
Abstract
Group II phospholipase A2 (PLA2) has been proposed to play an important role in inflammation and defense against bacterial infection. We investigated tissues of transgenic mice expressing the human group II PLA2 gene by immunohistochemistry using rabbit anti-human group II PLA2 antibodies, and by in situ hybridization by probing with human group II PLA2 mRNA anti-sense (test) and sense (control) riboprobes. By immunohis-tochemistry, human group II PLA2 was found in various mouse tissues and cell types including hepatocytes, proximal tubule cells of the kidney, epithelial cells of the renal pelvis, urinary bladder and ureter, granulosa cells of Graafian follicles, aortic intima and media, cartilage, epiphyseal bone, bronchial epithelial cells, and connective tissue cells in the dermis. By in situ hybridization, group II PLA2 mRNA was localized in hepatocytes, epidermal cells, dermal cells, connective tissue fibroblasts, epithelial and smooth muscle cells of the urinary bladder, and cells of Bowman's capsule. These results show that human group II PLA2 is expressed in large amounts in hepatocytes and many extrahepatic tissues of the transgenic mice. These animals provide a useful new tool for studies on the metabolism, in vivo effects, and physiological and pathological roles of phospholipase A2.
Keywords
P
It has been proposed that group II PLA2 is an acute phase protein produced by hepatocytes (Crowl et al. 1991) and that the enzyme plays an important role in inflammation (Vadas et al. 1993). However, studies on the site of production and the role of PLA2 and its inhibition by pharmacological interventions in various disease processes have been hampered by the lack of suitable experimental animal models in which to study the expression of the human group II PLA2. For this purpose, transgenic mice expressing the human group II PLA2 gene were recently generated (Grass et al. 1996). The aim of the present study was to investigate the distribution of human group II PLA2 by immuno-histochemistry and to localize the expression of the human group II PLA2 gene by mRNA in situ hybridization in tissues of transgenic mice.
Materials and Methods
Production of Transgenic Mice
Transgenic mice were produced by standard methods as described in detail elsewhere (Grass et al. 1996). Briefly, a 6.2-kb HindIII restriction fragement containing the human group II PLA2 gene (Kramer et al. 1989) was prepared and micro-injected into (C57BL/6 x SJL) F2 hybrid one-cell mouse embryos, which were then transferred to pseudopregnant ICR mice and developed to term. The transgenic founders were bred to C57BL/6J mice to produce G1 animals.
Immunohistochemistry
Tissues from various organs of three transgenic mice and one control mouse were fixed in 10% phosphate-buffered formalin and embedded in paraffin. Sections were collected on polylysine-coated slides, reacted with an IgG fraction (4.5 mg/ml) of a rabbit anti-group II PLA2 antiserum (Nevalainen et al. 1993), diluted 1:10,000 with 0.15 NaCl in 0.05 M Tris-buffer, pH 8.6, containing 1 % bovine serum albumin, and the primary immunoreaction was localized (Hsu et al. 1981) by using a Vectastain avidin–biotin-peroxidase complex (ABC) kit (Vector Laboratories; Burlingame, CA) according to the manufacturer's instructions. The slides were lightly counterstained with hematoxylin. For controls, the primary antibody was replaced by preimmune rabbit serum.
In Situ Hybridization
In situ hybridization was performed on sections of formalin-fixed, paraffin-embedded tissues by probing with group II PLA2 anti-sense (test) and sense (control) single-stranded RNA-probes as described earlier (Nevalainen et al. 1995), with some modifications. Briefly, a 0.45-
Results
Tissues from three transgenic mice (male, 6.5 months of age, line 854, #1300; female, 4.5 months of age, line 703, #2059; male, 3 weeks of age, line 854, #3532) and one control mouse (female, 4.5 months of age, #2063) were studied by immunohistochemistry (Table 1) for human group II PLA2 enzyme protein and by in situ hybridization (Table 2) for the mRNA of human group II PLA2.
Liver
There was a strong immunoreaction in the portal areas of the liver lobules. Hepatocytes were intensely stained, whereas other cell types, including Kupffer cells, were unstained (Figure 1). There was no immunoreaction in the liver or other tissues of the non-transgenic control mouse (Figure 2). Similarly, there was no immunostaining when the primary antibody was replaced by preimmune rabbit serum. The in situ hybridization signal for the mRNA of group II PLA2 (anti-sense riboprobe) was evenly distributed over hepatocytes (Figure 3). In situ hybridization with the control (sense) riboprobe gave negative results in liver (Figure 4) and other tissues. In situ hybridization with both anti-sense and sense riboprobes gave negative results also in the tissues of the non-transgenic control animal.
Urinary Organs
Bowman's capsule in a number of glomeruli of the transgenic animals showed positive immunoreaction (Figure 5). There was granular immunoreaction in the apical cytoplasm of proximal tubule cells (Figure 6). Epithelial cells of the renal pelvis, ureter, and urinary bladder were also strongly immunostained. In situ hybridization gave a clear signal in Bowman's capsule but proximal tubules were devoid of the mRNA of group II PLA2 (Figures 7 and 8). The epithelium of the urinary bladder gave a strong in situ hybridization signal, and there was weaker positive signal for the mRNA of group II PLA2 in the smooth muscle cell layer of the bladder (Figures 9 and 10).
Skin
There were many strongly immunoreactive, elongated connective tissue cells in some sections of the dermis (Figure 11). Basal cells of the epidermis showed weak immunoreaction. There was a distinct in situ hybridization signal in the epidermis (Figure 12). Fibroblasts, histiocytes, and mast cells (identified by metachromasia in Toluidine blue-stained slides) were found in the dermis (Figure 13) in areas containing cells with positive immunoreaction for group II PLA2 (Figure 11). In these areas there was strong in situ hybridization signal for the mRNA of group II PLA2 (Figure 14). However, it was not possible to resolve unequivocally which cell type in the dermis contained the mRNA of group II PLA2. Fibroblasts in the loose connective tissue of the neck showed positive in situ hybridization signal for the mRNA of human group II PLA2 (Figure 15).
The spleen, pancreas, salivary glands, and Paneth cells of the small intestinal mucosa contained no immunoreactive material. The mRNA of human group II PLA2 was localized in the smooth muscle cells of the small intestinal wall by in situ hybridization, but Paneth cells were devoid of signal.
Immunohistochemical detection of human group II PLA2 in tissues of transgenic micea
Reproductive Organs
The granulosa cells of the Graafian follicles of the ovary contained immunoreactive material (Figure 16). The Fallopian tube and uterus were devoid of immunoreaction. There was neither immunoreaction nor in situ hybridization signal in the testis or epididymis.
More recently, in situ hybridization has been performed on other line 703 animals (a female, age 3 months and two males, age 2 months and 16 months). These results (data not shown) confirm the presence of the mRNA of human group II PLA2 in Graafian follicles and its absence in the testis.
In situ hybridization of the mRNA of human group II PLA2 in tissues of transgenic micea
Circulatory Organs, Muscle, and Supporting Tissues
The intima and media of the aorta were strongly immunostained (Figure 17). However, there was no in situ hybridization signal in the aorta. The heart and skeletal muscle cells were devoid of immunoreactive material. Connective tissue fibers and tendon were occasionally weakly immunoreactive. The adipose tissue was devoid of immunostaining. Fibroblasts showed positive in situ hybridization signal (Figure 15). Articular cartilage and epiphyseal bone were immunoreactive (Figure 18). There was strong immunoreaction in the chondrocytes and a weaker reaction in the matrix of articular cartilage (Figure 19). Cortical bone was devoid of immunoreaction. In situ hybridization gave negative results in bone and cartilage.
Lung
Some bronchial epithelial cells contained immunore-active material, whereas other cell types of the lung were devoid of immunoreaction. There was a weak in situ hybridization signal in occasional bronchial epithelial cells.
The cells of lymph nodes, bone marrow, and peripheral inflammatory cells were devoid of immunore-active material. Neither the brain nor peripheral nerves showed immunoreaction. In situ hybridization gave negative results in bone marrow and neural tissues.
The immunohistochemical results are summarized in Table 1. The immunoreaction for group II PLA2 in hepatocytes was somewhat weaker in the younger animal (3 weeks, male) than in the older animal (6.5 months, male) of the same transgenic line. The in situ hybridization results are summarized in Table 2.
Discussion
Phospholipase A2 has been implicated in the pathology of diseases involving inflammation (Vadas et al. 1993). Increased concentrations of group II PLA2 have been measured in serum in various diseases including acute pancreatitis, sepsis, infections, rheumatoid arthritis, and multiple injuries (Nevalainen 1993). It has been proposed that group II PLA2 is an acute phase protein (Crowl et al. 1991).
The cellular source of circulating group II PLA2 has not been established unequivocally. Human hepatoma-derived cells were found to express increased amounts of group II PLA2 when stimulated in vitro by the cytokines interleukin 1, interleukin 6, and tumor necrosis factor (Crowl et al. 1991). Enhanced expression of group II PLA2 was demonstrated in human hepatocellular carcinoma cells and surrounding liver tissue (Ying et al. 1994). Expression of group II PLA2 was recently demonstrated in the non-neoplastic hepatocytes of a patient who had an epitheloid hemangioendothelioma of the liver (Nevalainen et al. 1996). Group II PLA2 has been detected by immunohistochemistry (Kiyohara et al. 1992; Nevalainen and Haapanen 1993), and the mRNA of group II PLA2 by Northern blotting (Kramer et al. 1989) and in situ hybridization (Nevalainen et al. 1995), in many tissues other than the liver. In addition to hepatocytes, other potential sources for group II PLA2 include the Paneth cells of the small intestinal mucosa (Nevalainen et al. 1995), cartilage (Nevalainen et al. 1993), prostate (Nevalainen and Haapanen 1993), and lacrimal glands (Nevalainen et al. 1994). In the rat, smooth muscle cells have been shown to express group II PLA2 (Nakano and Arita 1990). Both neutrophil (Rosenthal et al. 1995) and mast cell (Murakami et al. 1992; Chock et al. 1994) secretory granules have been shown to contain group II PLA2.
