
Abstract
A number of cDNAs (SC1, QR1, and hevin) have been shown to be similar to SPARC (secreted protein acidic and rich in cysteine), a matricellular protein that regulates cell adhesion, cell cycle, and matrix assembly and remodeling. These proteins are 61–65% identical in the final 200 residues of their C-termini; their N-terminal sequences are related but more divergent. All have an overall acidic pl, with a follistatin-like region that is rich in cysteine, and a Ca+2 binding consensus sequence at the C-terminus. Using degenerate primers representing the most highly conserved region in SPARC, SC1, and QR1, we identified a 300-
Keywords
SPARC (
SPARC is a member of a family of proteins that exhibit a similar basic structure. These proteins have clusters of acidic residues imparting an overall acidic pI, a follistatin-like region that is cysteine-rich, and a high-affinity Ca2+ binding region termed an EF hand (Lane and Sage 1994). Members of this family include follistatin-like protein (TSC-36), albondin, and testican (Alliel et al. 1993; Shibanuma et al. 1993) as well as QR1, an extracellular matrix protein confined to postmitotic neural retina cells (Casado et al. 1996; Guermah et al. 1991), and SC1, a protein expressed widely by embryonic and adult neurons in the central nervous system (Johnston et al. 1990). The SC1 expressed by Bergmann glial cells is deposited along the length of their radial fibers during the process of granule cell migration in the developing cerebellum (Mendis et al. 1994). Subcellular fractionation demonstrated that the SC1 116/120-kD doublet is associated with synaptosomes (Mendis et al. 1996). Hevin, the human homologue of SC1 which has been isolated from high endothelial venules found in tonsils (Girard and Springer 1995), inhibits attachment and spreading of endothelial cells on fibronectin substrates (Girard and Springer 1996). In addition, hevin-treated cells exhibit a rounded morphology and do not form focal adhesions (Girard and Springer 1996). SPARC has similarly been shown to inhibit cell spreading on collagen and to induce cell rounding in cultured endothelial cells and fibroblasts (Sage et al. 1989).
In this study we used polymerase chain reaction (PCR) methods, in addition to library screening, to search for other SPARC family members. We describe the cloning of SC1 from a mouse whole-brain cDNA library. We subsequently used ribonuclease protection and in situ hybridization assays to examine the distribution of mouse SC1 mRNA in various organs and during development. Our results indicate that the primary site of expression of SC1 mRNA is the brain, although lower levels of transcript were noted in several other organs/tissues. In the vasculature, SC1 was associated with the adventitia and/or media of medium and large vessels in all organs examined. Because SPARC and hevin perform overlapping functions in vitro, it is possible that these proteins compensate for each other during development or response to injury.
