
Abstract
Zinc is an essential element in mammalian development. However, little is known about concentrations of zinc in specific regions/organs in the embryo. We have employed selenite autometallography (AMG) and TSQ histofluoroscence to detect histochemically reactive (chelatable) zinc in whole midsagittal embryos and sections from neonatal mice. Chelatable zinc exhibited a broad distribution, being particularly localized to rapidly proliferating tissues, such as skin and gastrointestinal epithelium. Zinc was also observed in various types of tissues such as bone and liver. In the perinatal central nervous system, zinc was present almost exclusively in choroid plexus. The two methods used demonstrated generally similar distributions with some exceptions, e.g., in liver and blood. The ubiquity of zinc in the embryo, particularly in rapidly proliferating tissues, suggests a widespread role in fetal physiology.
Z
Materials and Methods
Specimen Collection
Pregnant CD-1 female mice (conception confirmed by vaginal plug after mating; designated E0.5) at 13.5, 15.5, and 17.5 days after conception (E13.5, 15.5, and 17.5, respectively) were anesthetized with sodium pentobarbital and the embryos quickly dissected out in cold PBS, pH 7.4 and frozen on dry ice. Mouse pups on the day of birth, designated P0 (postnatal day 0) were sacrificed and processed as above. Whole specimens were then sectioned (20-μm thick) on a cryostat (−20C), thaw-mounted on glass slides, dried overnight, and stored at −70C until needed.
Selenite Autometallography
For the selenite method, pregnant females and newborn pups were injected IP with sodium selenite (20 mg/kg) and allowed to survive for 1 hr. Specimen collection proceeded as described above. Slides were then immersed in a developer solution as described previously (Danscher 1982). Sections were counterstained with toluidine blue, cressyl violet, and hematoxylin and eosin (H&E) alternatively.
TSQ Histofluorescence
Histofluorescence with TSQ was performed as described previously (Frederickson et al. 1987). Tissue images were captured into a PC workstation with a digital camera (SPOT RT; Diagnostic Instruments, Sterling Heights, MI) using a UV-2A filter block (330-380 nm; barrier 420 nm). Tissue autofluorescence was assessed and eliminated using sections treated with a solution not containing TSQ. Previous treatment with sodium selenite resulted in elimination of TSQ fluorescence.
Results
Zinc has been demonstrated histochemically by a variety of approaches, beginning with the description of dithizone, a zinc-specific chelator, by McNary (1954). This technique was largely supplanted by Timm's histochemistry or autometallography (AMG) for characterization of chelatable metals (Danscher et al. 1973; Haug 1975). Most recent zinc studies have employed variations of the AMG method (e.g., Sorensen et al. 1998; Kristiansen et al. 2001) or one of a number of zinc-specific fluorescent dyes (for review see Kimura and Aoki 2001). Because of known differences in the staining patterns produced by the two approaches and various other factors (see Discussion), we employed both AMG and TSQ histofluorescence to characterize the distribution of tissue chelatable zinc in the mouse fetus and on the day of birth.
Chelatable zinc was concentrated in a variety of tissues and exhibited distinct patterns of distribution in midsagittal sections at E13.5, 15.5, 17.5, and P0. At E13.5, chelatable zinc was barely detectable by selenite (not shown). Although also very low when assessed with TSQ, some signal was observed in the digestive tract epithelium, epidermis, and cartilage (Figures 1–3). At E15.5, TSQ histofluorescence was also observed in bone (Figure 3). Selenite staining generally overlapped that of TSQ at E15.5, although some labeling of hepatocytes and blood vessels was observed (data not shown). Again, at E17.5 and P0, when most organs are fully developed, the two methods demonstrated similar patterns of zinc distribution, being most prominent in the alimentary tract and skin. The liver parenchyma at this stage was heavily labeled with the selenite method but not by TSQ. Other tissues exhibiting more moderate concentrations of zinc were pancreas, blood, cartilage, choroid plexus, and vertebrae. Tissues that displayed very little or no zinc were brain and spinal cord, striated muscle, lung, thymus, and thyroid gland. Under high-magnification light microscopic analysis, zinc appeared primarily in cell cytoplasm. The reader is referred to Figure 4 and Table 1 for an overall description of the findings at E17.5.