Recently, we established transgenic mouse lines that express human group II PLA2 at serum levels that were 8- to 10-fold higher than serum levels in non-transgenic littermates (Grass et al. 1996). The current transgenic mice were produced in B6SJLF2 embryos and backcrossed to C57BL/6 mice. The latter strain lacks a functional group II PLA2 gene (Kennedy et al. 1995). The antiserum used in the current immunohis-tochemical tests was produced in a rabbit immunized by recombinant human group II PLA2 (Nevalainen et al. 1993). This antiserum was found to crossreact with the murine PLA2 found in the Paneth cells of the SJL mice. Because we saw no immunoreaction in the Paneth cells of C57BL/6 or the transgenic and non-transgenic mice used in this study, we are certain that the transgenic mice used in the current study lack the functional endogenous group II PLA2 gene. In addition, we have compared the catalytic activity of PLA2 in serum of our transgenic and non-transgenic mice with that of non-transgenic C3H/He and Balb/c mice (Grass et al. 1996). Both C3H/He and Balb/c mice have been shown to possess a functional group II PLA2 allele (Kennedy et al. 1995). Therefore, this experiment served to compare the level of PLA2 activity in our mice with mice containing a completely functional endogenous group II PLA2 allele. There were similar levels of serum PLA2 activity in our non-transgenic mice (on a C57BL/6 background) and in both C3H/He and Balb/c mice. Therefore, the 8- to 10-fold difference in serum PLA2 activity between our goup II PLA2 transgenic mice and their non-transgenic littermates was not an overestimate due to the absence of a functional endogenous group II PLA2 allele. In addition, these results suggest that little of the detectable serum PLA2 activity in both C3H/He and Balb/c mice is due to group II PLA2.
In this study we investigated the distribution of human group II PLA2 and the expression of the human group II PLA2 gene in tissues of transgenic mice. By immunohistochemistry, the enzyme protein was localized in hepatocytes, Bowman's capsule of the kidney glomeruli, cytoplasmic granules of the proximal tubule cells of the kidney, and epithelial cells of the renal pelvis, ureter, and urinary bladder. By in situ hybridization, clear signals for group II PLA2 mRNA were found in hepatocytes, Bowman's capsule, epithelial and smooth muscle cells of the urinary bladder, smooth muscle cells of the small intestinal wall, epithelial cells of the epidermis, and dermal and connective tissue cells, including fibroblasts. However, there was no clear signal in kidney tubules, aorta, or cartilage by in situ hybridization. These results suggest that group II PLA2, although present in these structures, is not synthesized by these tissues but is derived from elsewhere, e.g., blood plasma. Alternatively, the expression levels in the kidney tubules, aorta, and cartilage may be below the level of detection of the current method of in situ hybridization.
A 6.2-
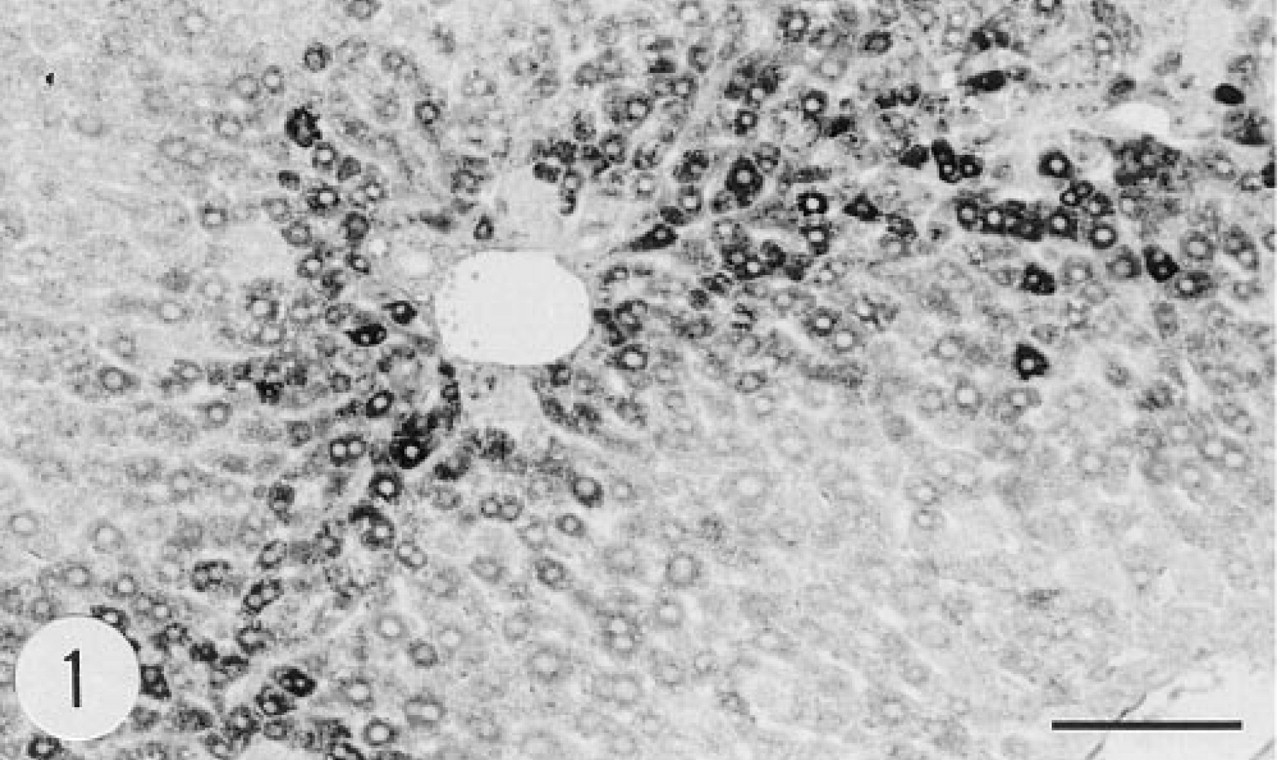
Immunoreaction for human group II PLA2 in the cytoplasm of hepatocytes of a transgenic mouse, female, age 4.5 months. Anti-group II PLA2, avidin-biotin-peroxidase complex (ABC). Bar = 100 μm.
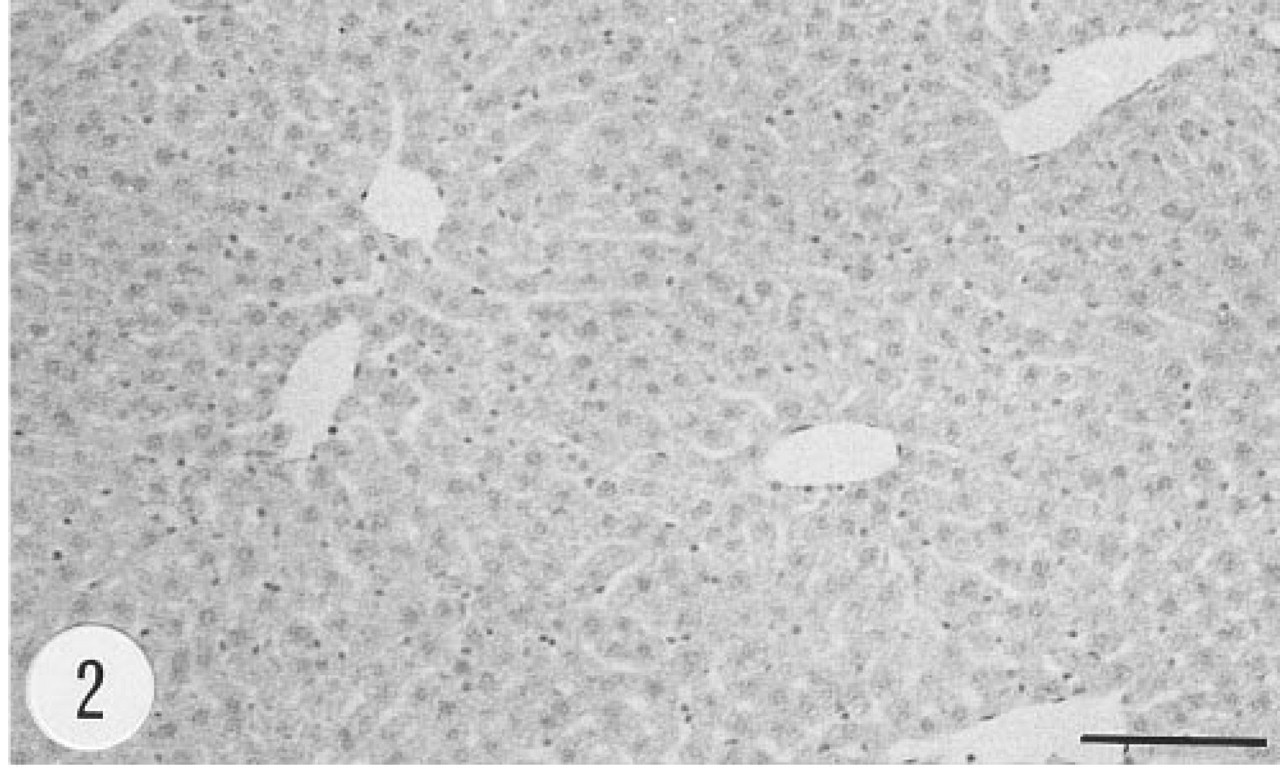
Absence of immunoreaction for human group II PLA2 from the liver of a non-transgenic control mouse, female, 4.5 months. Anti-group II PLA2 antibody, ABC. Bar = 100 μm.
