uPA, thus limiting ECM degradation (Baker et al. 1990). Recently, it has been established that during complex formation and inactivation of uPA, PAI-2 undergoes a stable conformational change in which an exposed reactive site loop is cleaved and inserted into the central β-sheet (Shore and Day 1995; Wilczynska et al. 1995). The structurally rearranged PAI is referred to as the relaxed conformation (PAIr). The uPA-PAIr complex contains a structural motif that binds with high affinity to the multifunctional clearance receptor low-density lipoprotein receptor-related protein (LRP) (Cubellis et al. 1990; Nykjaer et al. 1992; Andreasen et al. 1994). Once formed, the uPA-PAIr complex is very rapidly internalized to endosomes by LRP and is subsequently degraded in lysosomes (Jensen et al. 1990). By this mechanism, receptor-bound, inactivated uPA is rapidly removed from the cell surface and dissociated from uPAR (Conese et al. 1995). uPAR is recycled to the cell surface, thus sustaining focused proteolytic activity (Herz et al. 1992). In support of a role for LRP in regulation of cell surface expression of uPA activity, disruption of the LRP gene has been reported to compromise physiological processes that involve cell migration and the activity of the PA cascade (Herz et al. 1992). Therefore, it has been proposed that the rapid internalization of receptor-bound, inactivated uPA is a determinant of an invasive phenotype of a cell.
Recently, we have developed and characterized a monoclonal antibody that recognizes an epitope of the relaxed conformation of PAI-2 (McPhun et al. and Saunders et al., submitted for publication). For the first time, it is therefore possible to identify sites of PAIr formation at the tissue level. To further investigate the expression of immunoreactive PAIr and its co-expression with other PA cascade components, we have characterized the tissue-specific localization of immunoreactive PAIr, PAI-2, LRP, uPA, and uPAR in the same samples of human term gestational tissues.