Materials and Methods
Isolation of SC1 cDNA from Mouse Whole-brain Library
Degenerate primers designed according to the C-terminal regions of SPARC, QR1, and rat SC1 (Figure 1) were used in a primary screen of a mouse whole-brain cDNA library (gift from Jim Boulter, Salk Institute) by the MOPAC procedure as described by Lee and Caskey (1990). The primers were as follows: (SF = SPARC family) SF.forward (SF.for) 5′-atg(ca) g(agct)ga(tc)tgg(ct)(agct)aa-3'; SF.reverse-2 (SF.rev-2) 5′-(ga) ca(ga)tg(ac)tccat(agct)gg-3'; SF.reverse (SF-rev) 5′-(ag)aa(tc) tgcca(ag)tg(agct)ac(agct)gg-3'; and SF.internal (SF.int) 5′-aa (ag)aa(tc)ta(tc)(ac)a(tc)atgtat-3'. Primers were used at a concentration of 10.85 μM (SF.for) and 12 μM (SF.rev-2). The library was diluted 1:100 (original concentration of library was 5 × 107 pfu/ml), and 5 μl was used in the polymerase chain reaction (PCR). Taq polymerase was added after the samples were heated to 94C for 5 min. The reaction proceeded with an initial cycling at 1 min 94C, 1 min 50C, and 3 min 72C (three cycles), followed by 40 cycles at 1 min 90C, 1 min 55C, and 2 min 72C, with a final extension of 10 min at 72C. Reaction products were resolved on a 4% NuSieve gel (FMC BioProducts; Rockland, ME) and were transferred to Duralose-UV membrane (Stratagene; La Jolla, CA) via Southern blot. Blots were probed with SF.rev and SF.int oligonucleotides that had been 5′-labeled with [γ32P]-ATP (New England Nuclear; Boston, MA), 5′ exchange buffer, and polynucleotide kinase (Gibco BRL; Grand Island, NY). Bands that hybridized with the probes were isolated on 4% NuSieve gel and were excised from the gel. The agarose was subsequently removed with Agarase (Boehringer Mannheim Biochemicals; Indianapolis, IN). These cDNA clones were subcloned into a blunted Eco RV site in a Bluescript II SK (+/-) vector (Stratagene) and were transformed into competent XLI-Blue bacteria. Positive clones were selected and were cultured in Luria broth. Pure DNA was isolated by mini prep (Qiagen; Chatsworth, CA), and clones were sequenced in conjunction with T7 and T3 primers (Stratagene) according to the procedure of Sanger et al. (1977). Analysis of DNA sequences was carried out on a VAX computer with the Genetics Computing Group program of the Wisconsin Genetics Group (Devereux et al. 1984). The program BLASTX (NIH) was used to compare the amino acid sequences with all the sequences present in the National Center for Biotechnology Information nonredundant protein database.
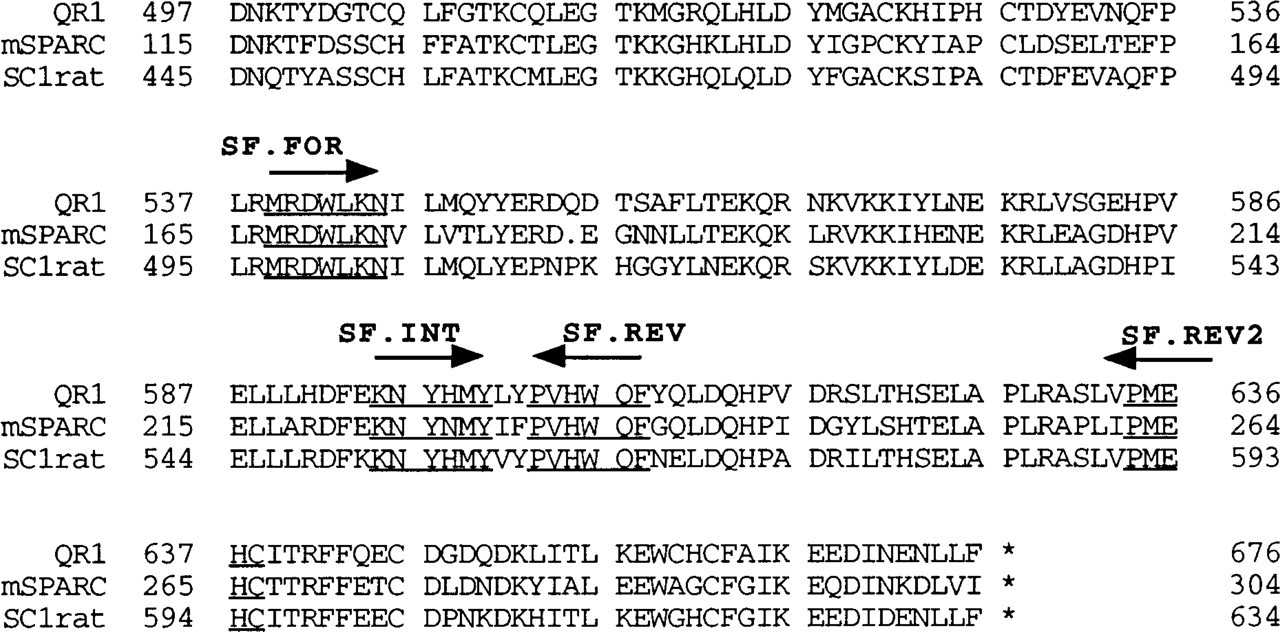
Alignment of C-termini of mouse SPARC, rat SC1, and quail QR1. Conserved regions allow the design of degenerate oligonucleotides for use in PCR library screens. Arrows indicate direction of oligonucleotide synthesis. Underlined sequences indicate sequence used for degenerate oligonucleotide synthesis.