In our earlier study, human group II PLA2 mRNA was found by Northern blotting in samples of the kidney of transgenic mice (Grass et al. 1996). The current in situ hybridization results show localization of the mRNA of human group II PLA2 in Bowman's capsule of the glomerulus, whereas proximal tubules contained immunoreactive granules in the apical cytoplasm but no in situ hybridization signal. It can be assumed that human group II PLA2 circulating in the blood plasma of transgenic mice may enter primary urine through the glomerulus in the same way as other small molecular weight proteins. The proximal tubule cells then absorb PLA2 from the primary urine, and accumulate and metabolize the protein in lysosomes. Our current in situ hybridization results support the idea of tubule absorption of group II PLA2 from primary urine as opposed to local production in the proximal tubule cells. Epithelial cells of the renal pelvis, ureter, and urinary bladder were strongly immunoreactive for group II PLA2, and a clear signal for the mRNA of human group II PLA2 was detected by in situ hybridization in the latter cells as well as in smooth muscle cells of the urinary bladder wall.
Articular chondrocytes and the matrix of cartilage of the transgenic animals contained immunoreactive human group II PLA2. However, we were unable to demonstrate unequivocally the presence of group II PLA2 mRNA in cartilage in the current transgenic animals by in situ hybridization. Human chondrocytes synthesize group II PLA2 and are believed to be a source of this enzyme found in synovial fluid in various arthropathies (Nevalainen et al. 1993; Kortekangas et al. 1994). The role of group II PLA2 in cartilage and bone is largely unknown. It has been proposed that the expression of group II PLA2 is connected to bone formation (Ellies et al. 1992). In general, other roles than the release of arachidonic acid from membrane phospholipids are less well established for PLA2. An antibacterial effect has been proposed for PLA2 (Wright et al. 1990; Harwig et al. 1995; Weinrauch et al. 1996).
The intimal and medial layers of the aorta of the transgenic mice contained immunoreactive human group II PLA2. Rat aortic smooth muscle cells express group II PLA2 when stimulated by inflammatory mediators, including cytokines and endotoxin, both in vivo (Nakano and Arita 1990) and in vitro (Nakano et al. 1990). Endotoxin and cytokines induce the expression of group II PLA2 also in a number of other cell types, including human choriodecidua (Nguyen et al. 1994), cultured rat astrocytes (Oka and Arita 1991), and human umbilical vein endothelial cells (Murakami et al. 1993). In the human, group II PLA2 has been localized by immunohistochemistry in both smooth muscle cells (Vadas et al. 1993; Andersen et al. 1994) and elastic fibers (Nevalainen et al. 1995) of blood vessels. In the latter study, group II PLA2 mRNA was not detected in vascular structures by in situ hybridization, and therefore it was postulated that the immunoreactivity seen in vessel walls may represent PLA2 absorbed from blood plasma rather than produced locally in the vessel wall (Nevalainen et al. 1995). In the current study, in situ hybridization for human group II PLA2 mRNA gave negative results in the aorta. Therefore, it is feasible to postulate that absorption from blood plasma may explain the presence of human group II PLA2 in the aortic wall of transgenic mice.

Positive in situ hybridization signal for the mRNA of human group II PLA2 in the liver of a transgenic mouse, female, 4.5 months. Anti-sense riboprobe. Bar = 50 μm.

Negative in situ hybridization signal for the mRNA of human group II PLA2 in the liver of a transgenic mouse, female, 4.5 months. Control (sense) riboprobe. Bar = 50 μm.
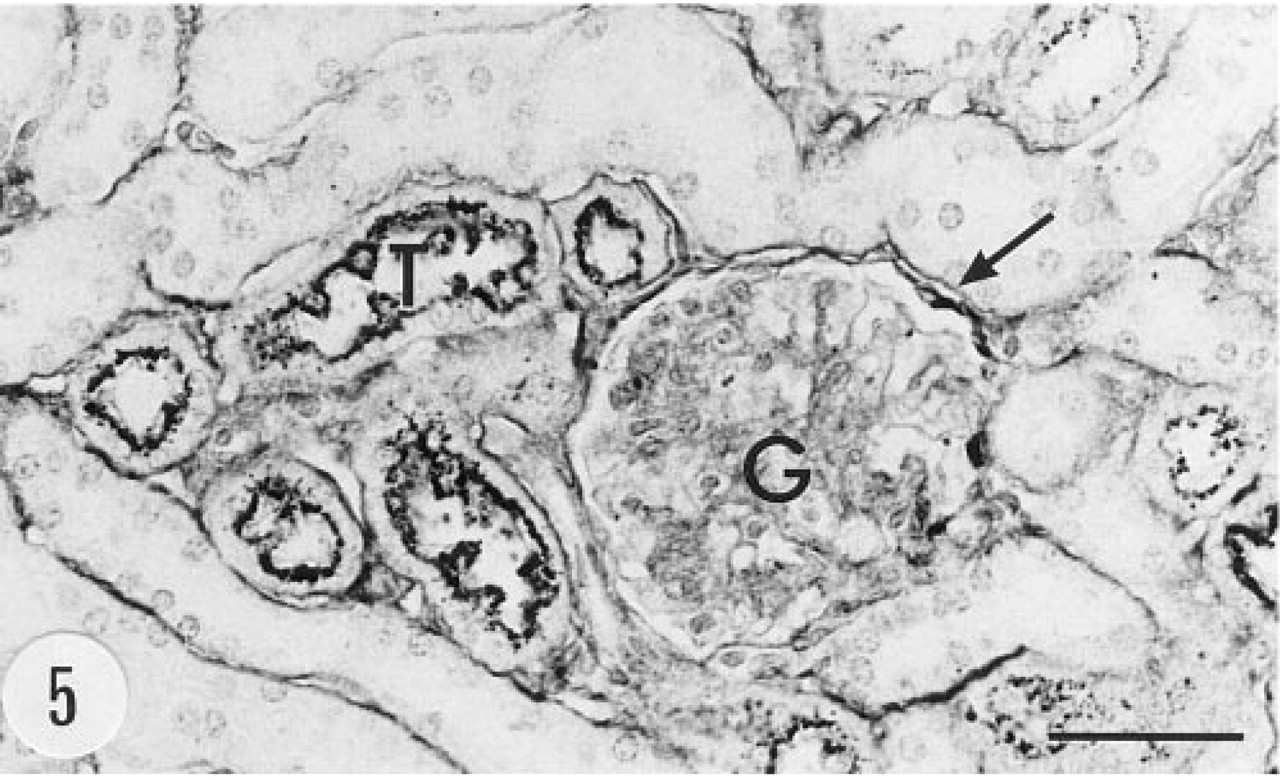
Immunoreaction for human group II PLA2 in the kidney. Bowman's capsule (arrow) and proximal tubules (T) show immunoreaction. Male, 6.5 months, anti-group II PLA2 antibody, ABC. G, glomerulus. Bar = 50 μm.
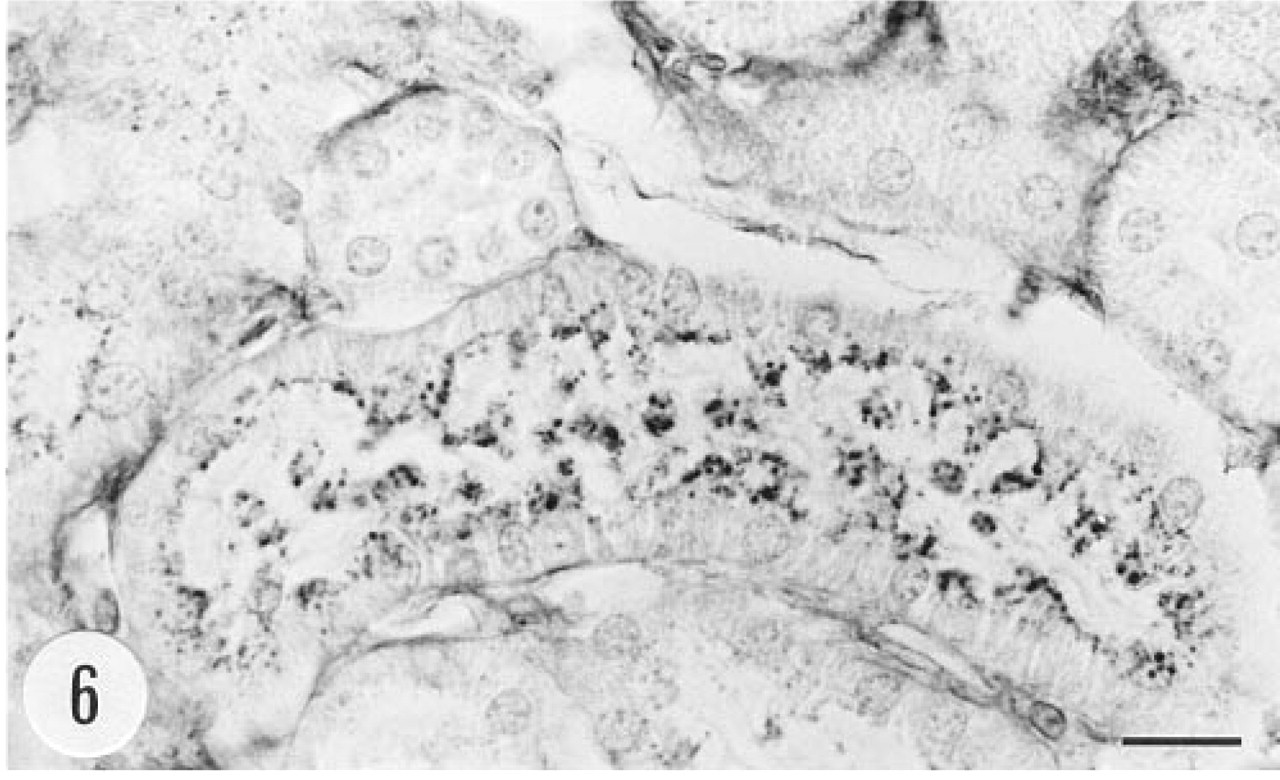
Small granules (most probably Iysosomes) in the apical cytoplasm of proximal tubule cells of a transgenic mouse show strong immunoreaction for human group II PLA2. Male, 6.5 months, anti-group II PLA2 antibody, ABC. Bar = 10 μm.