Human term gestational tissues express all components of the PA cascade and undergo extensive ECM remodeling at the time of labor and delivery (Feinberg et al. 1989; Granstrom et al. 1989,1991,1992; Kliman and Feinberg 1990; Hofmann et al. 1994; Kliman 1994). It has been suggested that localized degenerative changes in the amnion and chorion may be sufficient to reduce tensile strength and hence mediate fetal membrane rupture at term (Vadillo-Ortega et al. 1995). Similar proteolytic mechanisms may operate in the placenta, facilitating its separation from maternal tissues after labor. These tissues therefore represent a useful model for investigating the expression of immunoreactive PAIr and the co-expression of uPA cascade components.
Materials and Methods
Tissue Collection
Amnion, choriodecidua and placenta were collected from women at 37–40 weeks' gestation before (at Caesarean section, not in labor; n = 3), during (at Caesarean section, in labor; n = 3), and after (spontaneous-onset labor, normal vaginal delivery; n = 3) labor. Tissue was snap-frozen in optical cutting temperature (OCT) compound (Tissue-Tek; Miles, Elkhart, IN) by immersion in isopentane precooled on dry ice. Samples were stored at −80C until processed. Informed consent was obtained from all patients and the study was approved by the Research and Ethical Committees of the Royal Women's Hospital, Melbourne, Australia.
Immunohistochemistry
Immunostaining was performed using the peroxidase-anti-peroxidase method (DAKO LSAB + HRP kit; DAKO, Glostrup, Denmark). Briefly, 5-μm-thick cryostat sections were cut and mounted on poly-
Immunohistochemical staining levels of plasminogen activation cascade components in human gestational tissues a
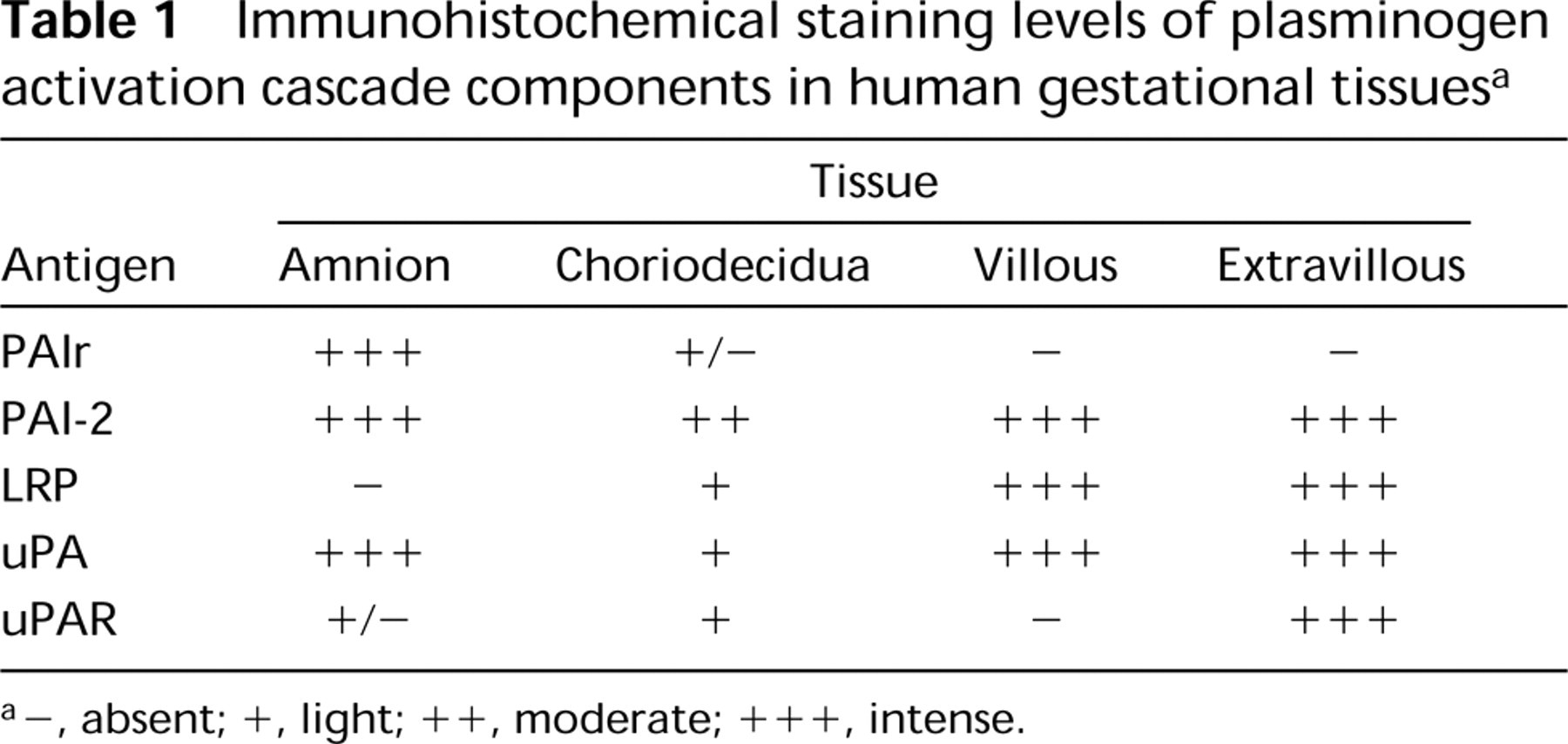
a -, absent; +, light; + +, moderate; + + +, intense.
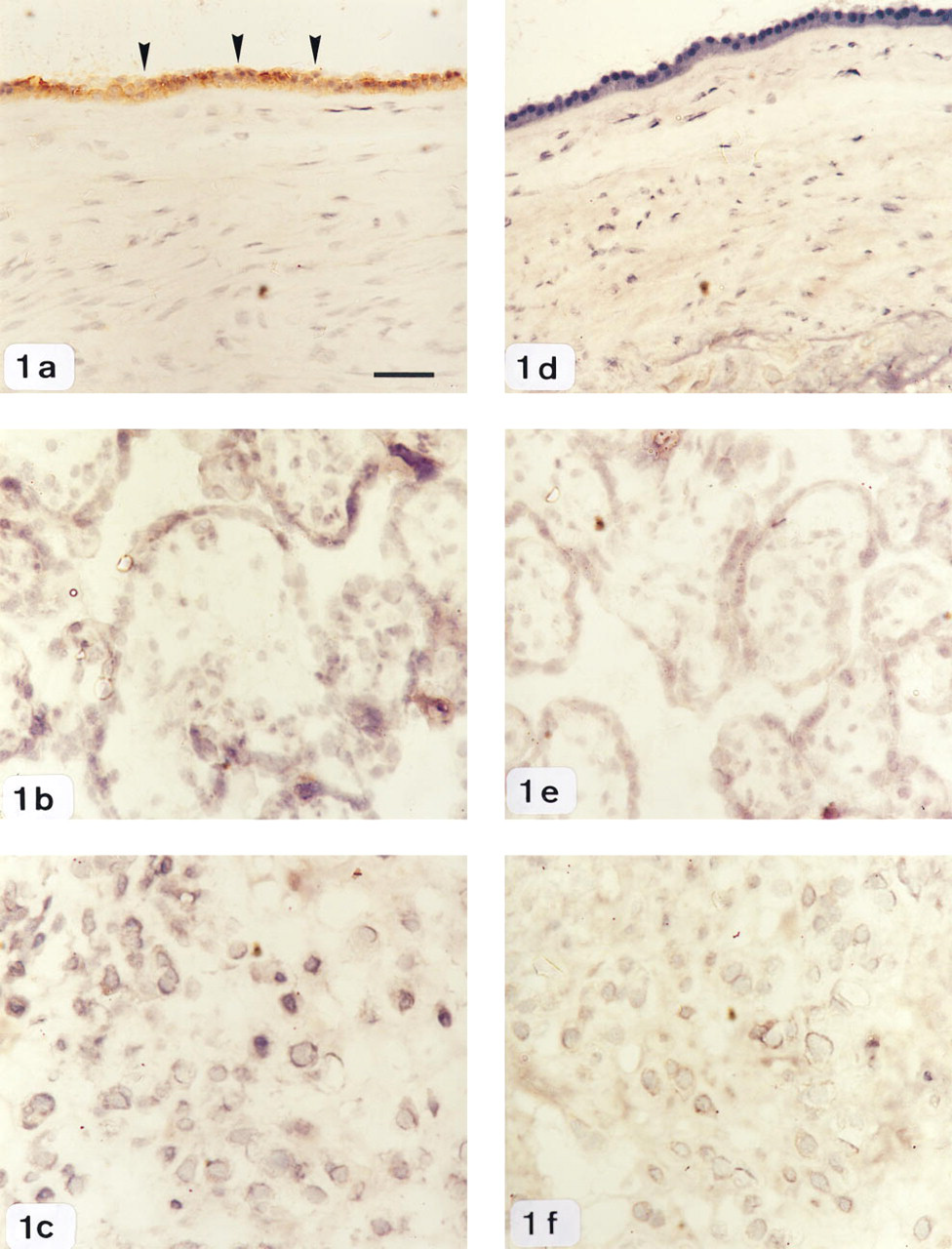
Immunohistochemical localization of the relaxed conformation of PAI-2 in human gestational tissues.
Negative control experiments for all antisera included (a) omission of primary antibody, (b) substitution of primary antibody with a monoclonal antibody of irrelevant specificity, and (c) substitution of primary antibody with normal nonimmune immunoglobulins of the same isotype at the equivalent protein concentration. In addition, preabsorption with available antigen for each of the uPA and PAI-2 antibodies was performed as a further control.
Assessment of Staining
Immunostaining was assessed semiquantitatively as the percentage of positively stained cells according to the following scale: 0, absent (no staining present); 1, light (1–30%); 2, moderate (30–70%); and 3, intense (70–100%). The sections were scored by two independent observers and the average data represent a consensus value of all observations.
