
Abstract
The arterial endothelial cells of the rete capillaries of the eel were examined by transmission electron microscopy on thin sections, on freeze-fracture replicas, by scanning electron microscopy, after cytochemical osmium impregnation and perfusion with peroxidase. The study revealed the existence of membrane-bound tubules and vesicles that open at both the luminal and abluminal poles of the cell and at the level of the intercellular space. The tubules are straight or present successive dilations and constrictions. They branch in various directions and intrude deeply into the cell cytoplasm, forming a complex tubular network within the cell. Immunocytochemical techniques were applied on immersion-fixed tissues and on perfusion of the capillaries with albumin and insulin. These demonstrated that the tubular–vesicular system is involved in the transport of circulating proteins. Furthermore, protein A–gold immunocytochemistry has revealed the association of actin with the membranes of this system. On the basis of these results, we suggest that the transendothelial transport of serum proteins takes place by a transcytotic process through a membrane-bound tubular–vesicular system and is equivalent to the large pore system presumed from functional studies.
V
The rete mirabile on the eel swimbladder wall constitutes a unique model for the study of the morpho-functional properties of blood capillaries (Rasio et al. 1993). Indeed, the rete mirabile with its vascular connections allows well-controlled perfusion experiments and precise measurements of capillary permeability (Rasio et al. 1977), assessments of capillary metabolic requirements (Rasio et al. 1993), and morphological studies of the vascular wall (Bendayan et al. 1975) with the ultrastructural visualization of transendothelial transport pathways for tracer proteins (Bendayan 1980; Bendayan and Rasio 1996a). Recent physiological and immunocytochemical studies performed with this blood capillary preparation have addressed the permeability characteristics and transendothelial transport of albumin and insulin (Bendayan and Rasio 1996a), demonstrating that transcytosis of insulin occurs preferentially to albumin through a particular population of plasmalemmal vesicles or tubules, probably governed by receptor-mediated mechanisms.
Although the morphological characteristics of the capillary tissue isolated from the eel swimbladder rete mirabile have already been reported (Dorn 1961; Bendayan et al. 1974,1975; Stray–Pedersen and Nicolaysen 1975; Wagner et al. 1987), in the present study we have investigated the fine structural features of the endothelial cells composing the arterial capillary wall of the rete mirabile. A combination of various techniques has demonstrated the existence of an intricate tubular–vesicular system that intrudes deeply the endothelial cells and opens at both the luminal and abluminal fronts of the cells as well as in the intercellular junctional space. This system is involved in transendothelial transport of circulating molecules because serum proteins were found within its luminal space. In addition, cytoskeletal proteins have been detected in close association with the membranes of the tubules. This tubular system may therefore be involved in the active transendothelial transport of serum proteins and may correspond to the large pore system that has been proposed to account for vascular permeability.
Materials and Methods
The morphological characteristics of the rete capillaries and their involvement in transcapillary transport were investigated in electron microscopy by applying various morphological and cytochemical techniques.
Tissue Preparation
Large eels of the genus Anguilla anguilla, weighing 1–1.5 kg, were caught in the St. Lawrence river and kept in running tapwater. After anesthesia with tricainemethane sulfonate, MS222 (Sandoz Pharmaceutical; East Hanover, NJ) added to the water at the concentration of 0.3 g/liter and abdominal incision, the two retia on the swimbladder wall were exposed and fixed by perfusion through the preretal artery with 1% glutaraldehyde in 0.1 M phosphate buffer with or without 2% acrolein and 5% DMSO (Bendayan et al. 1974,1975; Sandborn et al. 1975). The perfusion fixation was carried out for at least 10 min. The tissue was removed from the swimbladder and maintained in the same fixative for an additional hour. In some occasions, and to retain the endogenous serum proteins in situ, the capillary tissue was fixed by immersion with a 1% glutaraldehyde solution.
For transmission electron microscopy, the tissues were postfixed with 1% osmium tetroxide and embedded in Epon according to standard techniques. In a different set of experiments, osmium impregnation was carried out after glutaraldehyde fixation according to the techniques of Friend (Friend and Murray 1965), modified as follows. After 1-hr fixation with 1% buffered glutaraldehyde, the tissue samples were immersed for 48 hr in unbuffered 2% osmium tetroxide at 37C and then treated for 2 hr with 2% uranyl acetate at pH 6.0 before dehydration and embedding in Epon.
In a different series of experiments, the capillary tissue was fixed by perfusion with 1% glutaraldehyde for 15 min. After perfused washing with phosphate buffer, the capillaries were perfused with a 1% solution of horseradish peroxidase (Sigma Chemical; St Louis, MO). This was carried out for 30 min, after which the tissue was immersion-fixed for 2 hr in 1% glutaraldehyde. After rinsing, the capillary tissue was embedded in gelatin and thick sections were obtained with a Smith–Farquhar tissue chopper. These sections were then reacted with DAB (Graham and Karnovsky 1966; Simionescu et al. 1975) and processed for electron microscopy.
For scanning electron microscopy, thick segments of the tissue were sectioned using a Smith–Farquhar tissue chopper and digested with 2% diastase in Ringer's lactate solution for 15 min. After critical point-drying, the segments were mounted on supports and coated with gold before examination.
For the freeze-etching technique, platinum-carbon replicas were generated according to standard techniques after fixation of the tissue with buffered 1% glutaraldehyde.
For the immunocytochemical investigation of vascular permeability, the rete was perfused through the preretal artery with a Krebs-Ringers' bicarbonate buffer supplemented with glucose (5 mM) and bovine serum albumin (4 g/100 ml) and equilibrated with a gas mixture of 95% O2–5% CO2 (Rasio et al. 1977). Insulin was added to the perfusion medium to a final concentration of 0.5 g/100 ml. The perfusion was carried out for 30 min at room temperature (RT) under constant flow (0.6 ml/min) and pressure (45 cm H2O). At the end of perfusion, the tissues were fixed by immersion with 1% glutaraldehyde in 0.1 M phosphate buffer, post-fixed with 1% osmium tetroxide, and embedded in Epon. Thin sections were mounted on nickel grids and processed for protein A–gold immunocytochemistry (Bendayan 1995) for the detection of bovine serum albumin and insulin using specific antibodies (Bendayan and Rasio 1996a). The anti-BSA (Sigma) was a rabbit antibody and the anti-insulin (Miles Biochemicals; Etobicoke, Ontario, Canada) was a guinea pig antibody, both diluted at 1:200. For labeling, thin tissue sections mounted on grids were first treated with a saturated solution of sodium metaperiodate for 30 min, rinsed, and then incubated with one of the specific antibodies for 2 hr at RT. After a rinse of 0.01 M PBS, the tissue sections were incubated with the protein A–gold complex for 30 min, thoroughly washed with PBS and distilled water, and stained. The protein A–gold complex was prepared with 10-nm gold particles as previously detailed (Bendayan 1995).
For immunodetection of endogenous serum albumin, we used the capillary tissue that was fixed by immersion. The thin sections were pretreated with sodium metaperiodate as mentioned above and then incubated with the anti-BSA antibody diluted 1:25 overnight at 4C. The protein A–gold complex was then applied for 30 min at RT. The crossreactivity of the anti-BSA with eel serum albumin was previously assessed by an immunodot experiment.
For immunodetection of actin, the tissue was fixed with 1% glutaraldehyde and processed for embedding in Lowicryl K4M at −20C without postfixation with osmium tetroxide (Bendayan 1983). This milder condition of fixation and the embedding in Lowicryl are required for optimal detection of cytoskeletal protein antigenic sites but lead, however, to suboptimal preservation of cellular structures (Bendayan 1983; Schnittler et al. 1990). For the protein A–gold immunolabeling, a rabbit anti-actin antibody (Bendayan 1983) was used at 1:50 dilution according to the protocol described above.
Several control experiments were performed to assess the specificity of the different immunolabelings. These included omission of the specific antibodies in the labeling protocol or adsorption of the specific antibodies with their respective antigens before the immunolabelings (Bendayan 1983, 1995; Bendayan and Rasio 1996a).