Positive in situ hybridization signal for the mRNA of human group II PLA2 in Bowman's capsule of a glomerulus in a transgenic mouse, female, 4.5 months. Note the absence of the signal from the surrounding proximal tubules (compare with Figure 8). Anti-sense riboprobe, darkfield. Bar = 50 μm.
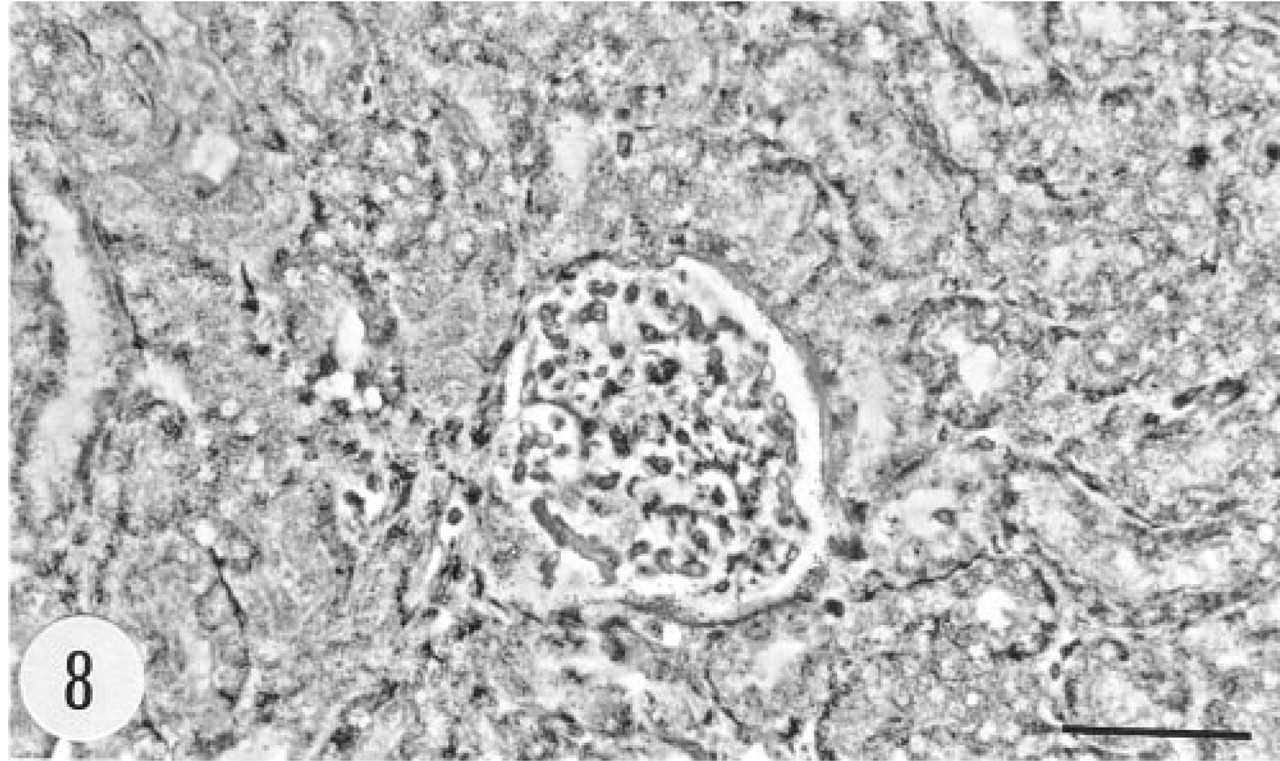
Phase-contrast view of the section showing the glomerulus and the surrounding proximal tubules in Figure 7. Anti-sense riboprobe. Bar = 50 μm.
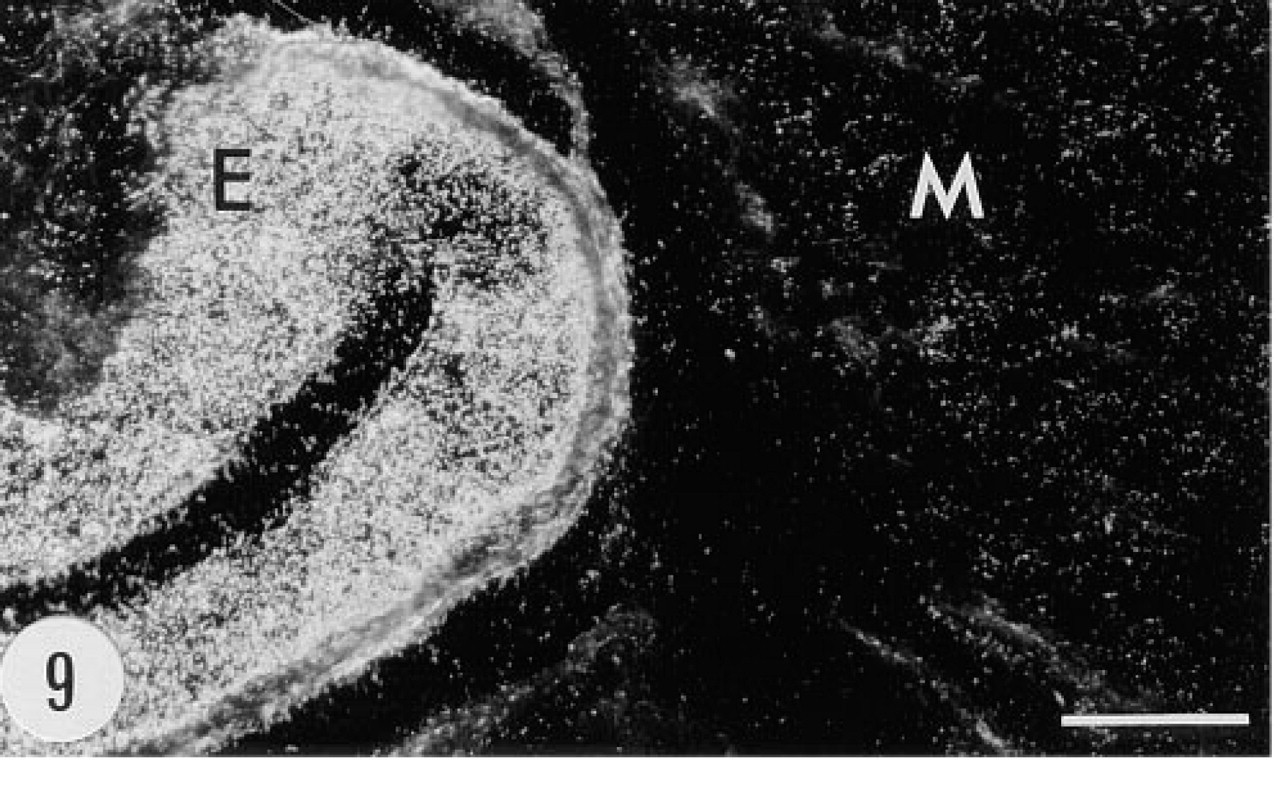
Positive in situ hybridization signal for the mRNA of group II PLA2 in the epithelium (E) and smooth muscle cell layer (M) of the urinary bladder of a transgenic mouse, male, 6.5 months. Anti-sense riboprobe, darkfield. Bar = 50 μm.