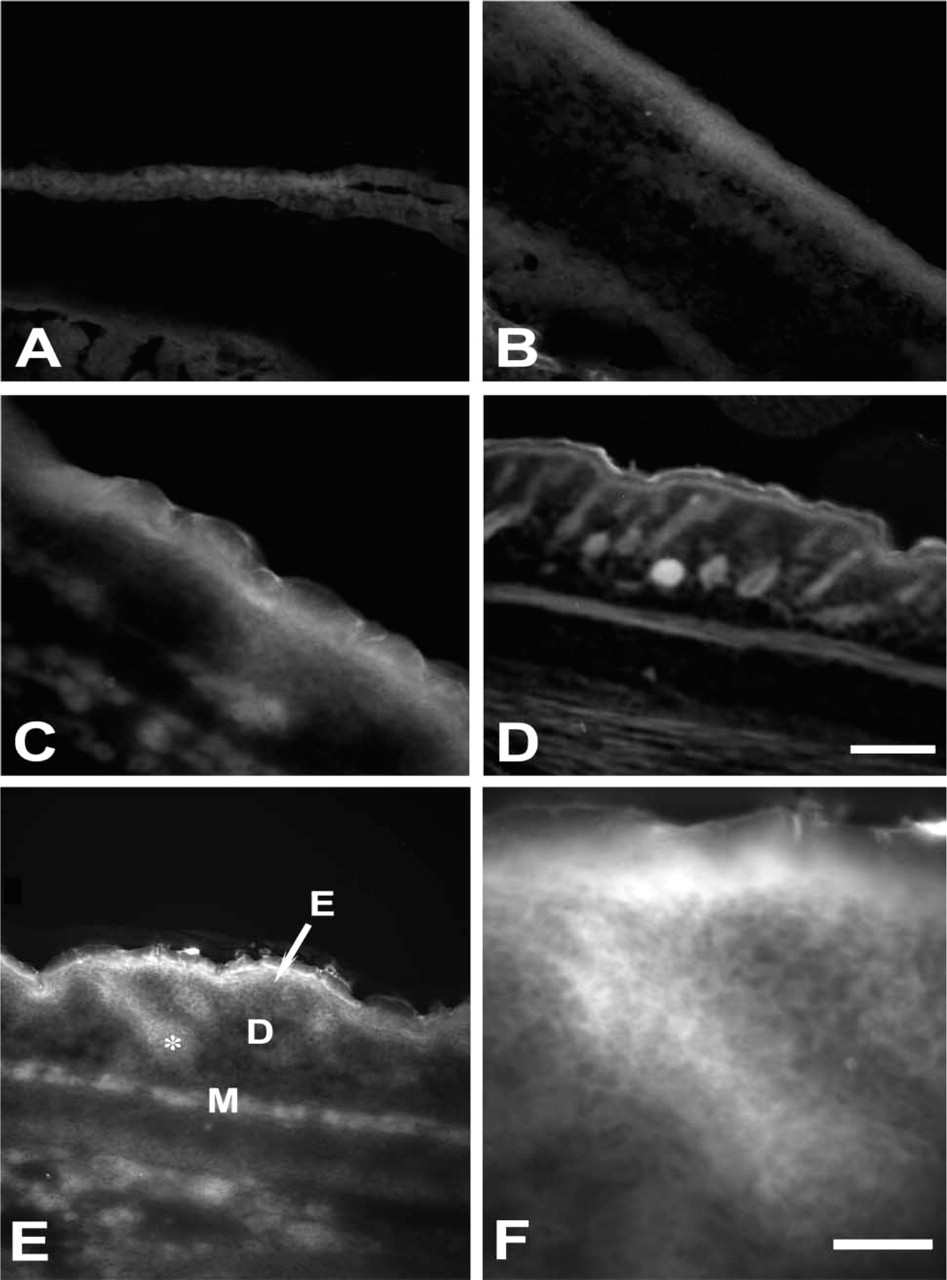
TSQ histofluorescence in skin at E13.5 (
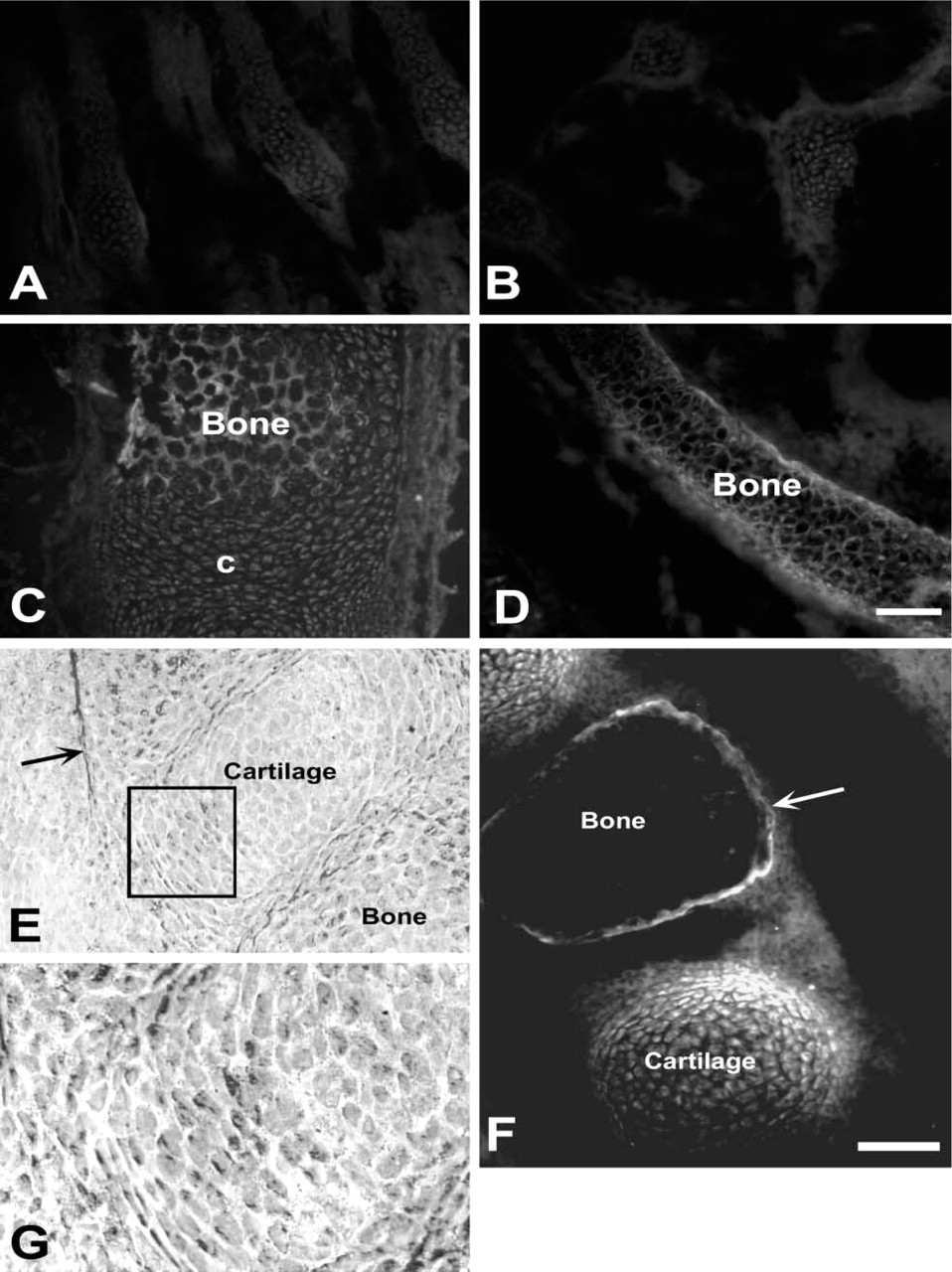
Zinc histofluorescence in cartilage and bone at E13.5 (
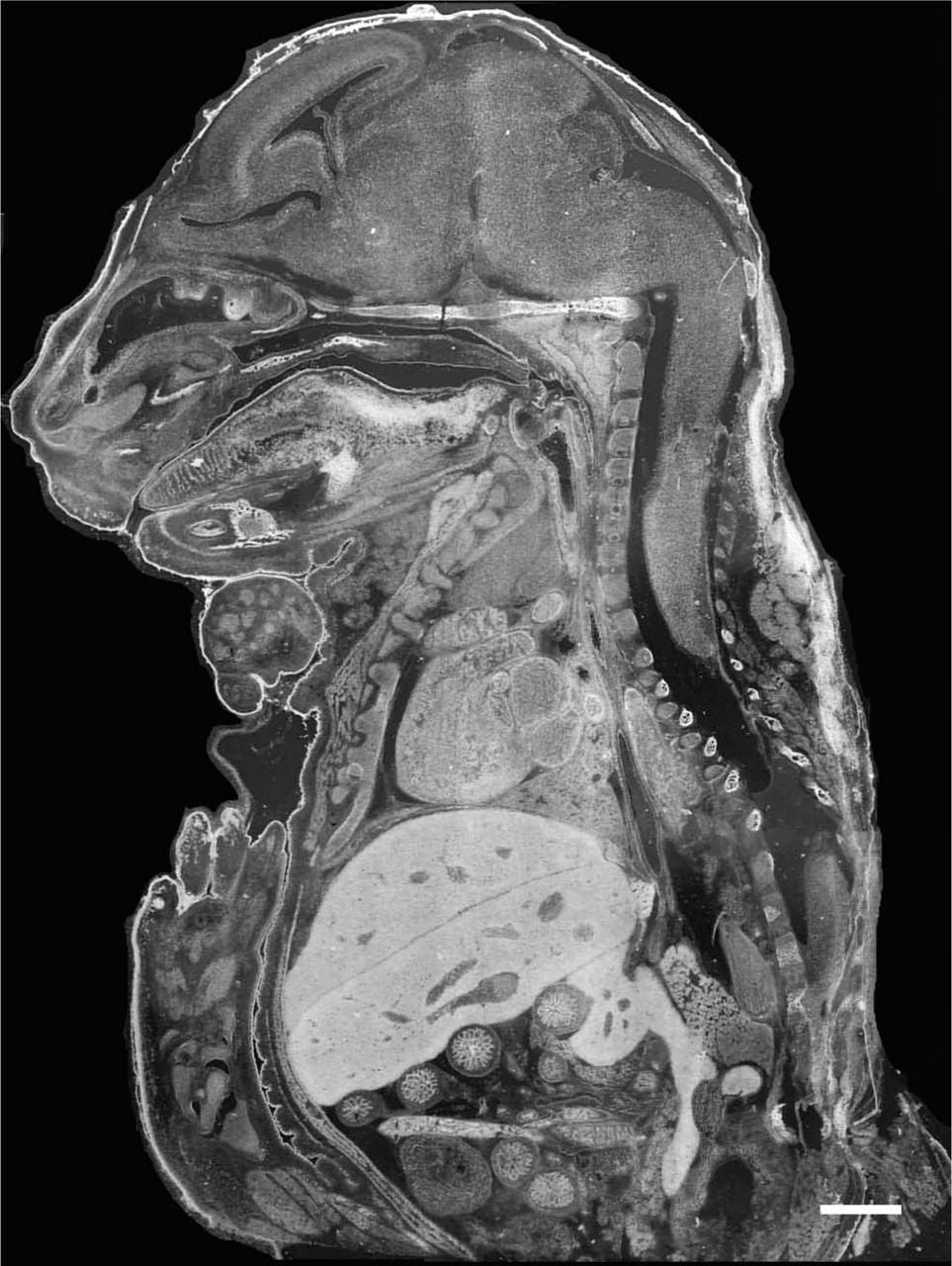
Frozen midsagittal section (12 μm thick) at 17.5 days after conception, stained with the selenite AMG method and photographed under darkfield illumination. Note the high concentration of zinc in liver, skin, and digestive tract among other tissues. For reference see Figure 2. Bar = 500 μm.