Western Blot Analyses
Human placental, chorion, and amnion tissue (1 g) were homogenized in 50 mM glycine and 0.5% Triton X-100 and centrifuged at 6000 rpm for 15 min at 4C to sediment any insoluble material. The supernatant was assayed immediately for protein concentration using the Pierce BCA Protein Assay kit (Pierce; Rockford, IL).
Fifty μg of tissue protein was diluted in SDS-PAGE loading buffer (100 mM Tris-HCl, pH 6.8, 200 mM β-mercapto-ethanol, 4% SDS, 0.2% bromophenol blue, 20% glycerol) and boiled for 5 min. Samples were size-fractionated on an 8% SDS-PAGE gel under reducing conditions (except for LRP, which was electrophoresed on a 6% SDS-PAGE gel under nonreducing conditions). The molecular weight of protein samples was estimated by comparison with a 250- to 30-kD protein standard (Gibco BRL; Grand Island, NY) run in parallel on all gels. Proteins were transferred to a nitrocellulose membrane (Schleicher and Scheull; Dassell, Germany) using a Semi-Phor semidry transfer unit (Hoefer-Scientific Instruments; San Francisco, CA). Nonspecific binding sites were blocked by incubating transfers in blocking buffer (5% nonfat dry milk, 50 mM Tris-HCl, 150 mM NaCl) at RT for 1 hr. The membranes were then incubated overnight at 4C in blocking buffer containing one of the aforementioned antibodies. The membranes were washed in 0.2% Tween 20/PBS and incubated with a horseradish peroxidase-conjugated secondary antibody in blocking buffer for 1 hr. After appropriate washing, antibody bound to antigen was detected by ECL (Amersham; Arlington Heights, IL) and autoradiography.
Results
Expression of Plasminogen Activation Cascade Components in Human Gestational Tissues
The tissue-specific expression of immunoreactive PA cascade antigens (PAIr, PAI-2, LRP, uPA, uPAR) in human gestational tissues (amnion, choriodecidua, and placenta) obtained at term was determined independently by two observers, using qualitative (tissue distribution) and semiquantitative (percent immunoreactive cells) procedures. Tissue-specific expression did not change in association with labor status (determined qualitatively or semiquantitatively; p <0.1; n = 3 per group). Therefore, the data obtained from these groups were pooled in all subsequent analyses (summarized in Table I).