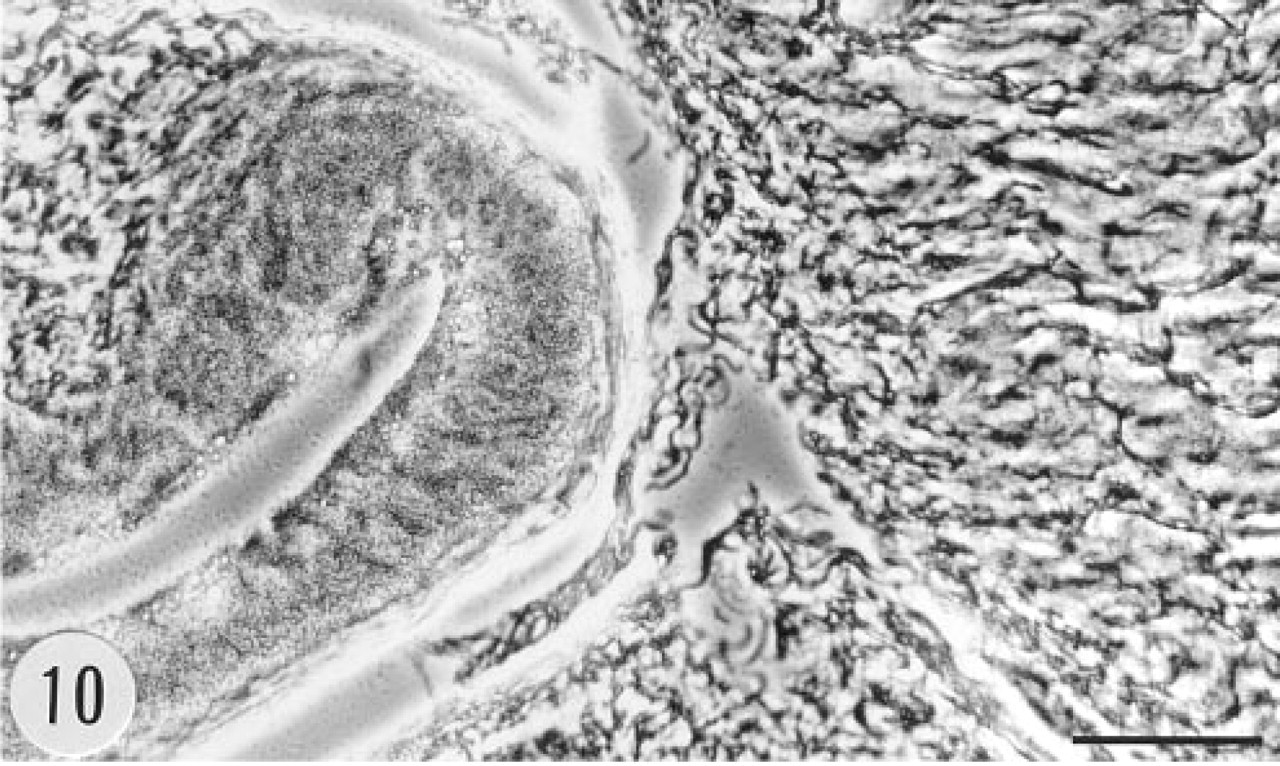
Phase-contrast view of the section showing the epithelium (left) and the muscular wall (right) of the urinary bladder in Figure 9. Anti-sense riboprobe. Bar = 50 μm.
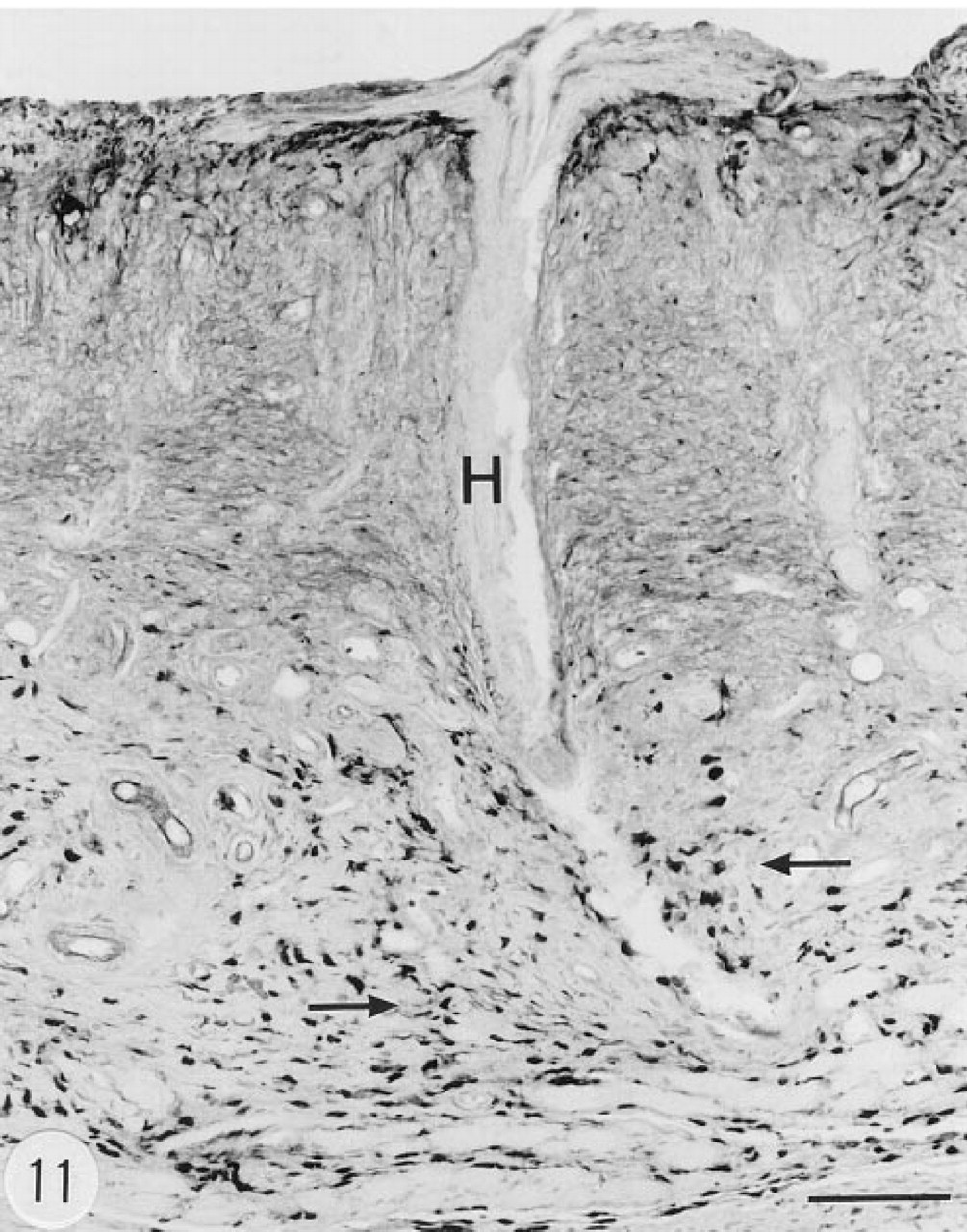
Immunoreaction for human group II PLA2 in the skin of a transgenic mouse. There is marked hyperkeratosis and a follicle with a deformed hair (H), and many small, elongated, strongly immunoreactive cells in the dermis (arrows). There are some immunoreactive cells also in the upper layers of the epidermis. Male, 6.5 months. Anti-group II PLA2 antibody, ABC. Bar = 100 μm.
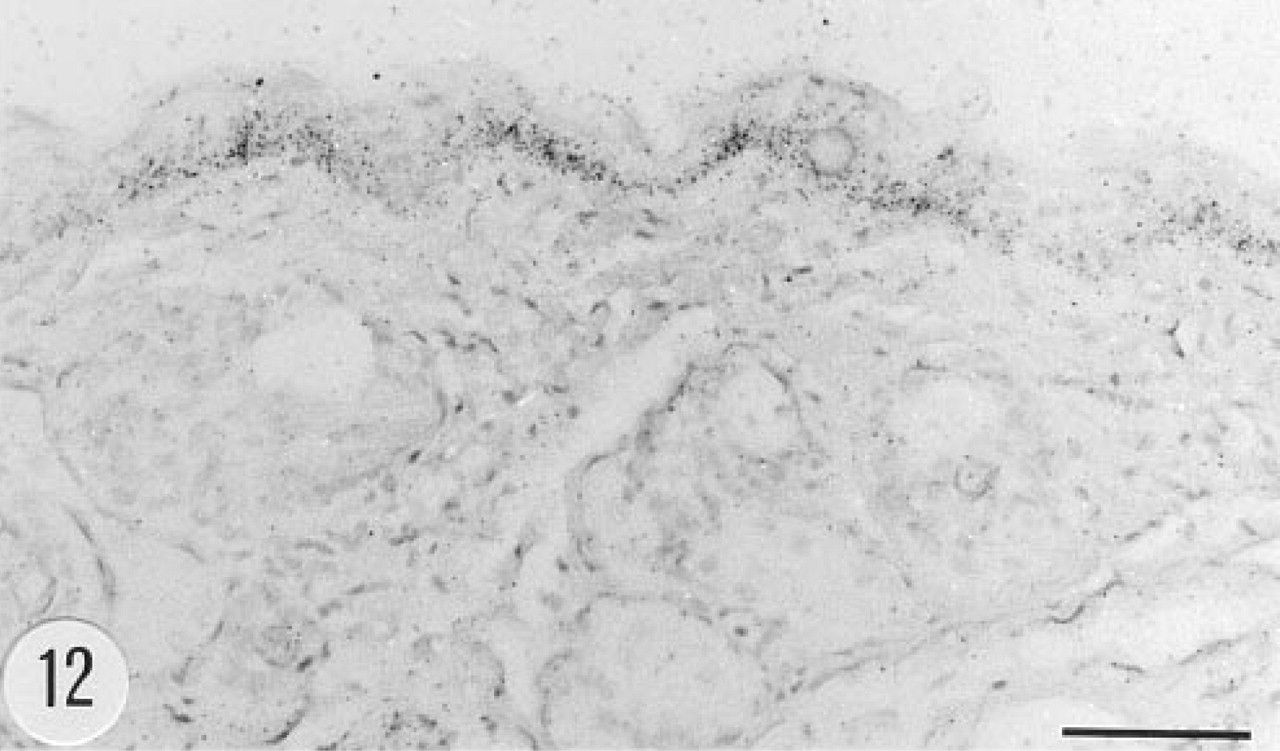
Positive in situ hybridization signal for human group II PLA2 mRNA in the epidermis of a transgenic mouse, female, 4.5 months. Anti-sense riboprobe. Bar = 50 μm.