Relative intensity of zinc labeling in various tissues at E17.5 with respect to published measurements of total zinc
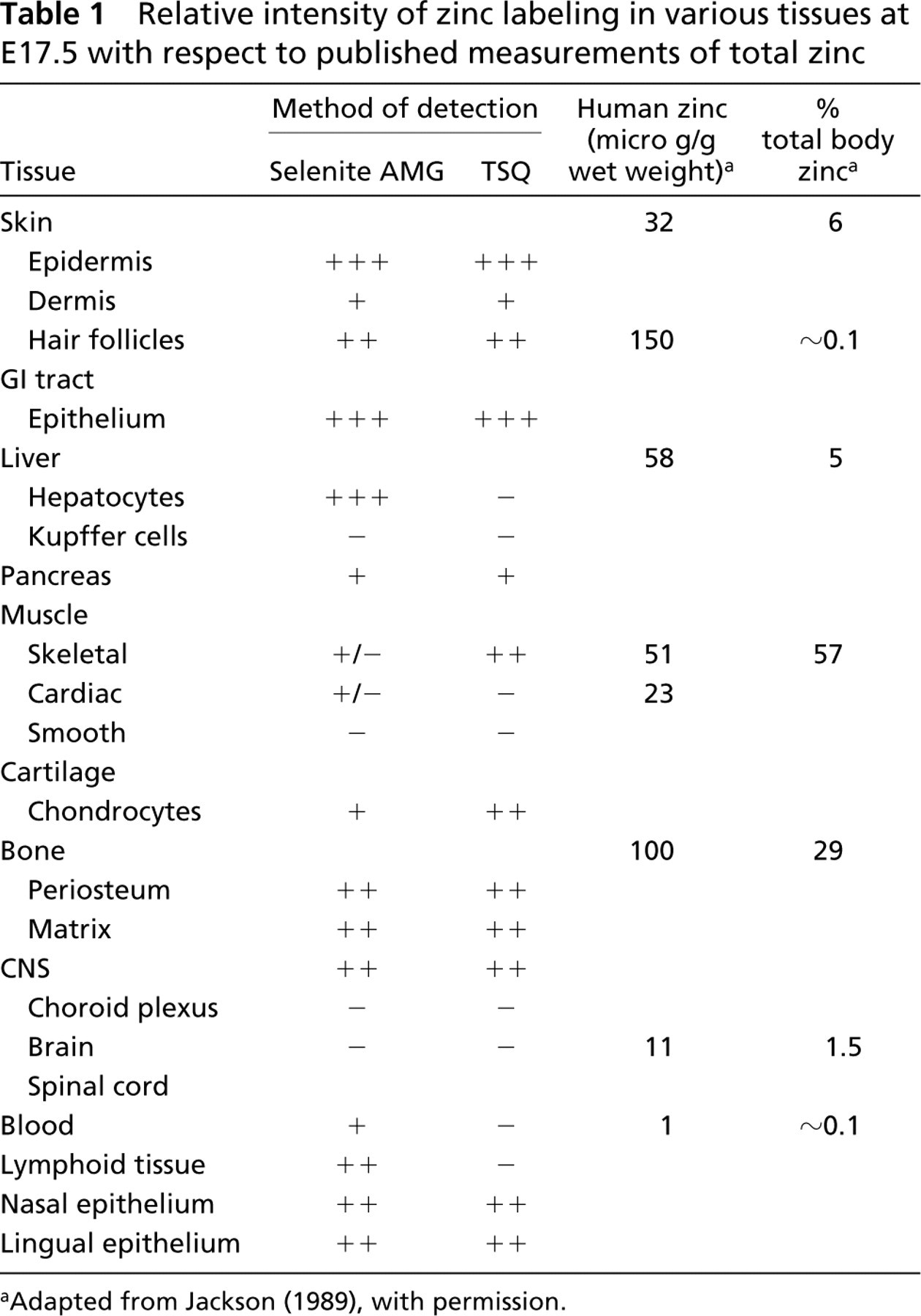
Adapted from Jackson (1989), with permission.