Results
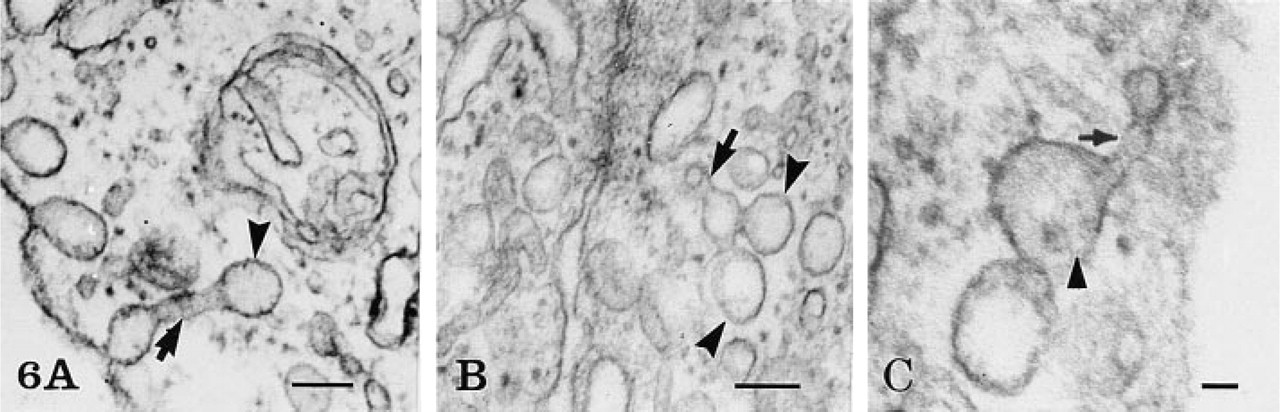
The rete mirabile of the eel swimbladder is composed of a rich pure network of blood capillaries arranged in parallel circuits. Arterial and venous capillaries alternate in regular fashion and carry blood in opposite directions. The arterial capillaries are connected at the poles of the rete by pre-retal and post-retal arteries, and the venous capillaries are connected by pre-retal and post-retal veins. In transmission electron microscopy, the arterial capillaries are formed by a high continuous endothelium, whereas the venous capillaries are thinner and fenestrated (Figure 1). Both endothelia rest on well-defined basement membranes and some pericytes surrounded by bundles of collagen fibers are found in the interstitial space (Figure 1). Tight intercellular junctions between endothelial cells seal the blood vessel lumina (Figure 2). At higher magnification (Figures 2–6), the endothelial cells display a tubular-vesicular system, composed of plasma membrane invaginations, which is particularly developed in the arterial capillaries. This system consists of vesicular and tubular membrane profiles that penetrate the endothelial cell cytoplasm and open at the luminal and abluminal spaces as well as at the level of the intercellular junctional space (Figure 2). Some of the tubules intrude deeply into the cell cytoplasm, forming channel-like structures (Figure 2). Observations made on tilting the tissue sections (20–30°) confirm the existence of true continuous membrane-bound tubular structures. In relatively thicker sections (Figure 3), these tubules can be followed over a path reaching several micrometers in length. The tubules can be straight or branched in various directions (Figures 2–6). Some of these branching structures form bottle-neck figures (Figures 4 and 4) because their narrow tubular connections have been cut out of the tissue section. The tubules either appear linear or exhibit a series of dilations and constrictions (Figures 2 and 2). In some instances, narrow tubular extensions are present, either arising from vesicular structures or extending between dilated vesicular profiles (Figure 6). In addition, narrower tubular structures are found close to the plasmalemmal membranes (Figure 5). Continuity of membranes is observed between the tubular structures and smooth membranes that could be assigned to the smooth endoplasmic reticulum (Figure 5). Occasionally, part of the membrane of this tubular-vesicular organelle appears coated (Figure 2). The continuity of membrane between the tubular system and the plasmalemmal membrane on both the luminal and abluminal fronts is evident. As reported previously (Sandborn et al. 1975), the tubular profiles appear to be particularly numerous in tissues fixed by the combined glutaraldehyde–DMSO solution, although in the absence of DMSO they are still present. Indeed, on osmium impregnation after fixation in glutaraldehyde (Figure 7) the tubular system was easily revealed, being highly contrasted by the electron-dense material. At low magnification (Figure 7A), the electron-dense delineated tubules appear as an intricate network with straight and dilated segments. Only occasionally was it possible to follow a tubule that appeared to extend from the luminal to the abluminal membranes (Figure 7B).
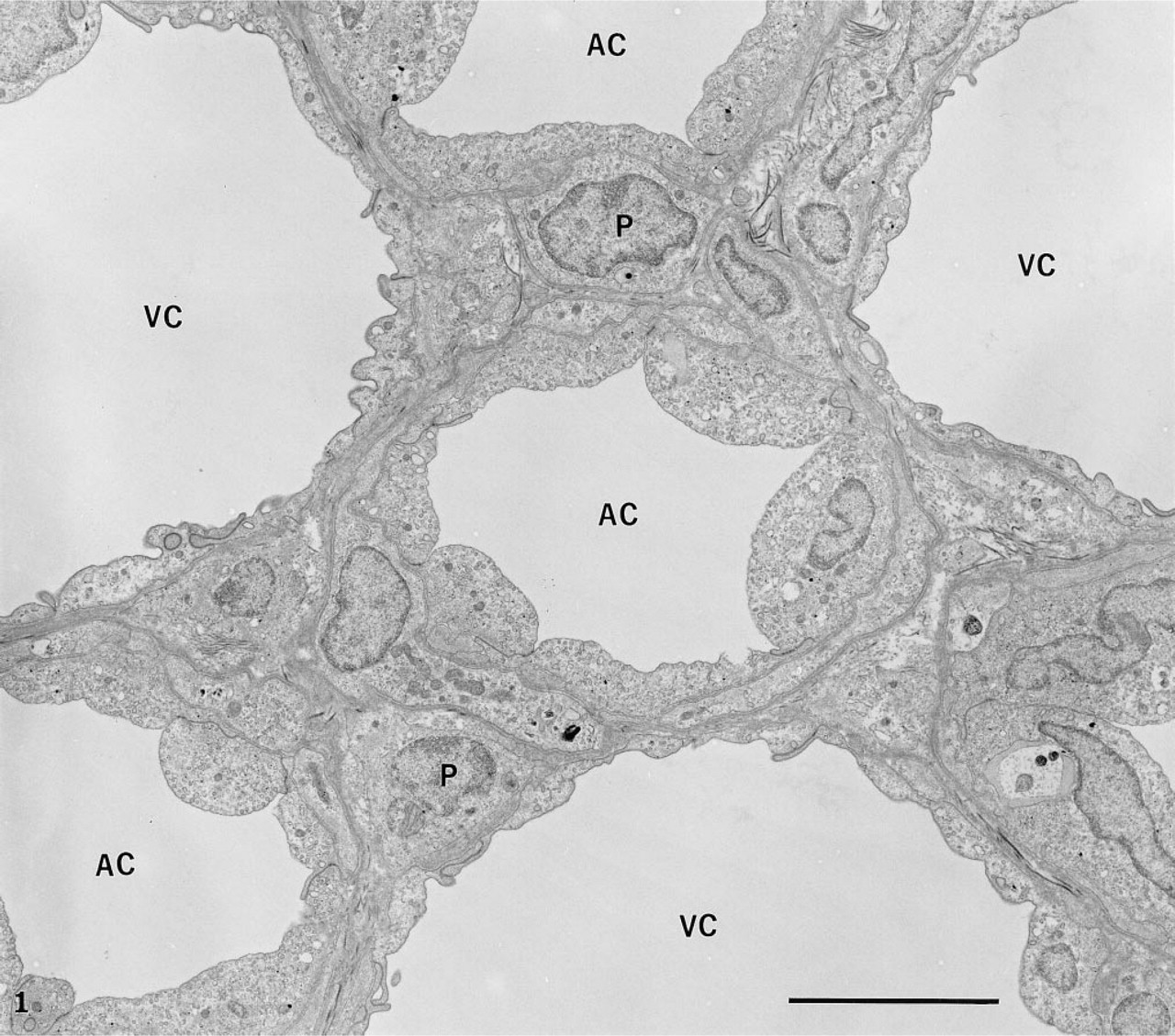
Transmission electron microscopy of the rete capillaries. The arterial (AC) and venous (VC) capillaries alternate. The arterial ones are composed of high endothelial cells, whereas the venous are thin and fenestrated. The intercellular space is limited to a few pericytes (P), the basement membranes, and some collagen fibers. Bar = 5 μ.
When prefixed tissues were perfused with peroxidase, the tracer was detected in association with the luminal plasma membrane, in the intracellular junctions as well as within the basement membrane (Figure 8). Extravasation of the tracer had therefore occurred, probably related to rupture of the interendothelial junctions as a consequence of the perfusion–fixation process. Nevertheless, vesicular and tubular profiles within the endothelial cells were labeled with the per-oxidase, demonstrating that after fixation these structures remained open to the luminal space (Figure 8). Most of these labeled structures were located at or close to the luminal plasma membrane. Occasionally, some vesicular profiles deep within the endothelial cells were also labeled, demonstrating that they are indeed connected to the luminal membrane. On the other hand, no peroxidase-labeled tubular profile was found connecting directly the luminal and abluminal fronts of the cells (Figure 8). Scanning electron microscopy at high magnification (Figure 9) revealed the existence of an endothelial intracellular network of interconnected tubules with dilations. The size of such structures falls within that of the tubulo–vesicular system observed by transmission electron microscopy. On replicas generated by freeze-fracture (Figure 10), long membrane-delineated intracellular tubules as well as vesicles are seen intruding deeply into the cell cytoplasm from the plasmalemmal membrane. The leaflets of these plasmalemmal membranes display their typical intramembrane particles. Replicas of lateral membranes also revealed the existence of tight occluding junctional elements composed of a few characteristic ridges or strands, as demonstrated previously (Rasio et al. 1993; Wagner and Kachar 1995).