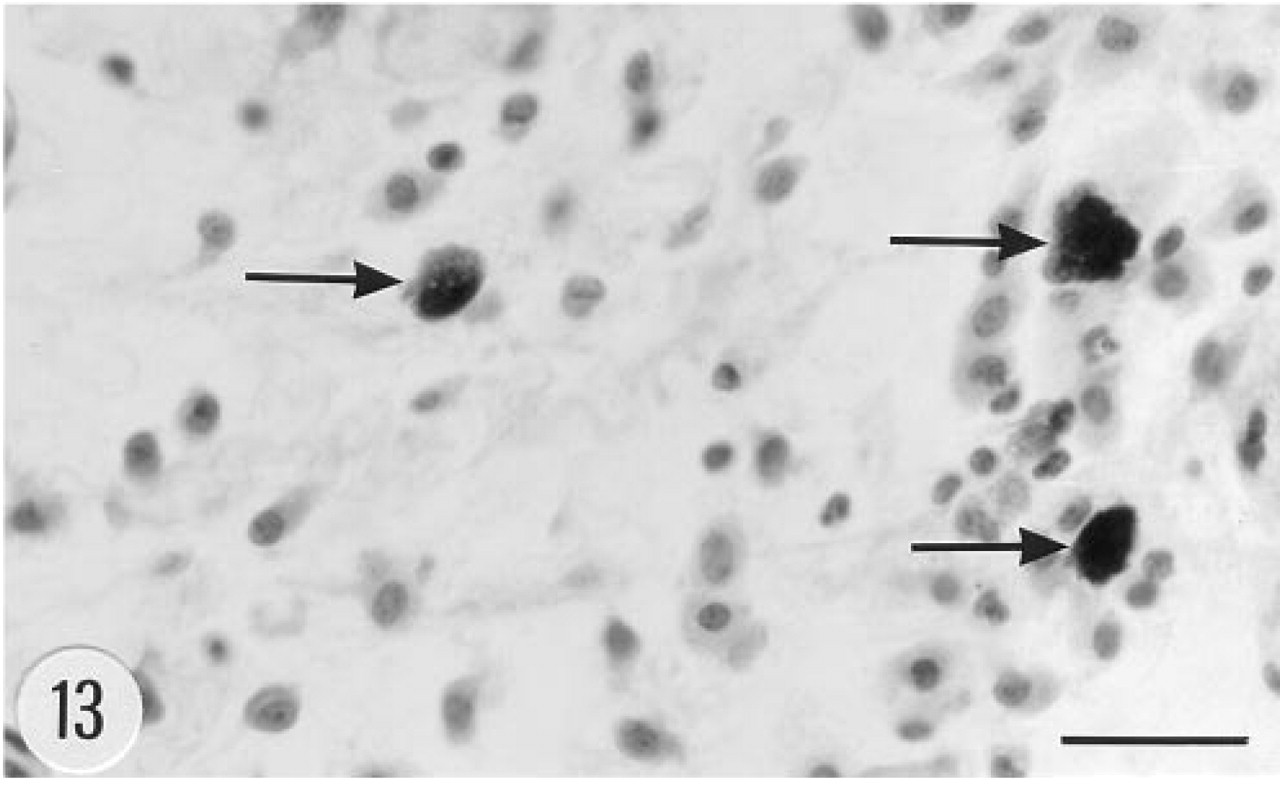
Fibroblasts, histiocytes, and mast cells (arrows) in the dermis of a transgenic mouse, female, 4.5 months. Hematoxylin–eosin. Bar = 25 μm.
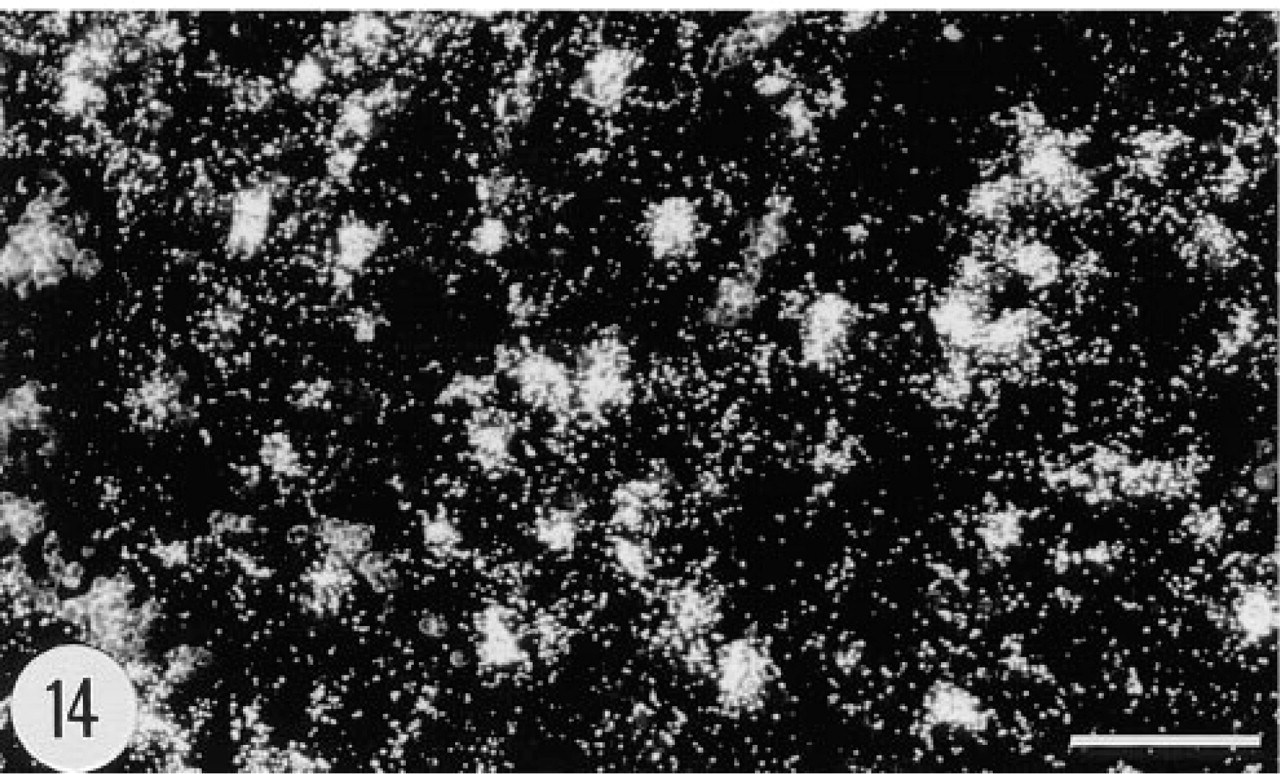
Positive in situ hybridization signal for the mRNA of human group II PLA2 in dermal cells in the skin of a transgenic mouse, female, 4.5 months. Anti-sense riboprobe, darkfield. Bar = 50 μm.
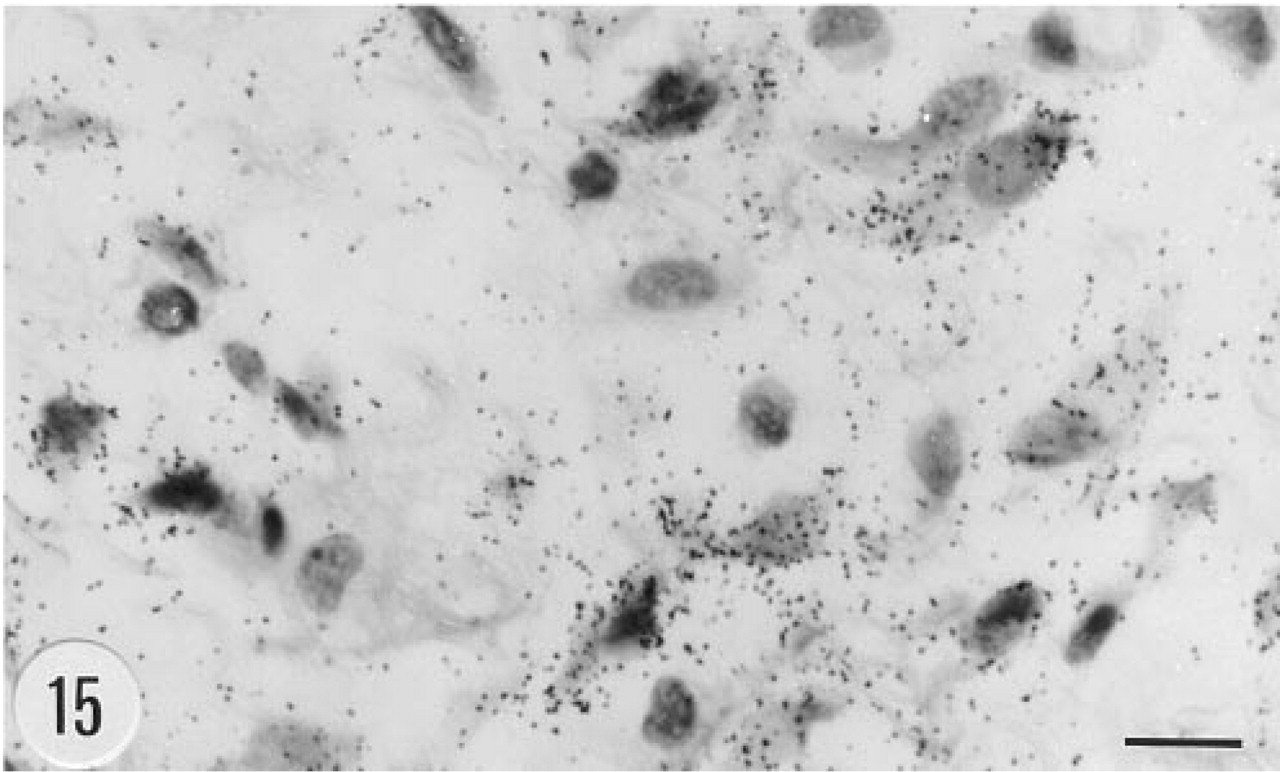
Positive in situ hybridization signal for the mRNA of human group II PLA2 in fibroblats in subcutaneous loose connective tissue from the neck of a transgenic mouse, male, 6.5. months. Anti-sense riboprobe, darkfield. Bar = 10 μm.