Alimentary Tract
Chelatable zinc was observed throughout the digestive tract beginning at E13.5 and continuing through E15.5 (not shown). At these ages, tissue concentrations of zinc were low and were detected mainly by TSQ. Labeling was typically restricted to the luminal aspect of the intestinal loops. At E17.5, TSQ and selenite showed an overlapping distribution (Figure 2) and were most heavily concentrated in the columnar epithelium lining the tract, beginning at the lower esophagus. In these epithelial cells, AMG zinc particles were distributed throughout the cell, although they did not typically reach the brush border (Figures 2A and 2B). Zinc was also evident, although to a much lesser extent, in the peritoneum enveloping most of the digestive tract, in the abdominal wall, and in mesenteric adipocytes. Zinc demonstrated by TSQ exhibited a similar pattern (Figures 2C and 2D). At birth, both methods were overlapping, resembling that at E17.5 (not shown)
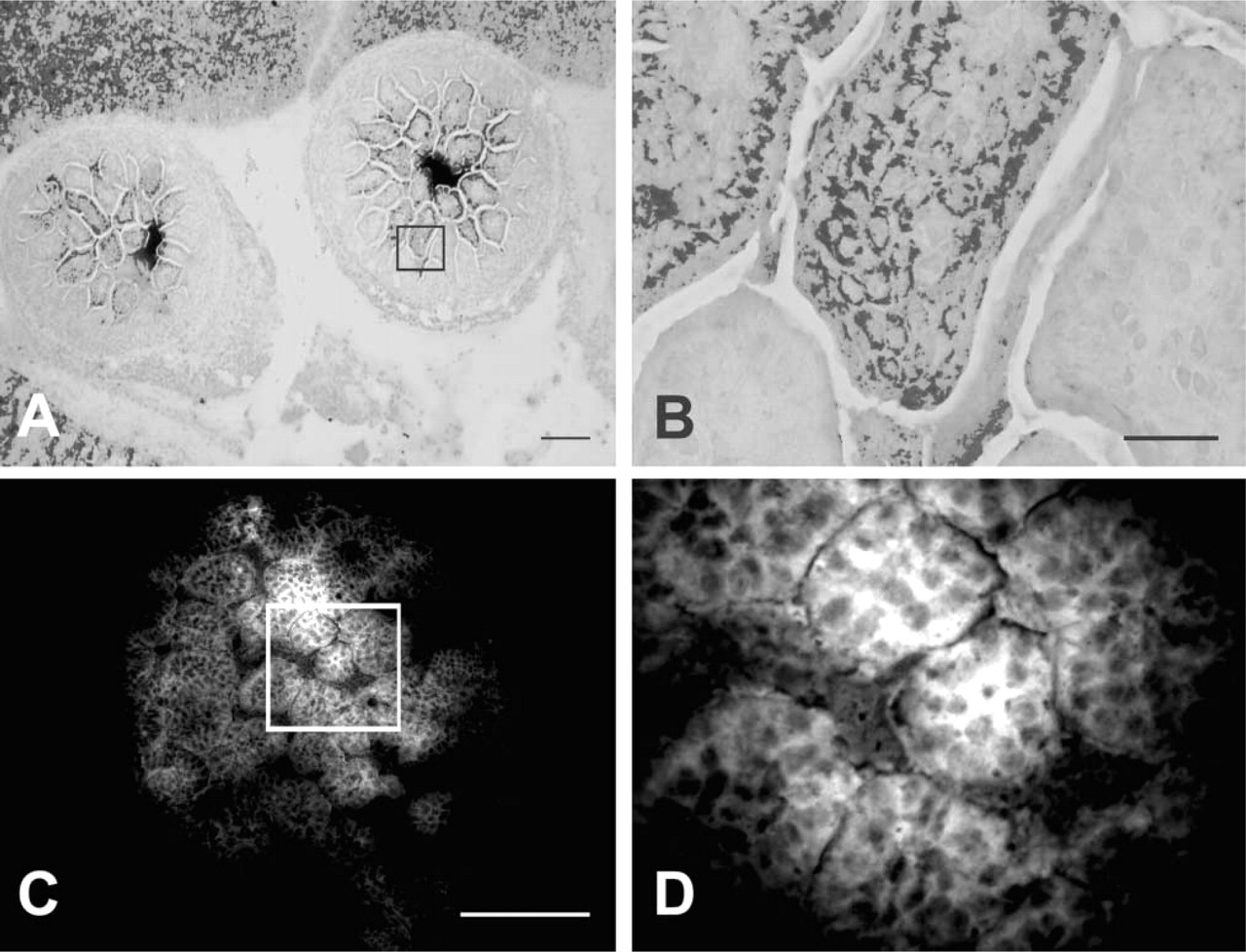
Distribution of selenite AMG-demonstrated chelatable zinc in the alimentary tract (
Liver
In the liver, selenite AMG labeling was primarily restricted to hepatocytes, beginning at E15.5 (data not shown). No specific organizational pattern of zinc-positive hepatocytes could be discerned. In addition, zinc appeared in biliary canaliculi (Figure 5A). Hepatic elements devoid of chelatable zinc were also observed, including hematopoietic cells, e.g., megakaryocytes (Figure 5B), and connective tissue (i.e., Glison's capsule) enveloping the liver. The TSQ method, in contrast, did not detect chelatable zinc in the liver.