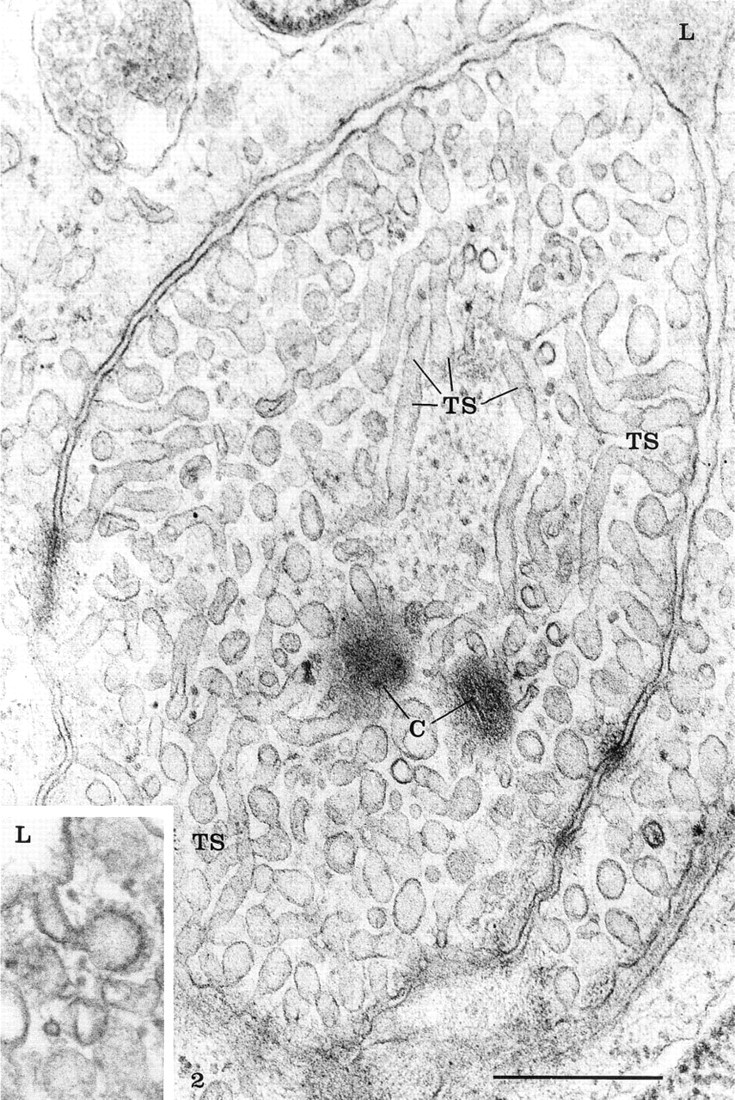
High magnification of an arterial endothelial cell illustrating the magnitude of the tubular system (TS). Membrane-bound tubular profiles are seen intruding into the endothelial cytoplasm from both the luminal and abluminal membranes as well as from the lateral membranes. Many of these tubules and vesicles are connected to the plasmalemmal membrane and open through a pore. Occasionally a diaphragm is seen at the level of the openings. Several tubules show branching connections on several directions and overlap.
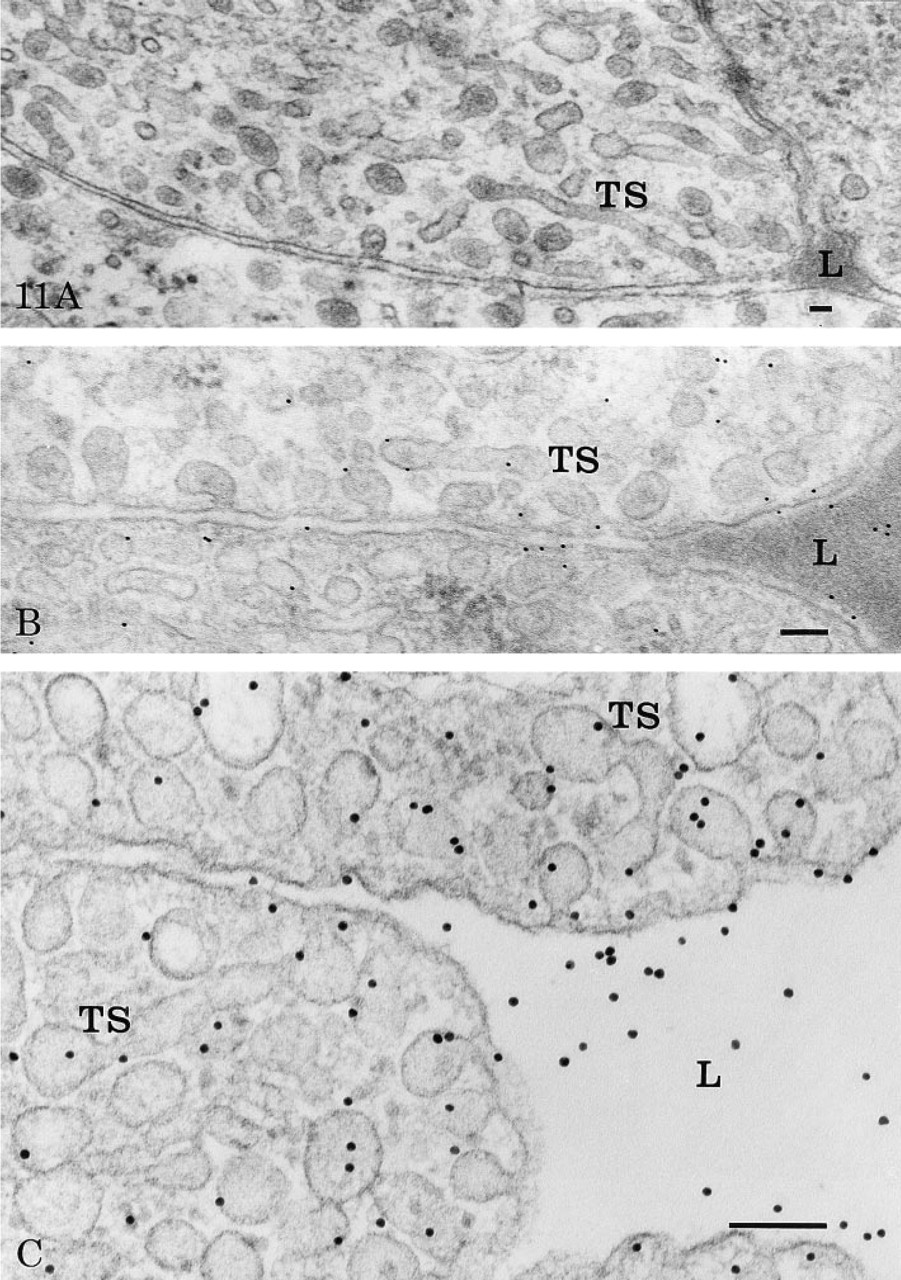
The particular arrangement of the rete capillaries allows countercurrent perfusion and accurate measurement of the transcapillary permeability of tracers present in the perfusate (Rasio et al. 1977). Application of immunocytochemical techniques with corresponding specific antibodies reveals that the path taken by the tracer proteins crosses the capillary wall. In a first series of experiments, we revealed endogenous serum albumin within the capillary tissues. For this, immersion-fixed tissues were used to retain blood within the capillary lumina. This is reflected by the presence of nucleated red blood cells surrounded by an electron-dense plasma (Figure 11A). In such tissues, the vesicular and tubular profiles of the endothelial cells frequently appeared to be filled with a dense material resembling that present within the capillary lumen (Figure 11A). The basement membranes as well as the interstitial space also appeared denser than on perfused-fixed tissues. To reveal endogenous eel serum albumin, we used an anti-BSA antibody. The cross-reactivity of this antibody with eel serum albumin was previously assessed and demonstrated by an immunodot experiment. The results have shown that eel albumin, although not as immunoreactive as bovine albumin, is still well recognized by the anti-BSA antibody at dilutions up to 1:500 (results not shown). Application of this anti-BSA antibody with the protein A–gold complex on such immersed-fixed tissue led to labeling by gold particles over the dense content of the capillary lumen, the interstitial space, and over the vesicular and tubular profiles (Figure 11B). These labeled structures were located throughout the cell, close to both the luminal and the abluminal plasma membrane. Such labeling reflects the presence of endogenous serum albumin within those compartments and demonstrates the transendothelial transport of endogenous serum albumin by the vesicular and tubular system.