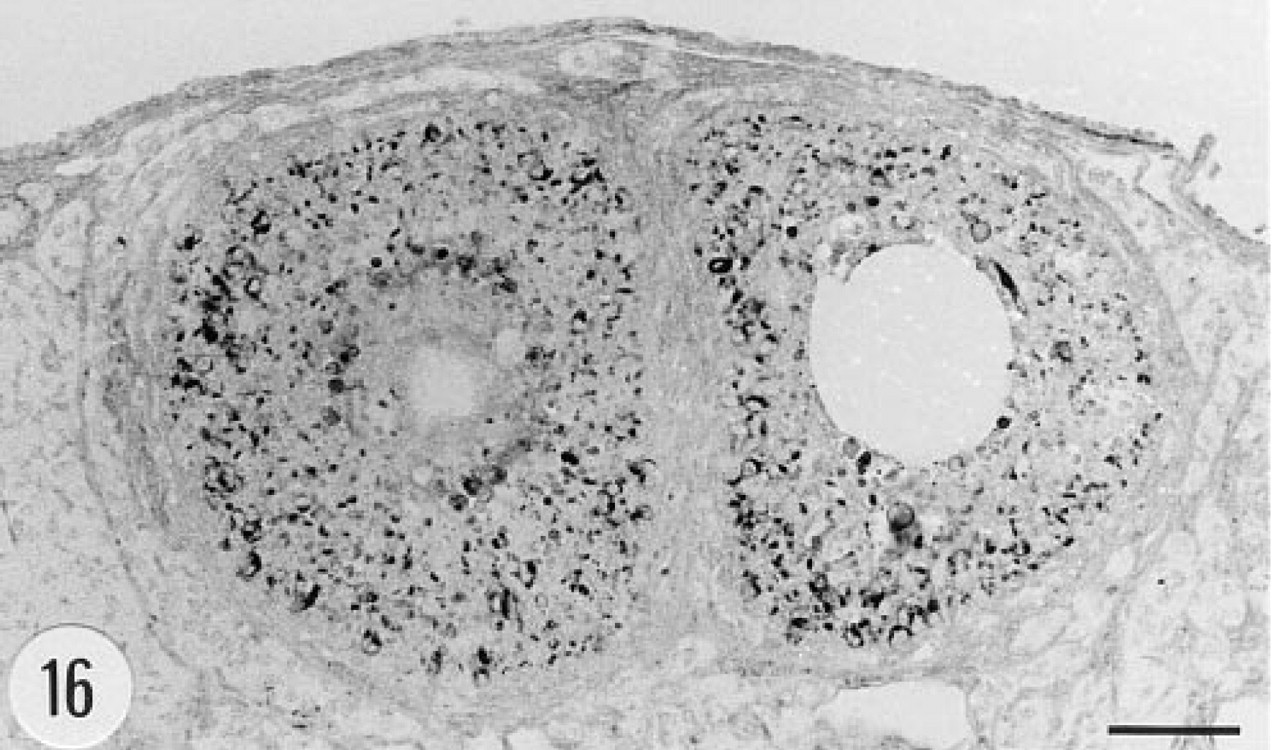
Immunoreaction for human group II PLA2 in the ovary of a transgenic mouse. The granulosa cells of the Graafian follicles contain immunoreactive material. Female, 4.5 months. Anti-group II PLA2 antibody, ABC. Bar = 50 μm.
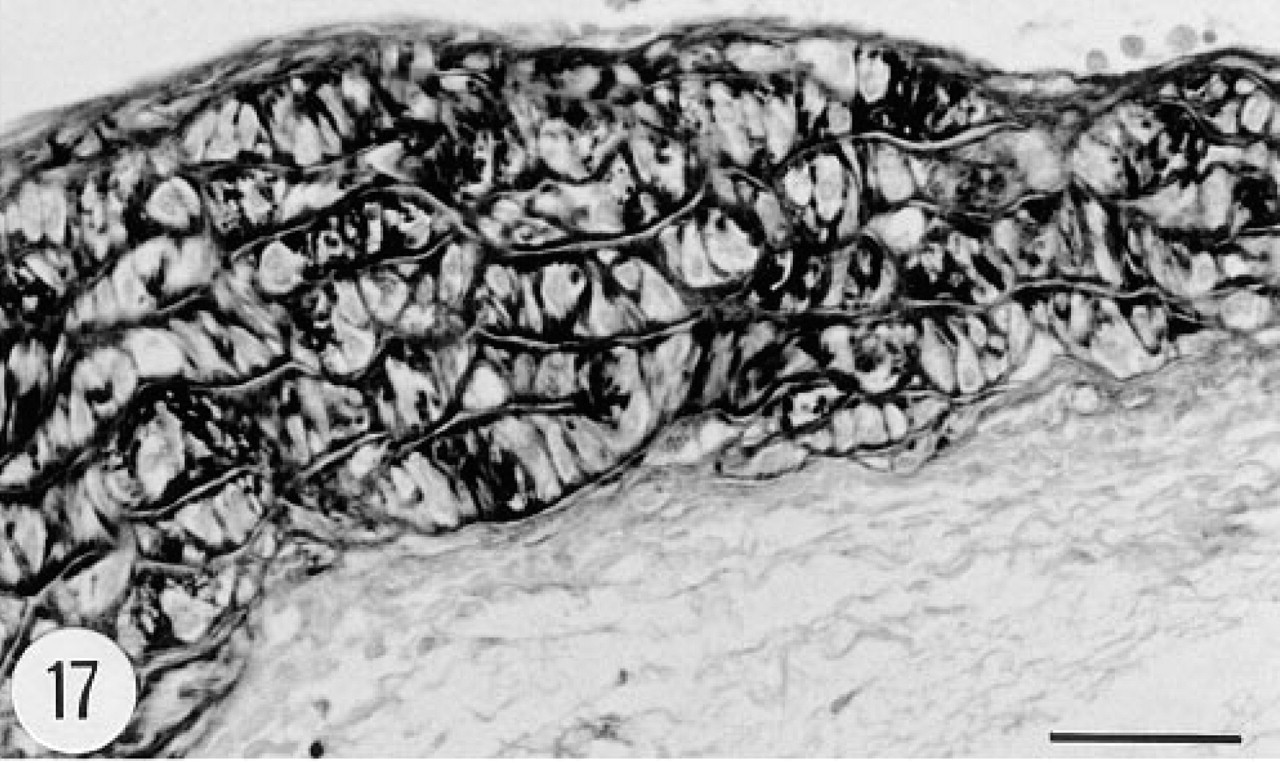
Immunoreaction for human group II PLA2 in the aorta of a transgenic mouse. Both the intima (top) and the media are strongly stained, whereas the adventitia (bottom) is devoid of immunoreaction. Male, 6.5 months. Anti-group II PLA2 antibody, ABC. Bar = 50 μm.