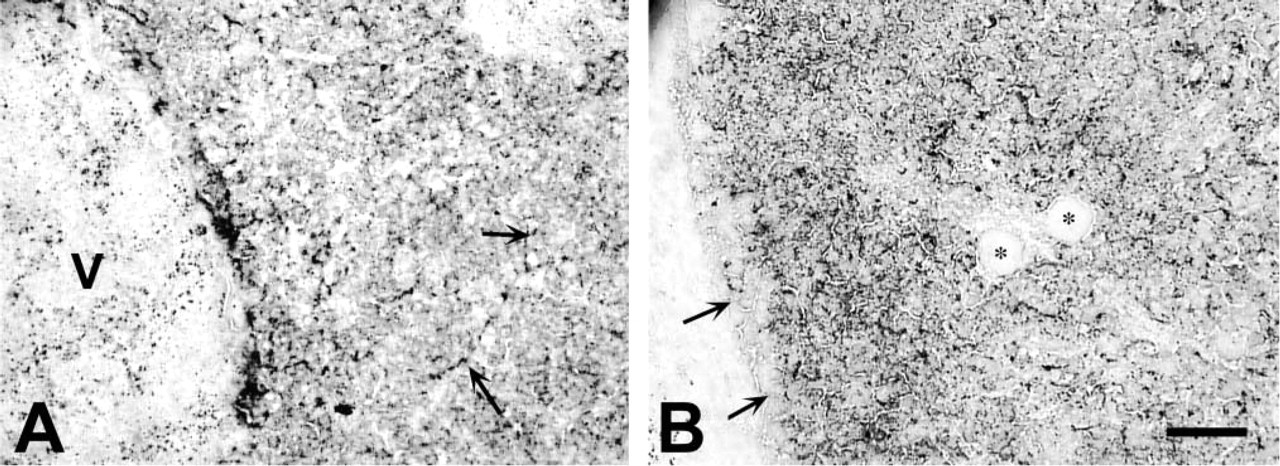
Liver stained with selenite AMG and H&E at E17.5. Note the abundant reaction throughout the tissue. (
Vasculature
Similar to the liver, blood vessels exhibited a method-specific differential zinc distribution. TSQ did not label zinc over blood vessel lumen profiles (data not shown) at any ages observed, whereas the AMG method demonstrated zinc over all blood vessels examined beginning at E15.5 and continuing through birth (Figure 5A). Silver particles did not appear to be localized to blood or endothelial cells but instead were situated over the lumen, apparently related to the plasma.
Skin
In skin, both AMG and TSQ protocols demonstrated the highest zinc concentrations in the epidermis, particularly in the stratum granulosum (Figures 1E and 1F). The AMG method, however, did not label skin until E17.5, whereas TSQ demonstrated some labeling at earlier ages (Figure 4). Hair follicles of the mystacial whiskers exhibited chelatable zinc with both methods, beginning at E17.5 (not shown).
Pancreas
The pancreas was moderately positive for zinc with both methods, beginning at E17.5. Both AMG reaction product and TSQ fluorescence were most concentrated in pancreatic islets, where labeling was distributed throughout the islet. Moreover, zinc was present in only a small percentage (i.e., <10%) of islet cells (data not shown). Virtually no labeling was demonstrated over the exocrine pancreas.
Bone and Cartilage
Up to and including E15.5, AMG reaction product was absent in bone, whereas TSQ demonstrated light labeling of an occasional vertebra and cartilage (Figure 3). At E17.5, both AMG and TSQ methods labeled bone, especially vertebrae, as well as skull and the long bones of the limbs (Figure 3). However, this labeling was inconsistent with many instances of even a single bone exhibiting both moderate and low levels of zinc. When present, zinc was most concentrated in periosteum and the bony matrix. In cartilage, zinc was most evident in intervertebral discs, mainly in chondrocytes at the periphery of the disc (Figures 3F and 3G). At P0 almost all bones contained zinc, although the bone marrow was noticeably unlabeled (see above discussion of megakaryocytes in liver). Cartilage on the day of birth exhibited little if any labeling with either selenite AMG or TSQ (not shown).
Central Nervous System