Relaxed Conformation of PAI-2
Intense immunostaining for the relaxed conformation of PAI-2 was localized to amnion epithelial cells (Figure 1). In contrast, PAIr immunoreactivity was not detectable in cells of the chorion and of the villous and extravillous tissue (Figures 1a–1c), although occasional staining in the fetal blood vessels of the placental villi was observed (data not shown). Immunostaining was not detected in negative control sections (Figures 1d–1f).
Identification of PAI-2 protein by immunohistochemistry.
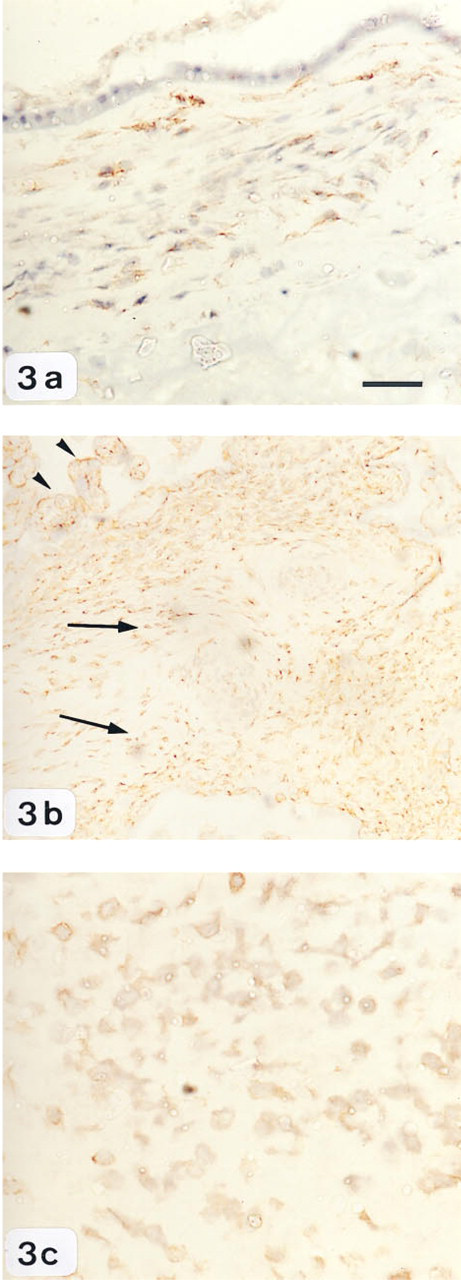
LRP detected in frozen sections of human gestational tissues by immunohistochemistry.
Plasminogen Activator Inhibitor Type-2
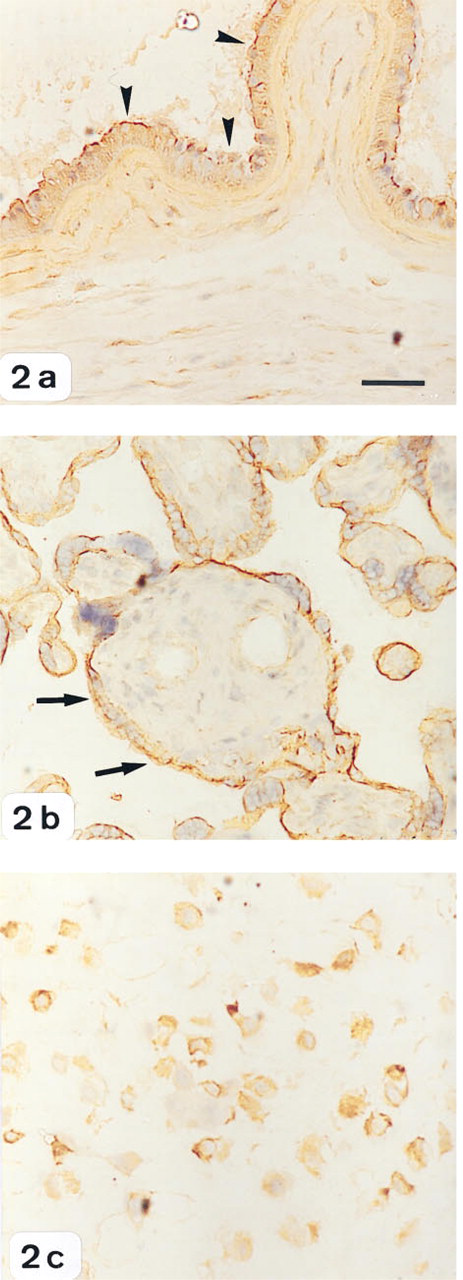
Positive staining for PAI-2 protein was present in amnion epithelium, chorion, and in villous and extravillous trophoblast cells. There were no labor-associated differences in the staining intensity of PAI-2 in any of the gestational tissues examined. Maximal staining was detected in the amnion, placental villi, and extravillous tissue (Figures 2a–2c), whereas chorionic cells showed only moderate PAI-2 immunoreactivity (Figure 2a). No staining was observed in negative control sections.