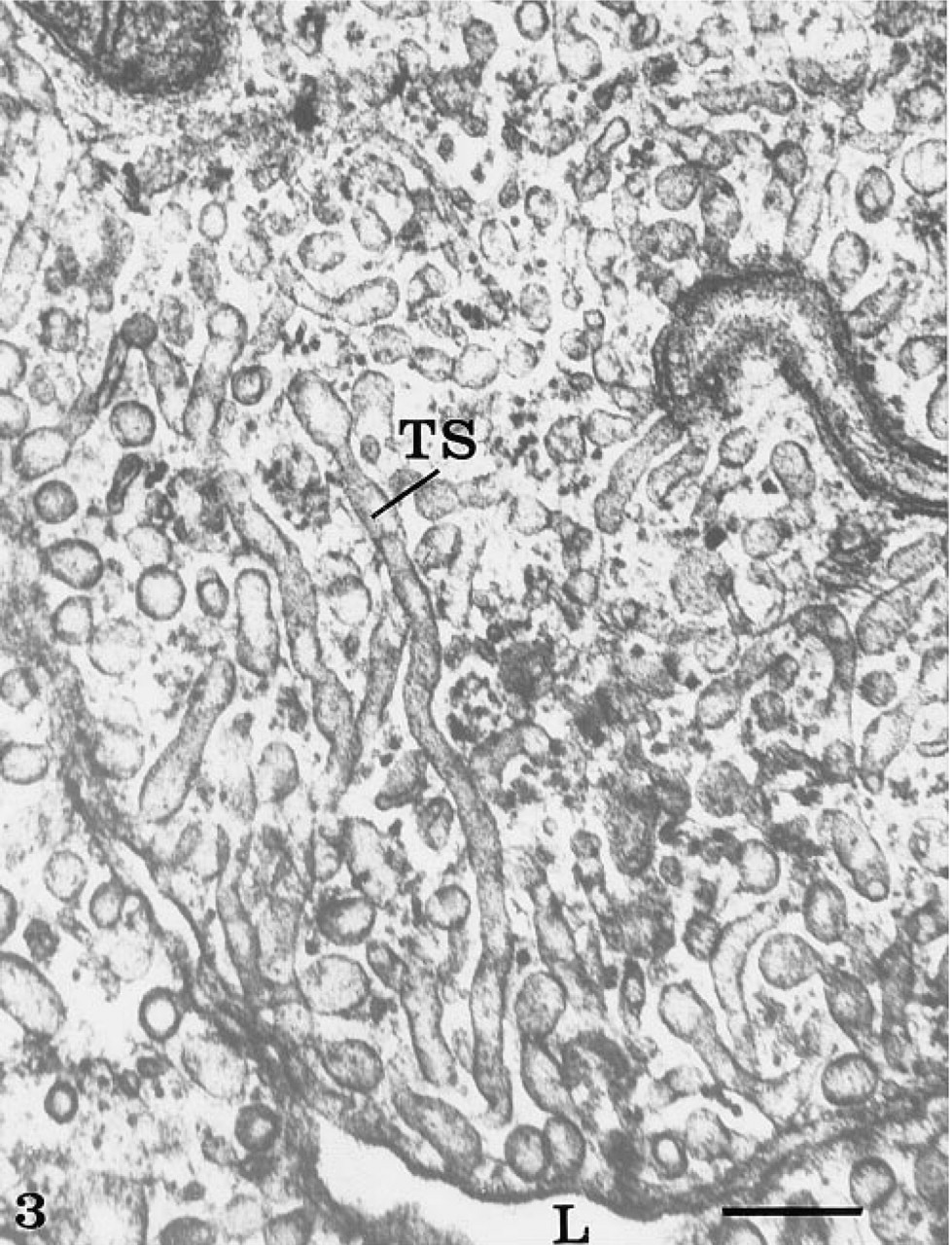
Thick section of an arterial endothelial cell, illustrating the complexity of the tubular network. Some tubules (TS) intrude deeply into the cytoplasm and are more than 1 μ in length. Bar = 0.5 μ.
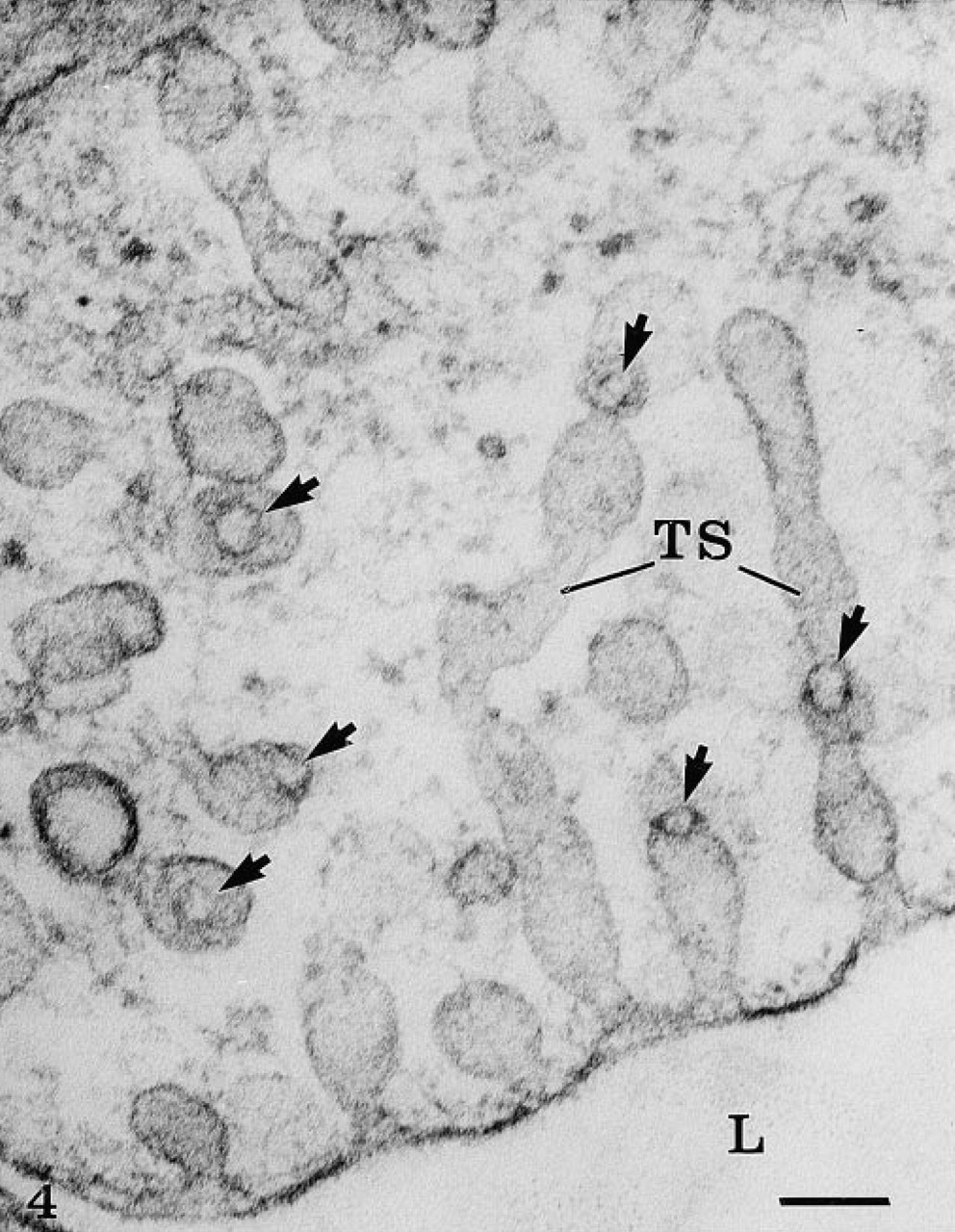
High magnification of several tubules (TS) demonstrating their arborization and the circular profile of the cut edges (arrows) caused by the sectioning process. Bar = 0.1 μ.
In a second series of experiments we used bovine serum albumin and porcine insulin as exogenous tracers. These tissues were perfused for 30 min as described in Materials and Methods. The specific antibodies were combined with the protein A–gold complex to reveal the location of their corresponding antigens in the capillary tissue sections with high resolution. Aside from variations in intensities of labeling due to differences in concentrations of the tracers and antibody titers, the pattern of labeling was similar for both proteins. Because the perfusions were carried out until steady-state conditions were reached, the labeling by gold particles reflecting the location of the tracers was found in both the arterial and the venous capillary lumina, in the endothelial cells, and in the interstitial space. In the arterial endothelial cells, the labeling was mainly located over the tubular–vesicular structures, in those opened at both the blood and tissue fronts of the cells as well as in those present inside the cell cytoplasm (Figure 11C). Very few gold particles were seen over mitochondria or nuclei. Notably, the intercellular junctions were devoid of any labeling. When labeling was found close to the junctions, it was confined to either the luminal or the basal side of the junctions but not at the level of the tight occluding complex. In the intercellular space, the labeling was located within the basement membranes and between the collagen fibers. Little labeling was detected in pericytes and, when present, it was associated with vesicular profiles (data not shown).