Chelatable zinc maintains a well-known distribution in the postnatal rodent CNS, particularly in the fore-brain and hippocampus (Czupryn and Skangiel-Kramska 1997; Slomianka and Geneser 1997; Nitzan et al. 2002; Valente et al. 2002). In contrast, neither AMG nor TSQ demonstrated chelatable zinc in the brain during gestation, with the single exception of ependymal and endothelial cells of the choroid plexus (CP) (Figure 6). Labeling for zinc in CP was first observed at E15.5. Demonstration of zinc in the CP was more robust at E17.5, with no apparent changes on P0. Chelatable zinc was demonstrated, albeit in low abundance, at E17, and was somewhat more concentrated at P0. At birth, labeling for zinc was present at all levels of the cord and hindbrain, particularly in the dorsal part (Figure 7).
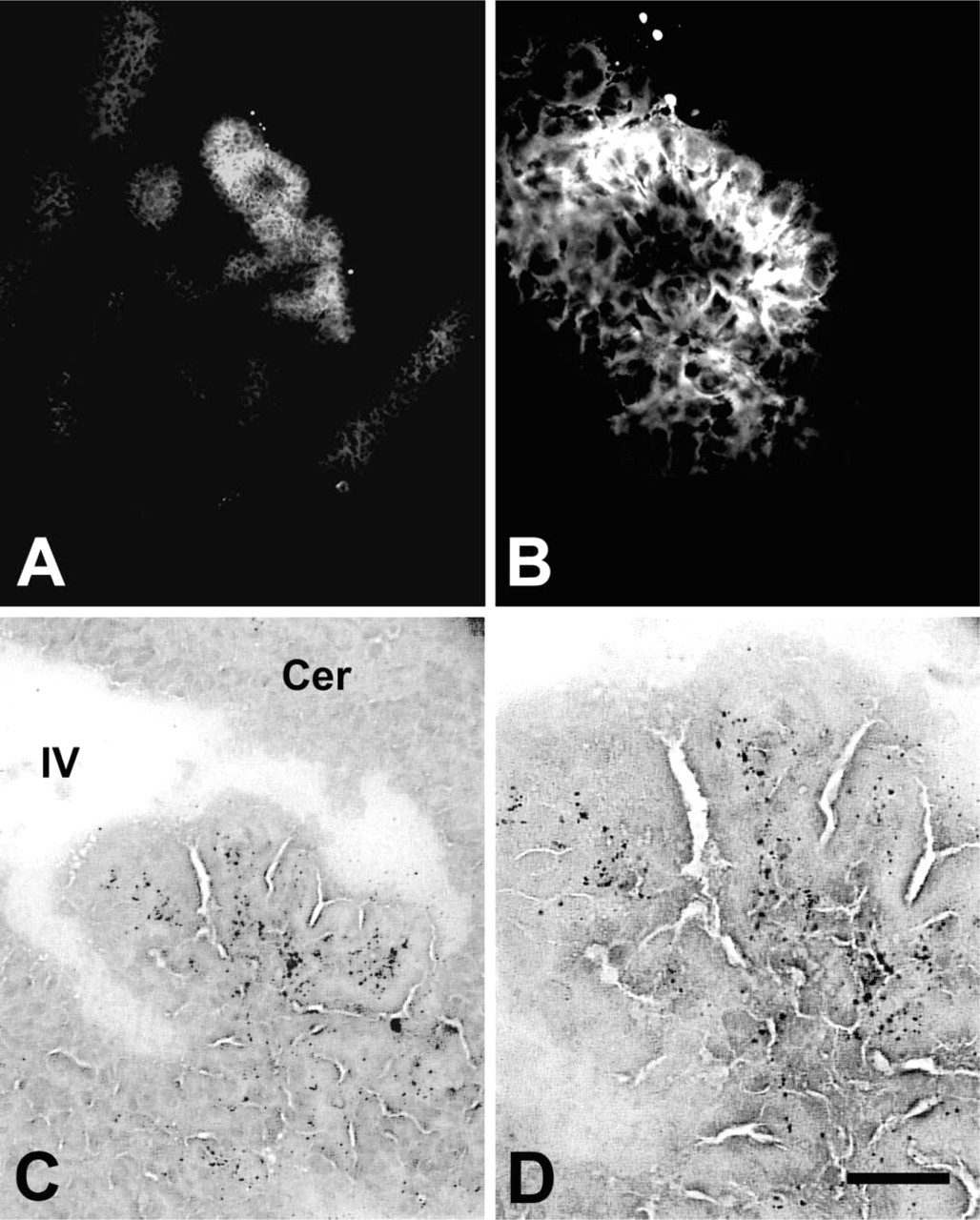
Chelatable zinc in choroid plexus of the fourth ventricle (IV) demonstrated by TSQ histofluorescence (
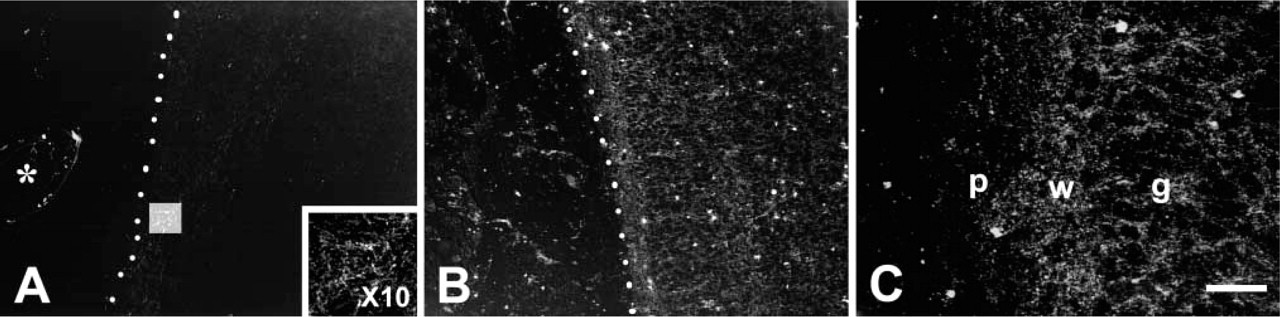
Darkfield images of spinal cord (hind lumbar) stained with selenite AMG (without counterstain) at E17.5 (
Other Areas
Another strongly zincergic region observed with selenite AMG and TSQ was the tongue epithelium (not shown). Parapharyngeal lymph tissue was moderately labeled with the selenite method, although not with TSQ (not shown). Testis exhibited low to moderate labeling with both methods (data not shown), as did bladder, whereas cardiac muscle was more intensely labeled by TSQ (data not shown).
Discussion
Methodological Aspects
In the present study we have used two different zinc-tracing methods to assess the distribution of zinc in the mid- to late-gestation mouse embryo and on P0. Overall, the two methods, sodium selenite AMG and TSQ histofluorescence, demonstrated a similar distribution of zinc in the developing mouse. A number of exceptions are noted above. For example, the liver was heavily labeled with the AMG method in the late prenatal and neonatal mouse but exhibited virtually no fluorescence with the TSQ method. This type of difference might be attributed to the fact that selenite AMG traces zinc indirectly, i.e., it is based on the complexing of zinc ions with an anion, e.g., selenite (Danscher 1982), and subsequent silver enhancement. In contrast, TSQ is a specific zinc-sensitive fluorophore (Frederickson et al. 1987). Furthermore, selenite and TSQ are inherently different types of molecules, the former being ionic and the latter hydrophobic. This difference may affect labeling of zinc in highly aqueous vs lipophilic environments (Marin et al. 2000).
It is noteworthy that the distribution of AMG reaction we observed in the liver is reminiscent of the staining patterns reported there previously for metal-lothioneins (MTs) I/II (Nishimura et al. 1989; Penkowa et al. 1999), including the strikingly “blank” megakayocytes shown in Figure 7. It is possible that the AMG reaction noted in the liver is due to zinc that is dynamically competed for by selenite from the pool of protein-bound zinc (e.g., MT-bound).