Low-density Lipoprotein Receptor-related Protein
LRP staining was localized predominantly in chorionic cells and trophoblasts of both villous and extravillous tissue, and did not change with labor onset (Figures 3a–3c). At the fetomaternal interface, intense LRP staining was detected in the trophoblast cell columns anchoring in the maternal tissue and in trophoblast cells lining the placental villi, whereas only light staining was detected in the cells within the chorion. In contrast, LRP immunoreactivity was not detectable in the amnion epithelium (Figure 3a). Immunostaining without primary antibody or its substitution with irrelevant or nonimmune immunoglobulins was also negative.
Urokinase Plasminogen Activator
uPA staining was localized predominantly in amnion epithelium, underlying chorion, and trophoblasts of both villous and extravillous tissue, and did not show any change in intensity with respect to the different stages of labor studied (Figures 4a–4c). The cytoplasm of amnion epithelium and of the villous and extravillous trophoblasts stained intensely compared with cells of the chorion, in which only minimal staining was observed. In control experiments, no immunostaining was observed when the primary antibody was omitted or substituted with nonimmune immunoglobulins.
Urokinase Receptor
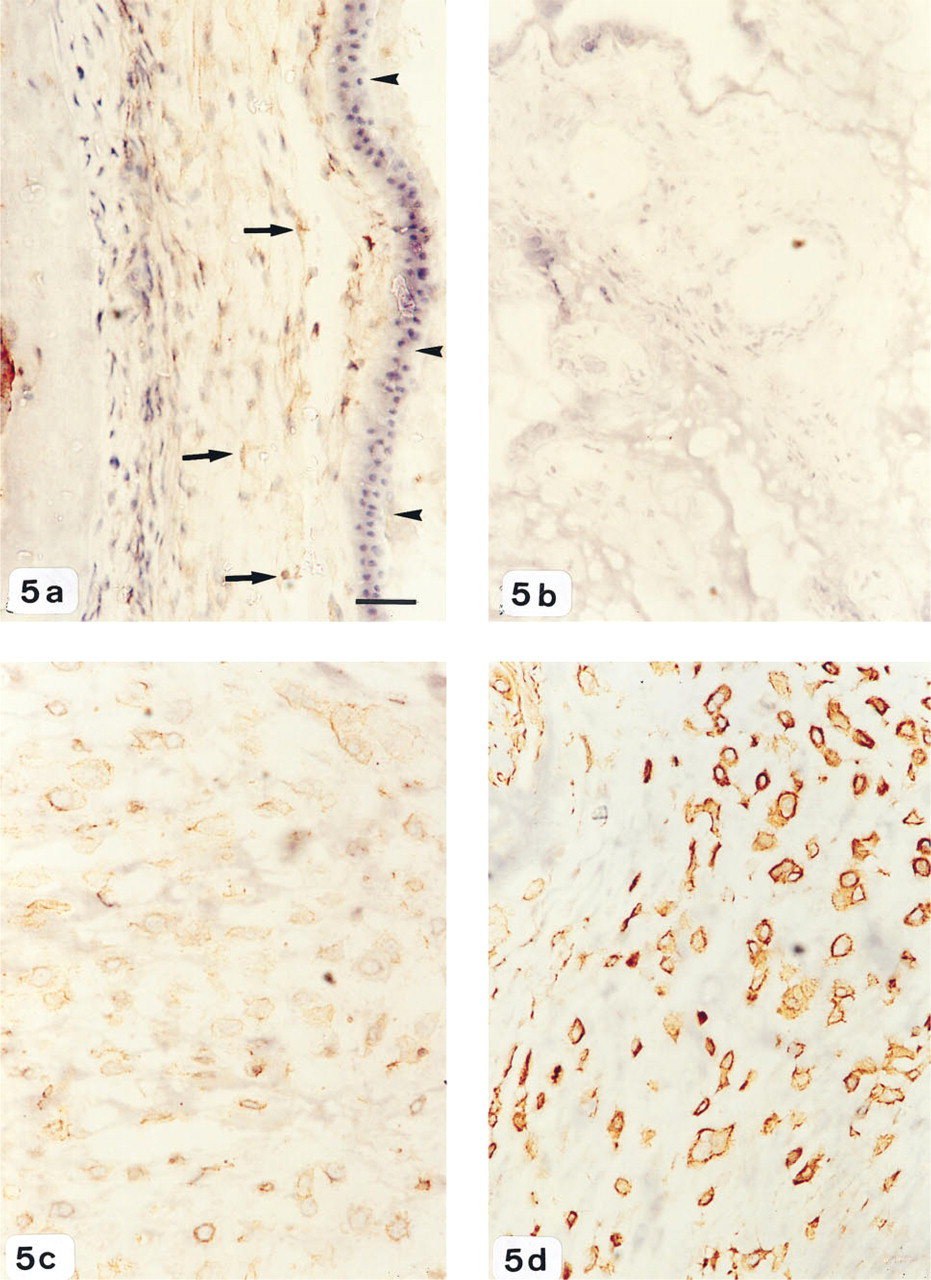
uPAR staining did not show any differences in intensity with labor. Cells of the chorion and amnion demonstrated very light uPAR immunoreactivity, whereas no detectable staining was observed in the placental villi (Figures 5a and 5b). Within the extravillous tissue, intense immunoreactivity for uPAR was present on the surface and cytoplasm of cells invading the decidual stroma (Figure 5c). uPAR-immunoreactive cells were identified as trophoblasts by their positive expression of cytokeratin (Figure 5d). No staining was observed when the primary antiserum was omitted or substituted with irrelevant or nonimmune immunoglobulins.
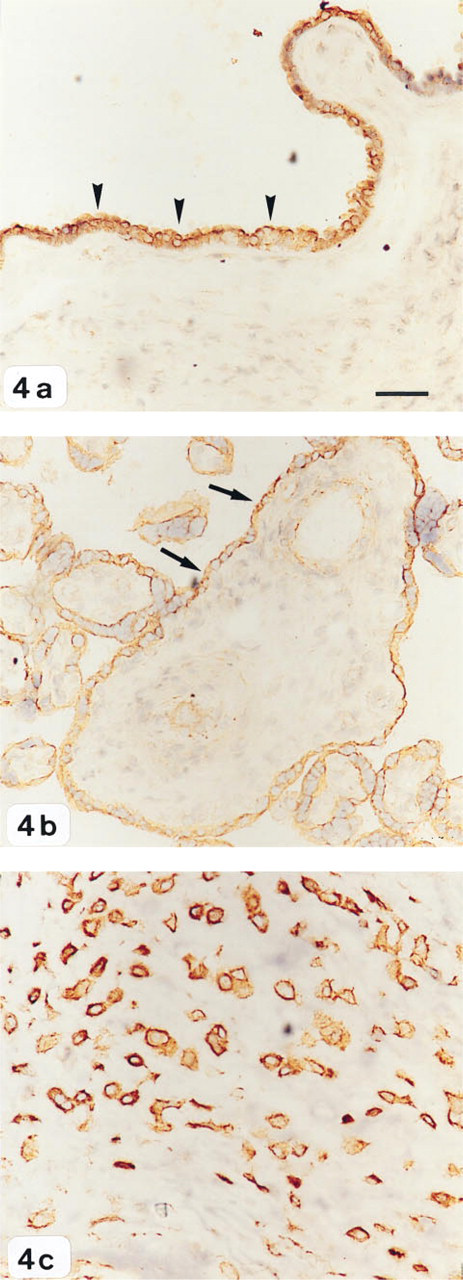
Identification of uPA protein in human gestational tissues by immunohistochemistry.