For the study of actin, a different set of experiments was performed in which actin antigenic sites were detected on immersion-fixed nonosmicated tissues embedded in Lowicryl. Because of these conditions of tissue preparation, the ultrastructural preservation was suboptimal. Strong labeling for actin was found on pericytes, particularly over filament-rich regions (data not shown). The labeling was also present in endothelial cells, but to a lesser extent. It was concentrated along the plasmalemmal membrane (Figure 12A), the intercellular junctional complex, and associated with the membranes of the vesicular and tubular profiles (Figure 12B).
Control experiments for the immunocytochemical studies resulted in an absence of specific labeling, very few gold particles being present in the capillary lumina and endothelial cells (data not shown).
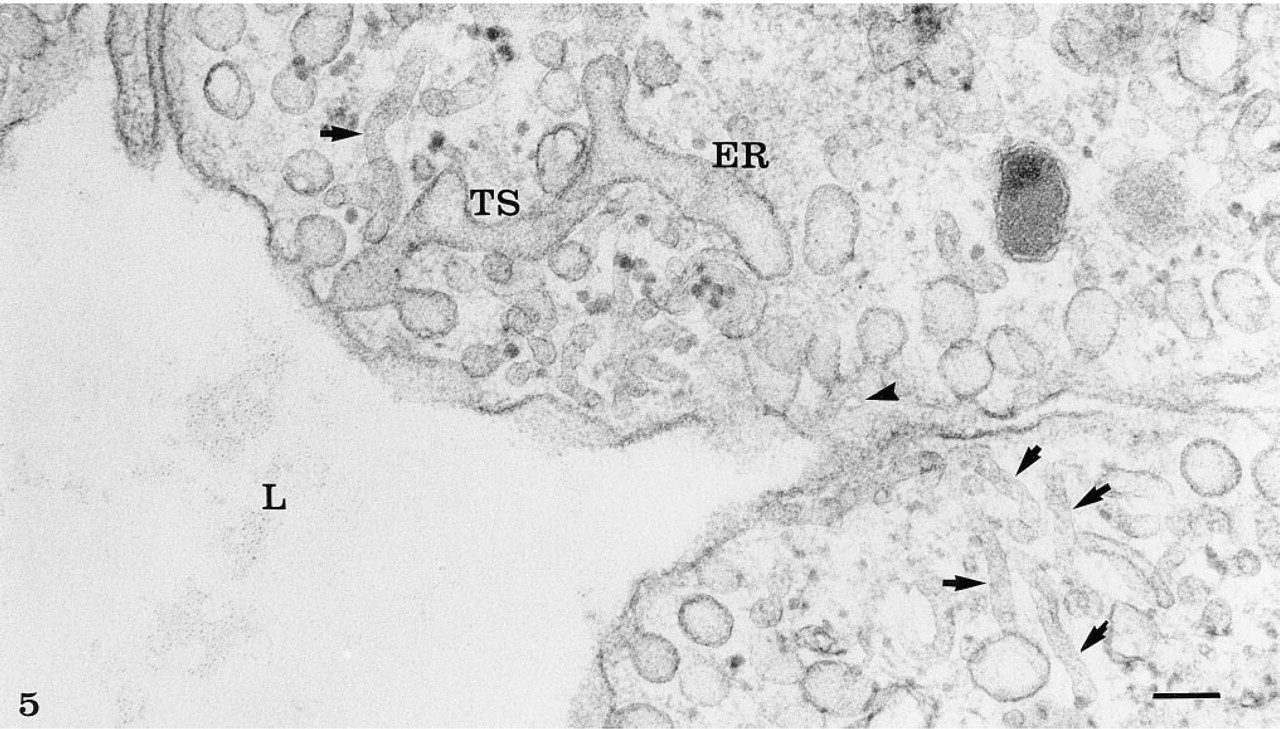
The tubular system (TS) opening at the luminal plasmalemmal membrane appears continuous with smooth cell membranes that could be assigned to the smooth endoplasmic reticulum (ER). Narrower tubules (arrows) are also seen close to the luminal membrane. Arrowhead indicates the opening of the vesicular profiles at the luminal membrane, as seen in front view. Bar = 0.1 μ.
(
Discussion
The rete mirabile of the eel swimbladder consists of a rich network of blood capillaries, the function of which is the build-up of the partial pressure of oxygen, allowing its diffusion into the swimbladder. This increase in oxygen pressure is achieved by a hairpin countercurrent multiplication mechanism that pumps lactic acid from the venous capillaries into the arterial ones. The resulting acidification of the arterial blood at the bladder pole of the rete raises the partial pressure of oxygen, which causes the gas to enter the bladder (Steen 1963). The arterial capillaries of the rete are continuous with a thick endothelium, and the venous ones are fenestrated. Both rest on well-defined basement membranes and are partially surrounded by pericytes. All the features of these endothelia have been reported to be similar to those of mammalian capillaries (Bendayan et al. 1975). Functional studies have also demonstrated that permeability coefficients of various molecules, including water, are of the same order of magnitude as those of the mammalian blood–brain barrier (Bolwig and Lassen 1975; Rasio et al. 1993). The endothelial cells, the arterial ones in particular, display a very well-developed vesicular–tubular system formed by membrane-bound vesicles and tubules that intrude deeply into the endothelial cells from both their luminal and abluminal poles. This tubular system also opens at the level of the lateral intercellular space. It branches in different directions, forming a network that appears on thin sections as tree-like structures. Occasionally, connections with the smooth endoplasmic reticulum are also observed and could relate to the close appositions reported to occur in other endothelial cells between cisternae of endoplasmic reticulum and caveolae (Bundgaard 1991). Such connections between organelles lead to the formation of a complex cellular microcirculatory system in endothelial and other cell types (Sandborn and Bendayan 1996). Studies on thick tissue sections and after osmium impregnation have demonstrated the magnitude and intricacy of this system. In addition to its unusual length and its deep intrusions inside the endothelial cell cytoplasm, the tubular system shows successive dilations and constrictions, particularly evident in transmission and scanning electron microscopy, which give the impression that the tubules are involved in dynamic peristaltic movement. In keeping with this, we have demonstrated the presence of actin associated with the tubular and vesicular membranes. Indeed, actin was detected not only in close association with the plasmalemmal membrane corresponding to the cortical filamentous cell web and the junctional complex, as reported previously (Schnittler et al. 1990), but also with the limiting membrane of vesicular and tubular profiles. These cytochemical results are consistent with the previous demonstration of actin and actin binding proteins associated with plasmalemmal vesicles or caveolae (Izumi et al. 1988; Lisanti et al. 1994) and with the requirement of an intact actin network for the functional properties of caveolae (Parton et al. 1994). As proposed for secretory cells and fibroblasts, in which actin was detected in close association with the membrane of vesicular structures and secretory granules (Bendayan 1983), the presence of actin at the level of the tubule and vesicle membranes would provide contractile properties to the system. Our cytochemical approach for the study of vascular permeability takes advantage of the good ultrastructural morphology provided by the combined use of glutaraldehyde and osmium tetroxide, of the very rapid immersion–fixation of the tissues which retains the tracer proteins in situ, and of the postembedding immunocytochemical approach with the use of gold particles, to generate labeling of high resolution over well-preserved cellular structures. Furthermore, the morphological approach chosen permits the use of optimal physiological conditions for the study of vascular permeability of either endogenous serum proteins or tracer serum proteins in their native form (Bendayan 1980; Bendayan and Rasio 1996a), and has demonstrated the involvement of the tubular system in the transcapillary transport of circulating molecules.