Positive clones from the primary screen were used as probes to screen the cDNA library for full-length clones. The library was plated at 5000 pfu/ml. Probes were random-prime labeled (Amersham; Arlington Heights, IL) with [α32P]-dCTP (New England Nuclear). Duplicate filter lifts (Amersham) from 20 plates, each containing 50,000 plaques, were hybridized overnight at 42C in 3 × SSC [1 × SSC is 0.15 M NaCl, 0.015M Na-citrate (pH 7.5)], 0.5% SDS (sodium dodecyl sulfate), 25 mM Tris-HCl, 0.2% Ficoll, 0.2% polyvinylpyrrolidone (
5′ RACE (rapid amplification of cDNA ends) (Frohman et al. 1988) was utilized to obtain the N-terminal portion of the clone. Polyadenylated mRNA from mouse whole brain was isolated on an oligo-dT column (Stratagene). A genespecific primer was designed against part of the existing 5′ sequence (5′-ggtgtgagtcacttacaggttgc-3'), 240
RNA Extraction and Northern Analysis
Total RNA was isolated from mouse tissues (C57BL/6J) and cultured cells with Tri-Reagent (Molecular Research Center; Cincinnati, OH). Samples were resuspended in water treated with diethyl pyrocarbonate (DEPC) (Sigma; St Louis, MO), and the concentration of RNA was calculated by
Mouse SC1 expression was analyzed by Northern blotting. An 800-
Ribonuclease Protection Assays
An 800-
RNAse protection assays were performed as previously described (Liska et al. 1990), with slight modifications. The RNAse digestion buffer was optimized for these probes at 5 μg/ml RNAse A (Sigma) and 2 μg/ml RNAse T1 (Sigma). The 28S riboprobe was used to normalize the signal in the assays. RNAse protection assays were repeated with RNA isolated from three different mice. Autoradiographs were analyzed by densitometric scanning.
In Situ Hybridization
The mouse SC1 and SPARC riboprobes were transcribed with an RNA transcription kit (Promega) in the presence of [α35S]-dUTP (Amersham). The specific activity of the probes was approximately 3–4 × 107 cpm/μg. The riboprobes were transcribed from a 800-
The in situ hybridization protocol was a modification of a method described by Holland et al. (1987). Kidney and adrenal glands from two Sprague-Dawley rats were excised and fixed in 10% buffered formalin phosphate (Fisher Scientific; Santa Clara, CA) and were embedded in paraffin. Kidneys from two C57BL/J6 mice were treated similarly. Liver, lung, heart, thymus, spleen, testis, and epididymis were removed from three (C57BL/6 × DBA/2)F1 mice and were fixed and embedded as described above. Sections of rat brain were used as a positive control for the SC1 riboprobe. Sections were rinsed in PBS, dipped in water treated with DEPC, and exposed to 0.1 M triethanolamine (pH 8.0) with 0.05% v/v acetic anhydride for 10 min. After an additional rinse in PBS for 5 min, slides were dipped in DEPC-water, dehydrated in graded solutions of ethanol (70–100%), and air-dried. Sections were treated overnight at 50C in prehybridization solution [0.6 M NaCl, 0.12 M Tris-HCl (pH 8.0), 8 mM EDTA (ethylenediaminetetracetic acid), 0.02% Ficoll 400, 0.02% polyvinylpyrrolidone, 0.1% Pentex BSA (Sigma), 500 μg/ml salmon sperm DNA, 600 μg/ml yeast total RNA, and 50% deionized formamide]. The samples were hybridized at 50C overnight in prehybridization solution that contained 10% dextran sulfate and 0.1 μg/ml of the SPARC and SC1 anti-sense and sense riboprobes. Samples were washed for 30 min at 50C in