Western Blot Analyses
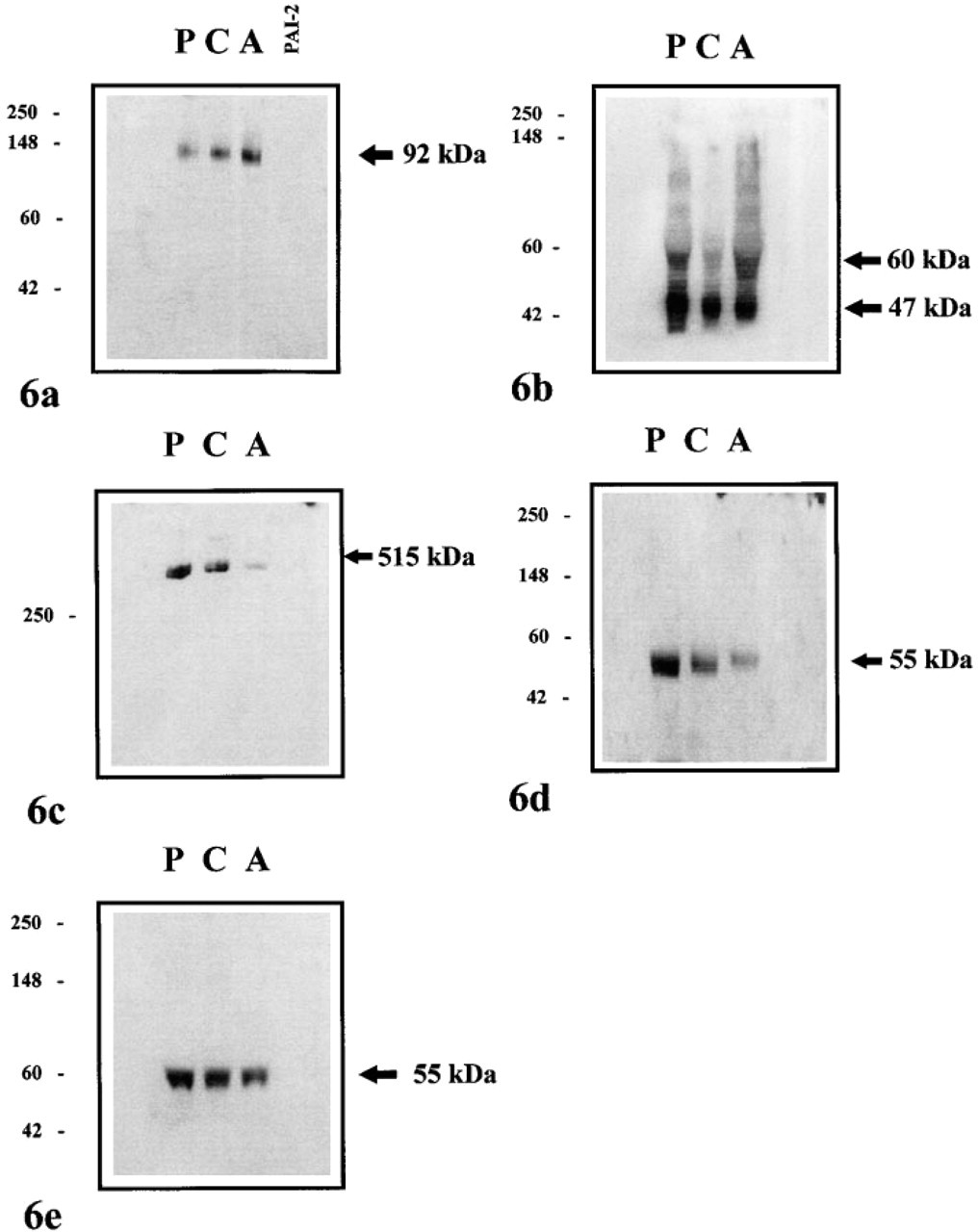
Human placenta, chorion, and amnion extracts were subjected to Western blotting as described in Materials and Methods. Using the PAIr monoclonal antibody at a concentration of 100 μg/ml, a single band at ∼92 kD, corresponding to the uPA-PAI-2 complex, was detected in all tissues (Figure 6a). No crossreactivity of PAIr antibody with purified PAI-2 standard was observed, confirming the specificity of this antibody for cleaved PAI-2.
Western blots probed with the PAI-2 polyclonal antibody revealed two prominent bands at 60 and 47 kD for all tissues examined (Figure 6b). The larger protein is the secreted, glycoslyated form of PAI-2, whereas the latter corresponds to the intracellular and nonglycosylated form of this inhibitor. The presence of higher molecular weight proteins was also detected, which most likely represent protease-inhibitor complexes.
Western blot analysis revealed that LRP was present predominantly in placental and chorion extracts, consistent with its cellular distribution reported in this study (Figure 6c). Very low levels of immunoreactive LRP were also found in amnion, but the expression of the receptor was always substantially lower than that detected in placenta and chorion. Immunoreactive uPAR and uPA were also detected in placenta, chorion, and amnion, with both proteins migrating at a molecular weight of 55 kD (Figures 6d and 6e).
Discussion
The aims of this study were to characterize the immunohistochemical localization of the relaxed conformation of PAI-2 (PAIr) in human gestational tissues obtained at term and to establish its co-expression with other components of the plasminogen activation cascade.
The data obtained in this study clearly establish the tissue-specific expression of PAIr immunoreactivity in human term gestational tissues. PAIr immunoreactivity was exclusively localized to amnion epithelium, with only minimal staining in the underlying chorion. PAIr immunoreactivity was not detectable in any of the trophoblastic tissues examined (villous and extravillous trophoblasts) by immunohistochemistry. In addition, the tissue-specific expression of PAIr immunoreactivity was not significantly altered in association with the onset of labor.