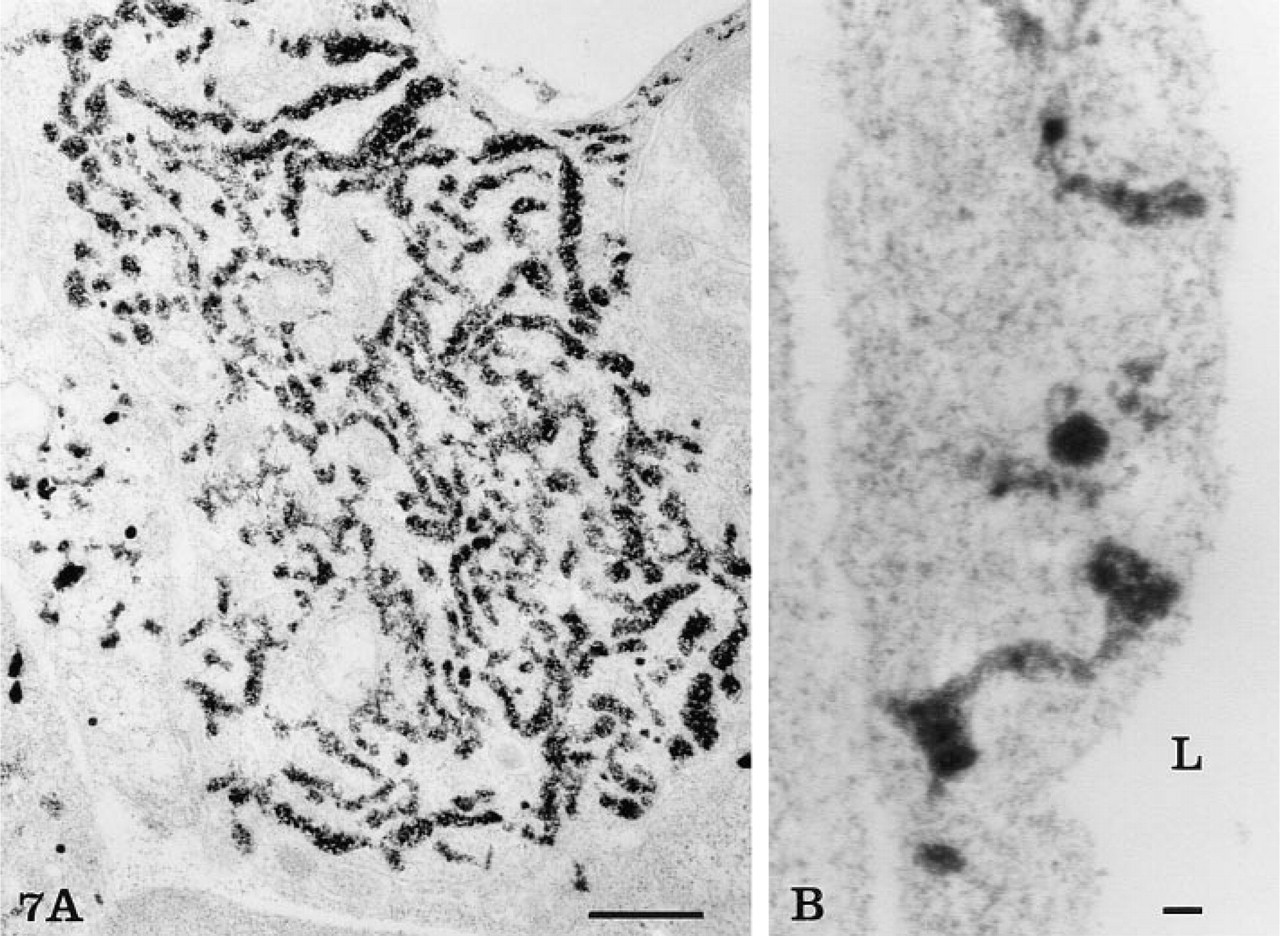
Osmium impregnation. At low magnification (
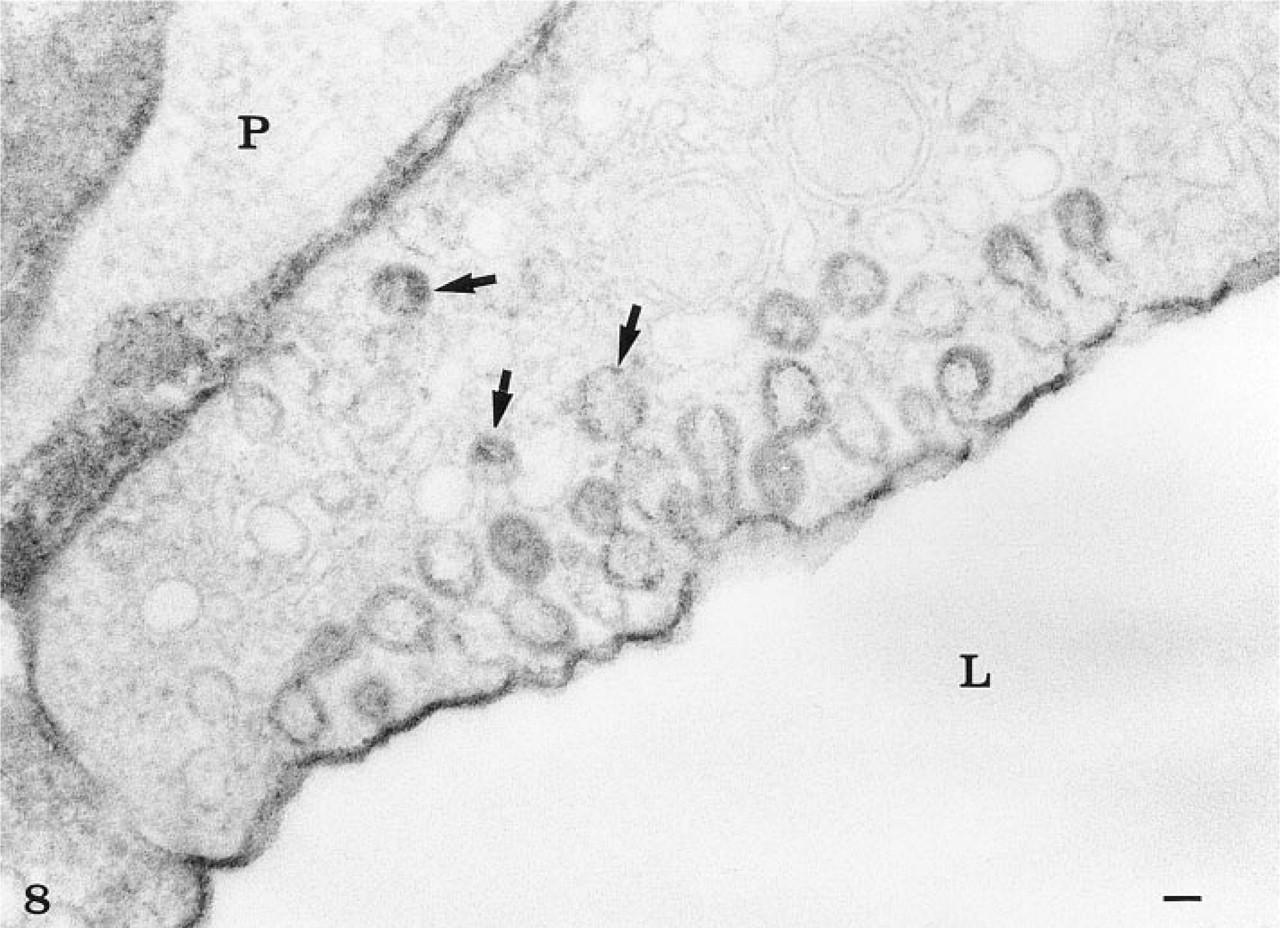
Peroxidase experiment. The capillary tissue was first fixed by perfusion with 1% glutaraldehyde, then perfused for 30 min with 1% horseradish peroxidase and then fixed again by immersion in 1% glutaraldehyde. The peroxidase was revealed by the DAB reaction. The cytochemical reaction product is associated with the luminal plasma membrane and the basement membrane. It is also present at the level of the intercellular junction. Vesicular and tubular profiles within the endothelial cell, particularly those located at or close to the luminal membrane, are also labeled. Some (arrows) located deep within the cytoplasm also appear to contain the reaction, demonstrating that even after fixation the peroxidase had access to those structures. P, pericyte. Bar = 0.1 μ.