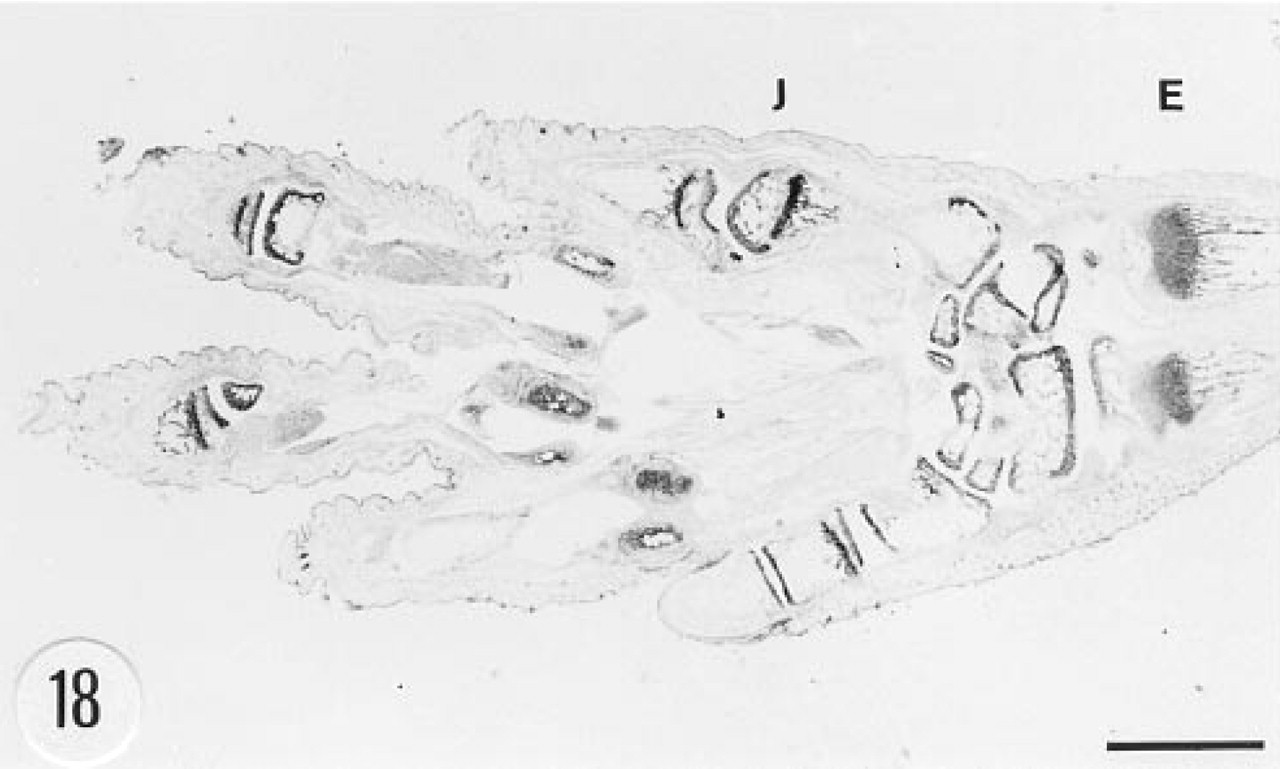
Immunoreaction for human group II PLA2 in the foot of a transgenic mouse. There is a strong immunostaining in the joints (J) and epiphyseal bone (E). Male, 3 weeks. Anti-group II PLA2 antibody, ABC. Bar = 1 mm.
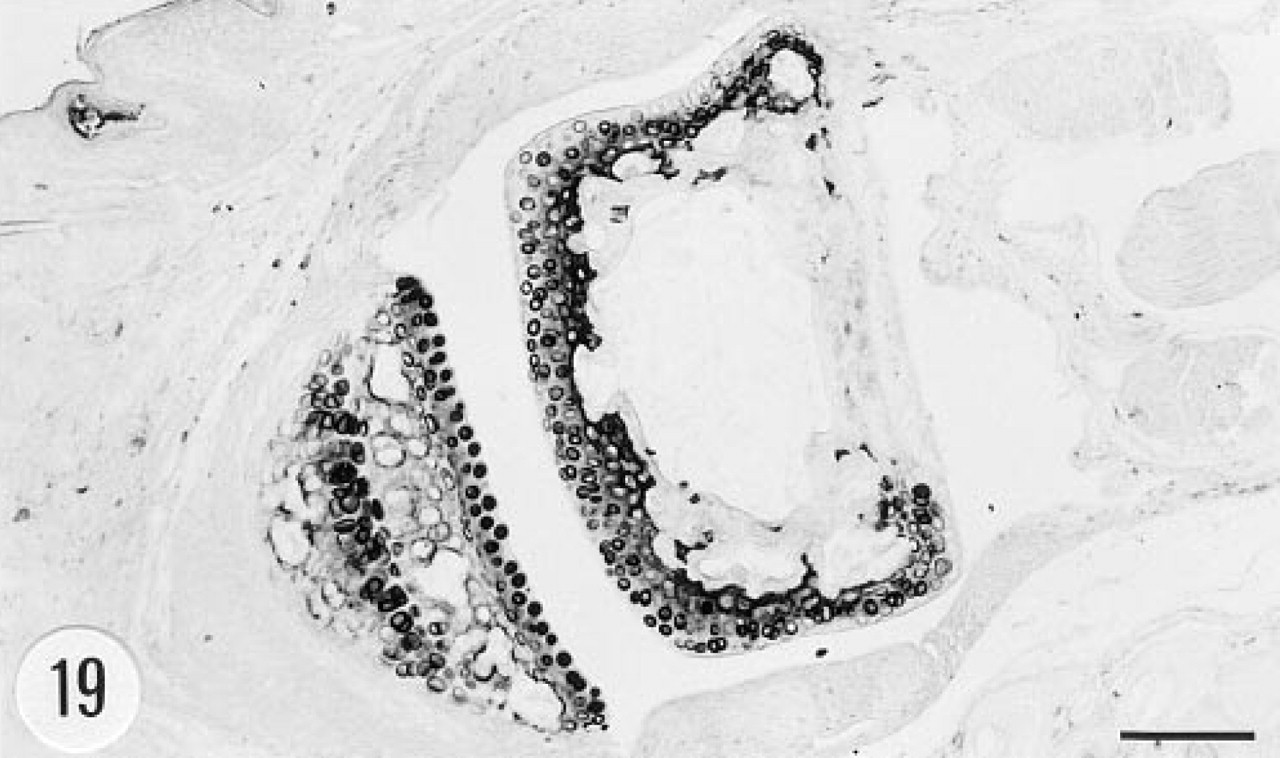
Immunoreaction for human group II PLA2 in the foot joint of a transgenic mouse. Articular chondrocytes are strongly immunoreactive. Male, 6.5 months. Anti-group II PLA2 antibody, ABC. Bar = 100 μm.
Transgenic mice expressing the human group II PLA2 gene have severe abnormalities in their skin. The animals lose hair and have marked epidermal and adnexal hyperplasia and hyperkeratosis (Grass et al. 1996). In the current study, human group II PLA2 was found by immunohistochemistry in elongated cells in the dermis. These cells most probably represent connective tissue cells. Dermal connective tissue cells and fibroblasts of the loose subcutaneous connective tissue contained the mRNA of human group II PLA2. In addition to the dermis, a few cells in the basal layer of the epidermis contained immunoreactive material. In situ hybridization gave positive signals for the mRNA of group II PLA2 also in epidermal epithelial cells. The expression of group II PLA2 is enhanced in the skin of patients with psoriasis (Andersen et al. 1994). Similar epidermal abnormalities, including hyperplasia and hyperkeratosis as in the current transgenic mice, are seen in psoriatic human skin. However, infiltrating neutrophils that form the epidermal microabscesses characteristic of psoriatic skin were not found in the transgenic mice. The mechanism by which the overexpression of human group II PLA2 in epidermal and dermal cells of transgenic mice and psoriatic human patients may result in abnormal skin phenotype remains to be resolved.
Granulosa cells of the Graafian follicles of the ovary contained immunoreactive human group II PLA2, whereas immunostaining gave negative results in the testis. PLA2 activity has been detected in ovaries (Okazaki et al. 1977), and the enzyme has been shown to regulate prostaglandin synthesis in these cells (Veldhuis and Demers 1989). Interestingly, pancreatic group I PLA2 was recently found by immunohistochemistry in the granulosa cells of rat Graafian follicles (Nomura et al. 1994).
An interesting negative finding in the current study was that human group II PLA2 was not detected in the Paneth cells of the small intestinal mucosa of the transgenic mice. These cells are known to synthesize group II PLA2 in the human (Nevalainen et al. 1995). Another notable negative finding was the absence of human group II PLA2 from the bone marrow, lymph node, and spleen cells of the transgenic mice. Group II PLA2 is also not present in these cells in the human (unpublished results; Nevalainen and Haapanen 1993).
As indicated above, human group II PLA2 was expressed in many tissues of transgenic mice, and the catalytic activity of PLA2 in sera of these animals was markedly increased over that of the control mice. Despite the high levels of circulating PLA2 activity and expression of group II PLA2 in various vital organs, the animals showed no signs of generalized inflammation and were capable of reproducing (Grass et al. 1996). These observations corroborate the notion that group II PLA2 is relatively non-toxic to intact tissues and cells (Martikainen et al. 1993; Morgan et al. 1993).
In conclusion, the human group II PLA2 gene is expressed in various organs of transgenic mice. The enzyme is synthesized in hepatocytes, epidermis, dermis, fibroblasts of subcutaneous connective tissue, and epithelial and smooth muscle cells of the urinary bladder of these animals. The high local and circulating levels of group II PLA2 are not associated with compromised health of these animals, except for skin and fur abnormalities. Transgenic mice provide a useful tool with which to study the physiological and pathological roles and the regulation of the synthesis and metabolism of group II PLA2.
Footnotes
Acknowledgements
Supported by Turku University Foundation, by the Academy of Finland, and by the Cancer Foundation of Southwestern Finland.
We thank Ms Tuula Manninen, Ms Outi Asunta, and Mr Jaakko Liippo for technical assistance, Ms Maija Ahlholm for secretarial help, Drs Markku Kallajoki and Kai Nyman for methodologic advice, and Dr Mark Swanson for critical reading of the manuscript.