PAIr is formed during the inactivation of uPA, when an exposed reactive site loop of PAI is cleaved and inserted into the central β-sheet (Shore and Day 1995). The generation of PAIr at cell surfaces mediates binding to the multifunctional clearance receptor LRP. Once bound, the LRP-PAIr complex is very rapidly internalized to endosomes and subsequently degraded in lysosomes (Jensen et al. 1990). By this mechanism, inactivated uPA is removed from the cell surface and dissociated from uPAR. Unoccupied uPAR is recycled to the plasma membrane, thus sustaining focused proteolytic activity (Herz et al. 1992). It has previously been suggested that the rate of turnover of uPAR may represent a functional determinant of the capacity of a cell to express proteolytic activity on its surface and therefore its potential to degrade ECM directionally and invade surrounding tissues. Under conditions in which LRP function is deficient, uPA-PAI complexes may not be cleared from the cell surface, leading to diminished cell surface uPA activity and reduced cell invasiveness (Gafvels et al. 1993).
In this study, immunoreactive PAIr was identified in amnion epithelium, indicating that the rate of complex formation (i.e., uPA-PAI-2) exceeds the rate of internalization and degradation. Consistent with this proposal, term amnion epithelium, as shown by immunohistochemistry and Western blotting, expresses uPAR and stains intensely for immunoreactive uPA and PAI-2 but not for immunoreactive LRP. We propose, therefore, that the presence of immunoreactive PAIr is indicative of the in situ inactivation of uPA and the absence or a low rate of uPA-PAI-2 complex inter-nalization. Such expression of uPA cascade components would confer a capacity to degrade ECM and directionally localize proteolytic activity on the cell surface. On the basis of these data, although capable of ECM degradation, amnion epithelial cells should not display an invasive phenotype. Similarly, the presence of immunoreactive uPA, PAI-2, and LRP, but not of uPAR or PAIr, in term syncytiotrophoblasts suggests that this cell type expresses the relevant proteolytic activity to achieve breakdown of the ECM but lacks the capacity to assemble uPA activity on the cell surface. In addition, although this cell type has the potential to internalize inactive PAIr complexes via expression of LRP, phenotypically, term syncytiotrophoblasts, like amnion epithelial cells, should display no invasive potential.
In contrast, the positive staining of choriodecidual and extravillous tissue for both uPAR and LRP in trophoblast cells suggests a role for the expression of these proteins in focused cell surface localized ECM degradation and cell migration. In this regard, it is of interest that known ligands of uPAR and LRP, uPA and PAI-2, are co-expressed by this cell type. Consistent with the rapid turnover of receptor-bound, inactivated uPA by LRP, no cell surface staining for PAIr was detected in trophoblast cells. Our analysis of uPA cascade components in term choriodecidual and extravillous tissue provides evidence that trophoblast cells are able to localize uPA activity at the cell surface via uPAR, to degrade ECM proteins, and to rapidly internalize uPA-PAIr complexes by LRP. Consistent with this proposal, trophoblast cells retain their migratory properties by virtue of their ability to “capture” uPA activity to recycled cell surface receptors. Therefore the co-distribution of uPAR and LRP is in keeping with their role as receptors that function to modulate cell surface bound proteolytic activity and confer to a cell an invasive phenotype, as emphasized by the absence of PAIr.
In summary, the data obtained in this study (a) demonstrate that uPA cascade components are expressed by human term gestational tissues but are not significantly altered in association with labor onset, (b) for the first time provide information about the cellular and tissue localization of PAIr formation, and (c) are consistent with the hypothesis that PAIr immunoreactivity is not expressed by invasive cells that express LRP. Conversely, cells of noninvasive phenotype will only accumulate PAIr immunoreactivity in the absence of LRP.
Western blot analyses of human placenta (P), chorion (C), and amnion (A) extracts (50 μg protein/ lane) for
In conclusion, these results indicate an association between known capacity for directional ECM degradation (i.e., the ability to invade) and the expression of immunoreactive components of the uPA cascade. On the basis of this association, we suggest that expression of both LRP and uPAR is necessary for the expression of an invasive phenotype by human gestational tissues. Confirmation of this proposal requires further experimentation to establish the rates of uPAR-uPA–PAIr turnover by invasive cell types.
Immunohistochemical analysis of uPAR expression in human gestational tissues.
Footnotes
Acknowledgements
Supported by Grants from the 3AW Community Services Trust, the R.E. Ross Trust, and the National Health and Medical Research Council of Australia. GER is in receipt of a National Health and Medical Research Council of Australia Senior Research Fellowship.
We gratefully acknowledge the assistance of clinical research midwife Sr Debra Rushford, and the midwifery and obstetric staff of the Royal Women's Hospital, Carlton, Victoria, Australia, for collection of human gestational tissues.