To address the issue of a possible false-positive reaction with selenite AMG, we employed a negative control, i.e., application of developer to sections derived from animals not previously exposed to selenite. Gold, silver, and mercury are the three metal species most likely to produce a spurious reaction (Danscher 1996) and will do so with or without previous treatment of tissue with sulfide or selenite. Kristiansen et al. (2001) note that this approach is the best-controlled method to ascertain specificity and, in fact, the specificity of the selenite method has been assessed previously in brain (Danscher 1982). In the present work, control animals did not exhibit autometallographic deposits.
A major consideration in employing both protocols is the dependence of the selenite method on various biological factors (e.g., Slomianka et al. 1990; Valente et al. 2002). In addition, gestation is a complex process that no doubt contributes additional factors that can affect selenite pharmacokinetics (Danielsson et al. 1990). It was therefore necessary to confirm the selenite findings by another method, preferably a direct method, e.g., TSQ. Finally, differences, albeit minor, that were obtained using the TSQ and AMG methods have been noted previously (Frederickson et al. 1992). In general, AMG permits mapping of the metal with reference to surrounding histology although, as mentioned above, it is subject to a variety of physiological influences such as transport mechanisms and tissue permeability (for review see Frederickson et al. 2000).
Although the present work assessed only chelatable zinc, our findings largely correlate with published concentrations of total tissue zinc (Table 1). This suggests that “free” or chelatable zinc, which exists as a fraction of total zinc, is maintained in balance with protein-bound intracellular zinc, possibly being stored and released as needed. Recent studies using fluorescent resonance energy transfer (FRET) have provided important support for this idea, demonstrating, for example, that nitric oxide induces release of zinc bound to metallothionein (Pearce et al. 2000).
Functional Aspects of Chelatable Zinc
The detection of zinc in bile canniliculi and throughout the digestive tract, including the intestinal lumen and the liver, illustrates a possible zinc enterohepatic circulation in utero. The interplay between secretion and absorption of zinc throughout the gastrointestinal (GI) tract has been described previously (Methfessel and Spencer 1973; Krebs 2000). It appears that zinc is essential for normal digestive tract physiology, although its precise contribution is not well understood (Semrad 1999). Our demonstration here of significant quantities of heterogeneously distributed zinc in the GI tract is consistent with the idea that zinc, and more specifically chelatable zinc, is involved in development and/or function of the fetal GI tract as well as in postnatal and adult animals.
Another area in which zinc distribution can readily be related to function is bone, where it is known to be stored (Jackson 1989; King et al. 2000). Zinc participates in bone formation and growth (Ma et al. 2001; Ma and Yamaguchi 2001; Ovesen et al. 2001). Zinc deficiency leads to a variety of bone pathologies, among which are impaired fracture healing and osteoporosis (Lowe et al. 2002). The localization of free zinc in bone suggests that it is secreted by local cells to exert effects in a paracrine, autocrine, and possibly an endocrine manner.
For decades zinc has been regarded as an essential element for the normal physiology of skin (Perafan-Riveros et al. 2002), possibly via its role as an antioxidant (Rostan et al. 2002). The present study shows that free zinc is abundant in skin, which is consistent with the thesis that a readily releasable pool of zinc is involved in skin development and function. Hershfinkel et al. (2001) have recently described an extracellular zinc-sensing receptor on skin cells, which mediates the intracellular release of calcium and, in turn, cell proliferation in a model of wound healing (Hershfinkel and Sekler, unpublished observations). The presence of significant quantities of chelatable zinc in epidermis may therefore suggest that this layer serves as a “trip wire” for initiating cell mechanisms in the dermis to respond to skin injury.
The CNS is one area of the developing mouse in which chelatable zinc, with the exception of the choroid plexus and spinal cord, is conspicuously absent. Areas of the mouse brain that contain high concentrations of free zinc postnatally, e.g., the hippocampus and amygdala, exhibit virtually no chelatable zinc before birth. It is interesting in this regard that, postnatally, the cerebellum contains among the highest concentrations of (total) zinc in the brain (Takeda et al. 2001), although little of it is the chelatable variety. In fact, the lack of chelatable zinc in the fetal brain does not indicate the absence of zinc, because many enzymes and transcription factors containing zinc are present even in early stages of mammalian development. It is therefore, interesting to speculate about the nature of the change that occurs during the days after birth leading to the establishment of a pool of free zinc from existing stores. This comes to be concentrated principally in synaptic vesicles in forebrain excitatory neurons (Frederickson et al. 2000).
Our study shows that, with a few outstanding exceptions such as brain, “free” or loosely bound zinc is present and is heterogeneously distributed throughout the fetal mouse. Furthermore, we have observed that chelatable zinc is particularly abundant in those organs known to possess high concentrations of (total) zinc. This suggests, first of all, a well-regulated system of zinc homeostasis, even at this early age. Second, it suggests a role for zinc in a wide variety of biological processes, especially because this zinc is present in its most readily usable form. Further research is needed to elucidate the specific functions subserved zinc in the various organs in which it is concentrated.
Footnotes
Acknowledgements
Supported by BGNegev (#3341) to IS and WFS. We wish to thank Dr Gershon Perach for assistance in data analysis.