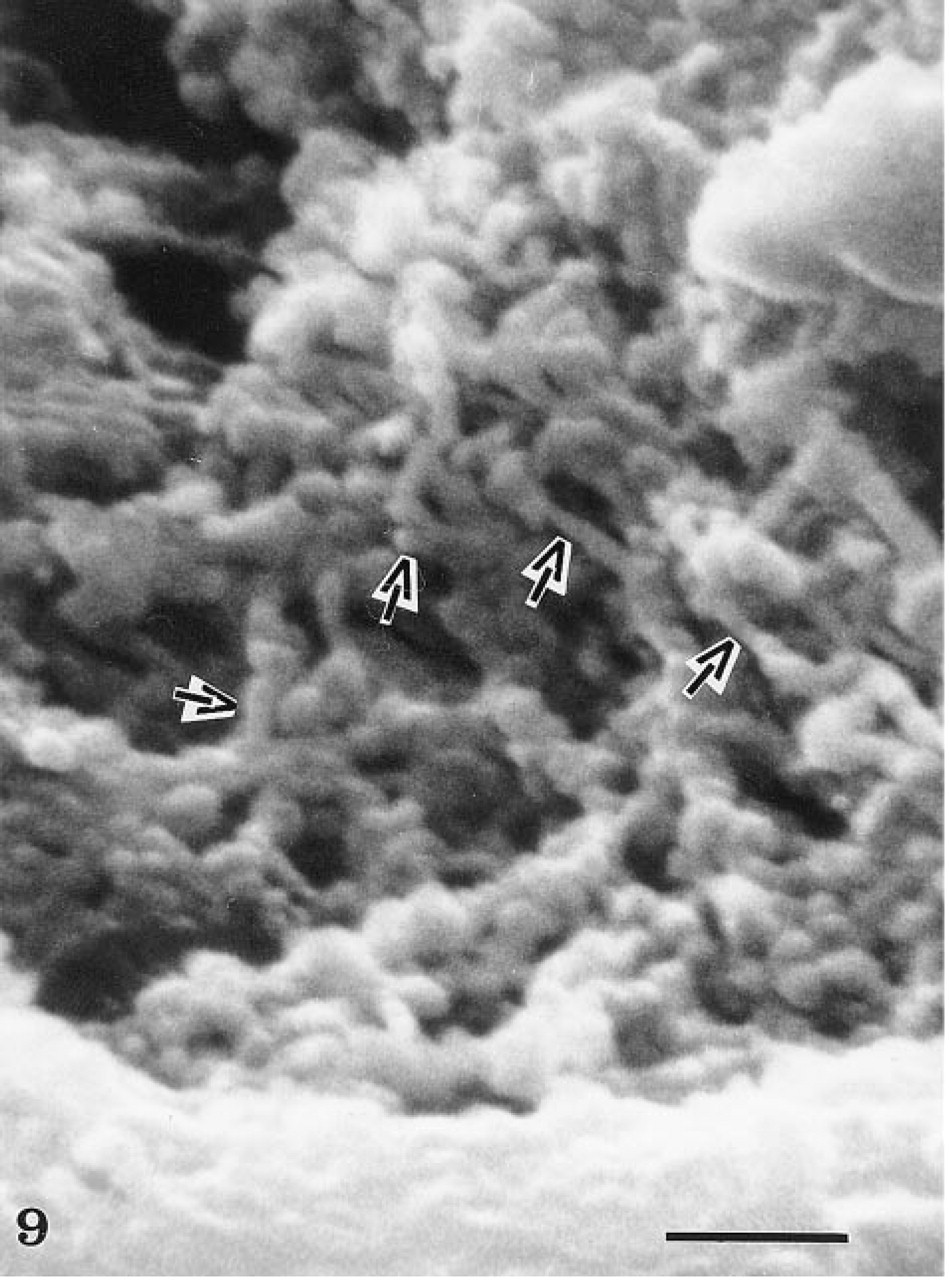
High-magnification scanning electron micrograph illustrating the tubular network (arrows) within the endothelial cell cytoplasm. Sporadic dilations of the tubular system are evident. Bar = 0.5 μ. (From Can J Physiol Pharmacol 53:613–623, 1974, with permission.)
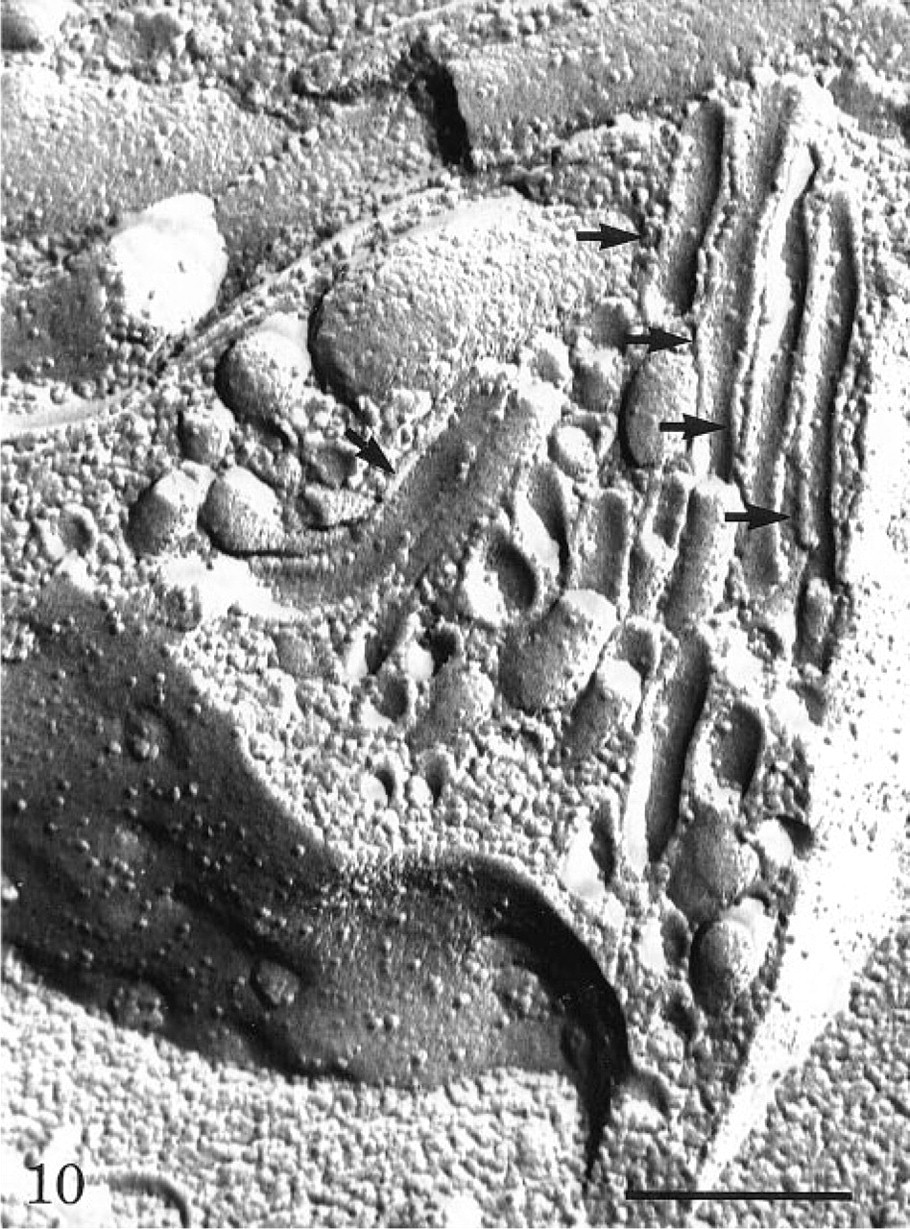
Freeze-fracture replica of an endothelial cell demonstrating many membrane-bound tubules (arrows) intruding into the cell, particularly from the abluminal plasmalemmal membrane. Intramembrane particles are seen on the luminal membrane leaflet. Bar = 0.5 μ.
(
The presence of tubules in the rete capillaries in such a significant number might be attributed to the large size of these endothelial cells and to the very active countercurrent transport taking place in these capillaries for adaptation of fish buoyancy to water depth (Steen 1963). The existence of similar deep membrane invaginations, long vesicular structures as well as transendothelial channels in endothelial cells of various vascular beds, has been reported extensively (Wolff 1966; Hashimoto 1972; Zweifach 1972; Simionescu et al. 1975; Bendayan and Sandborn 1976; Clough and Michel 1981; Siminonescu 1983; Shivers et al. 1984; Lossinsky et al. 1989; Wagner and Chen 1991; Kohn et al. 1992). Their function has been a subject of debate because the general consensus for transendothelial transport tends to favor a system of membrane-bound independent vesicles (Palade 1960, 1961; Karnovsky 1967; Schneeberger 1983; Simionescu 1983; Schnitzer et al. 1994, 1996). This vesicular theory, although also denied by Bundgaard (1983), presupposes the formation of vesicles arising from the plasmalemmal membrane at one front of the cell, their pinching off, and their migration across the endothelial cytoplasm towards the opposite pole of the cell for their final fusion with the plasmalemmal membrane (Palade 1960, 1961; Milici et al. 1987; Ghitescu and Bendayan 1992; Schnitzer et al. 1996). Such a shuttle process of isolated vesicles implies the generation of large amounts of vesicular membrane at one plasmalemmal front and their addition at the opposite front. With the existence of a dynamic tubular system formed by a network of membrane-bound tubules with the potential of opening on all sides of the cells, there is no need for continuous generation and disposal of vesicular membranes. Because cytoskeletal proteins are associated with the membranes of the tubular system, transendothelial transport of serum proteins could take place through this system by contractile (peristaltic) activity. The content of the tubules would be propelled along the rather stable membranes of the tubules. In such a system, there would be no translocation of membrane, the content of the tubules would remain extracytoplasmic, and the surface of interchange between the extracytoplasmic (extracellular) milieu and the cell cytoplasm itself would be increased severalfold. The controversies concerning tubules vs vesicles have been centered on tissue preparation artifacts. Indeed, breakdown of tubules could give rise to a series of vesicles. Conversely, the tubules and trans-endothelial channels could arise from elongated vesicles after their formation at the plasmalemmal membrane, with subsequent fusion processes (Simionescu et al. 1975). The breakdown of tubules into series of vesicular structures has been demonstrated repeatedly in various cell types as a direct consequence of fixation artifacts (Rosenbluth 1963; Tormey 1964; Mersey and McCully 1978; Wilson et al. 1990; Hyde et al. 1991; Sandborn and Bendayan 1996). Formation of tubules by fusion of vesicles is unlikely when the length of the tubules reaches several micrometers. This would require the synchronized fusion of a tremendous number of aligned vesicles. In addition, if fusion of vesicles were the generating event, one would not expect it to occur unidirectionally, leading to tubular structures, but rather in all directions, leading to large vacuoles. In support of the existence of tubular systems are results obtained from cryofixation–cryosubstitution experiments in which cellular structures are rapidly immobilized (Frokjaer–Jensen et al. 1988). In such tissues, as well as in those fixed in the presence of tannic acid (Bundgaard et al. 1979), virtually all the apparent free plasmalemmal vesicular profiles present in endothelial cell cytoplasm were reported to be interconnected, forming an integral part of channel-like structures (Bundgaard et al. 1979; Frokjaer–Jensen et al. 1988). In addition, in permeability studies it was found that tracer proteins of different molecular weights display differences in permeability coefficients, indicating that transport is compatible with diffusion processes rather than with vesicular bulk quanta movement from one cell front to the other (Bundgaard 1980; Bendayan and Rasio 1996a, b). Moreover, estimations made by Paaske and Sejrsen (1977) to account for albumin transport through vesicular shuttling show that this would require a tremendous amount of energy. This goes along with the theory that transfer of molecules through vesicular shuttling, even among cellular compartments, would require energy supplies far superior to those available to the cell (Ling 1969). Cytoplasmic tubular systems are not limited to endothelial cells nor to the transcytotic activity. They have been reported in other cell types, including the secreting parietal cells in the gastric mucosa (Sugai et al. 1985; Pettitt et al. 1995,1996). They have been involved in endocytic processes as part of the endosomal compartment in several epithelial cells (Marsh et al. 1986; Kobayashi and Robinson 1991; Tooze and Hollins-head 1991; Luo and Robinson 1992; Myers et al. 1993; Sandborn and Bendayan 1996; van Deurs et al. 1996). In this context, one should keep in mind that endothelial cells are a specialized form of epithelial cell in which endocytosis is mainly targeted to transcytosis. In addition to the endosomal compartment, tubular structures are also well established in the lysosomal system of various cell types (Oliver 1983; Beaudoin et al. 1985; Robinson et al. 1986; Sandborn and Bendayan 1996).
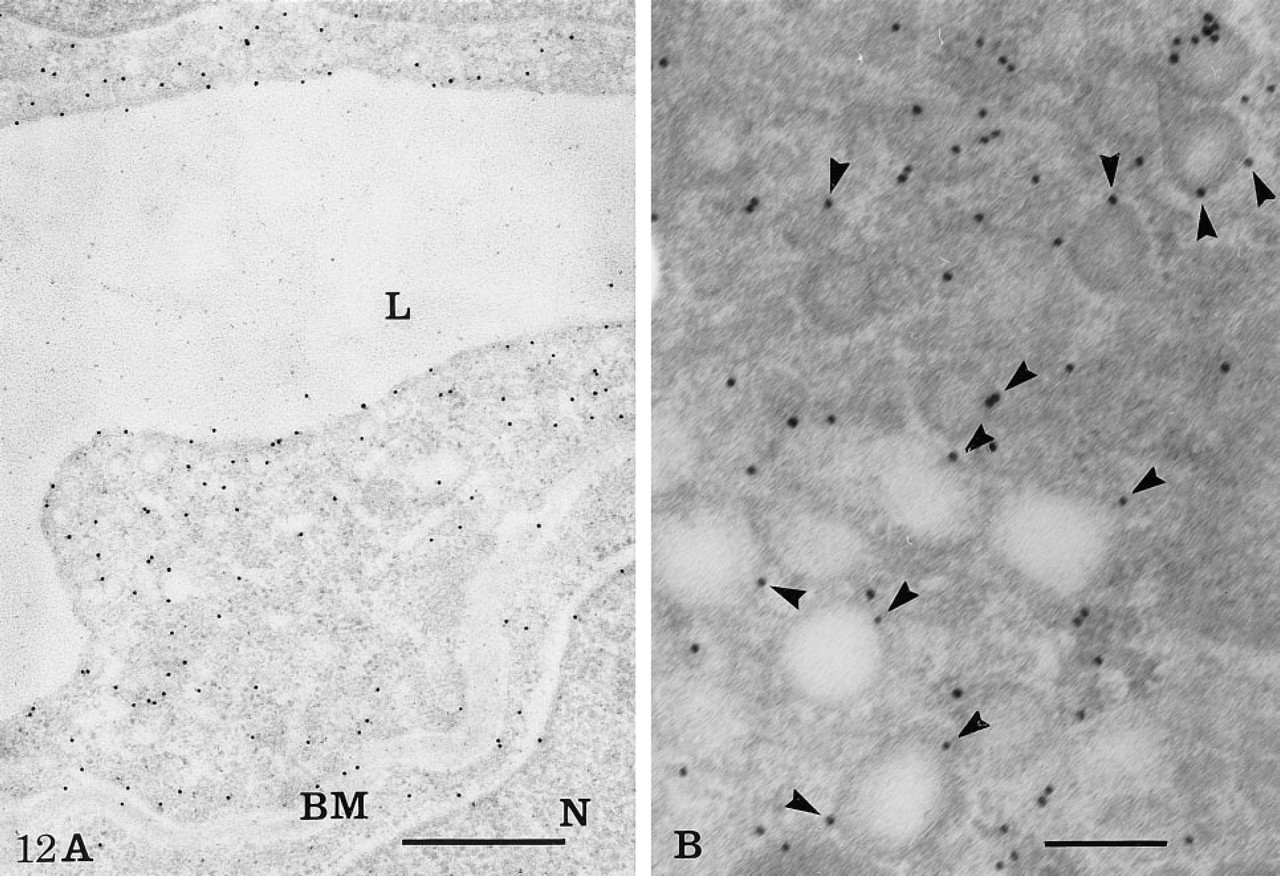
Immunocytochemical demonstration of actin in the rete capillaries. After fixation with glutaraldehyde and embedding in Lowicryl, actin antigenic sites were detected by a postembedding step on thin sections of the capillary tissue. At low magnification (
With respect to the functional properties of the tubular system in endothelial cells, we have shown the presence of circulating endogenous and exogenous albumin, insulin, and large serum proteins of various sizes (Bendayan and Rasio 1996a, b) within the tubules and their absence at the level of the intercellular junctions. Furthermore, physiological permeability studies using radiolabeled tracers have clearly demonstrated the existence of transendothelial transport across the capillary wall (Rasio et al. 1977, 1993; Bendayan and Rasio 1996a). The combined morphocytochemical and physiological studies indicate that large serum proteins are transported across the capillary wall through the tubular system. The selectivity in transendothelial transport, like the one demonstrated for insulin in the rete capillaries (Bendayan and Rasio 1996a), may be governed by the existence of many tubules within a given cell that could be related and could respond to specific binding sites existing at the plasmalemmal membrane. Several receptors to circulating serum proteins have been found on the plasmalemmal membrane (Bar et al. 1978; Vasile et al. 1983; Jefferies et al. 1984; King and Johnson 1985; Solenski and Williams 1985; Ghitescu et al. 1986; Schnitzer 1992). Active transport could therefore take place along tubules that would form or open only on specific signals triggered by ligand–receptor interactions. The tubular system need not be a permanently open channel for the free diffusion fluid-phase transport but could rather be an active system responding to specific membrane signals. In this context, confirming previous studies (Balin et al. 1987; Broadwell 1989), our peroxidase experiment was unable to demonstrate open static channels connecting luminal and abluminal fronts of the cells, channels through which transport could take place by simple diffusion. Therefore, the large pore transport system previously proposed to account for the transendothelial transport of large proteins might be represented by this tubular system. Its involvement in the concomitant transport of small molecules and its contribution to the small pore system, as defined by physiological means, could be predictably important and needs to be assessed.
Footnotes
Acknowledgements
Supported by research grants from the Medical Research Council of Canada.
We are grateful to Dr. L. Ghitescu for his interest and constructive discussion. We acknowledge the technical assistance of G. Mayer, D. Gingras, M.P. Dea, and J. Léveillé.