formamide buffer (50% formamide, 5% 20 × SSC, 1% 1 M DTT in DEPC-water), for 30 min at room temperature in 0.5 × SSC, and 30 min at 37C in 3.5 × SSC with 40 μg/ml RNAse A (Sigma) and 50 μl RNAse T1 (Sigma). Slides were incubated sequentially in 3.5 × SSC for 10 min at room temperature and 0.1 × SSC for 120 min at 65C. Sections were dehydrated with ethanol (solutions of 50, 70, 95, and 100%) containing 0.3 M sodium acetate and were subsequently air-dried. We performed autoradiography with NTB 2 emulsion (Kodak; Rochester, NY) diluted to 60% with a solution of 0.3% Dreft detergent; slides were exposed for 7 days at 4C, developed in Kodak D-19 developer, and fixed in GBX fixer (Kodak). Sample were counterstained with toluidine blue, dehydrated with ethanol (solutions of 70–100%), cleared in xylene, and mounted with Permount. Slides were examined and photographed with bright- and darkfield microscopy.
Results
Identification and Cloning of Mouse SC1
Degenerate oligonucleotide primers designed against the conserved C-terminal regions of mouse SPARC, quail QR1, and rat SC1 were used in a PCR screen of a mouse whole-brain cDNA library. Figure 1 shows the location of these primers in the amino acid sequence of these proteins. Isolated clones were sequenced and were compared to SPARC, QR1, and SC1. Clones that showed some similarity (80% or greater at the nucleotide level) to any of the three proteins were used to re-screen the library to retrieve the full-length cDNA. One of the clones isolated in this manner was identified as mouse SC1. The clone was lacking about 800
An 800-

Mouse SC1 sequence. The full-length cDNA sequence is shown with its corresponding translated amino acid sequence, as well as N-terminal and C-terminal non-coding sequences. The numbers to the left of the sequence refer to the nucleotide number, and the numbers to the right of the sequence refer to the amino acid number. The underlined amino acid sequence (1–18) represents a potential signal peptide sequence.
Expression of Mouse SC1 and SPARC in Different Tissues
Ribonuclease protection assays were performed with riboprobes for the N-terminal regions of SPARC and mouse SC1 on total RNA isolated from brain, heart, kidney, adrenal gland, lung, liver, eye, submandibular gland, epididymis, and testis. The expression pattern for these two mRNAs was similar (Figures 4A and 4B). Figure 5 is a graphic representation of the data shown in Figure 4. Compared to SC1 mRNA seen in brain, expression in other tissues was 60–90% lower in abundance. To ascertain whether SPARC and SC1 mRNAs were expressed by the same cell types, we performed Northern blots on a variety of cultured endothelial cell types (rat heart endothelium, bovine fetal and adult aorta, mouse aorta, human omentum, human aortic, and bovine retinal endothelial cells), as well as on smooth muscle cells and fibroblasts from mouse, bovine, and human sources. Although SPARC transcripts were abundant in these cells, SC1 was not expressed at detectable levels by endothelial cells, smooth muscle cells, or fibroblasts (data not shown).

SC1 clones from different species vary most at their N-termini. Numbers at either side of the sequences refer to amino acid position. The underlined sequence in mouse (m)SC1 refers to the longer stretch of glutamic acid residues found in mSC1, in comparison to rat SC1 and human hevin.
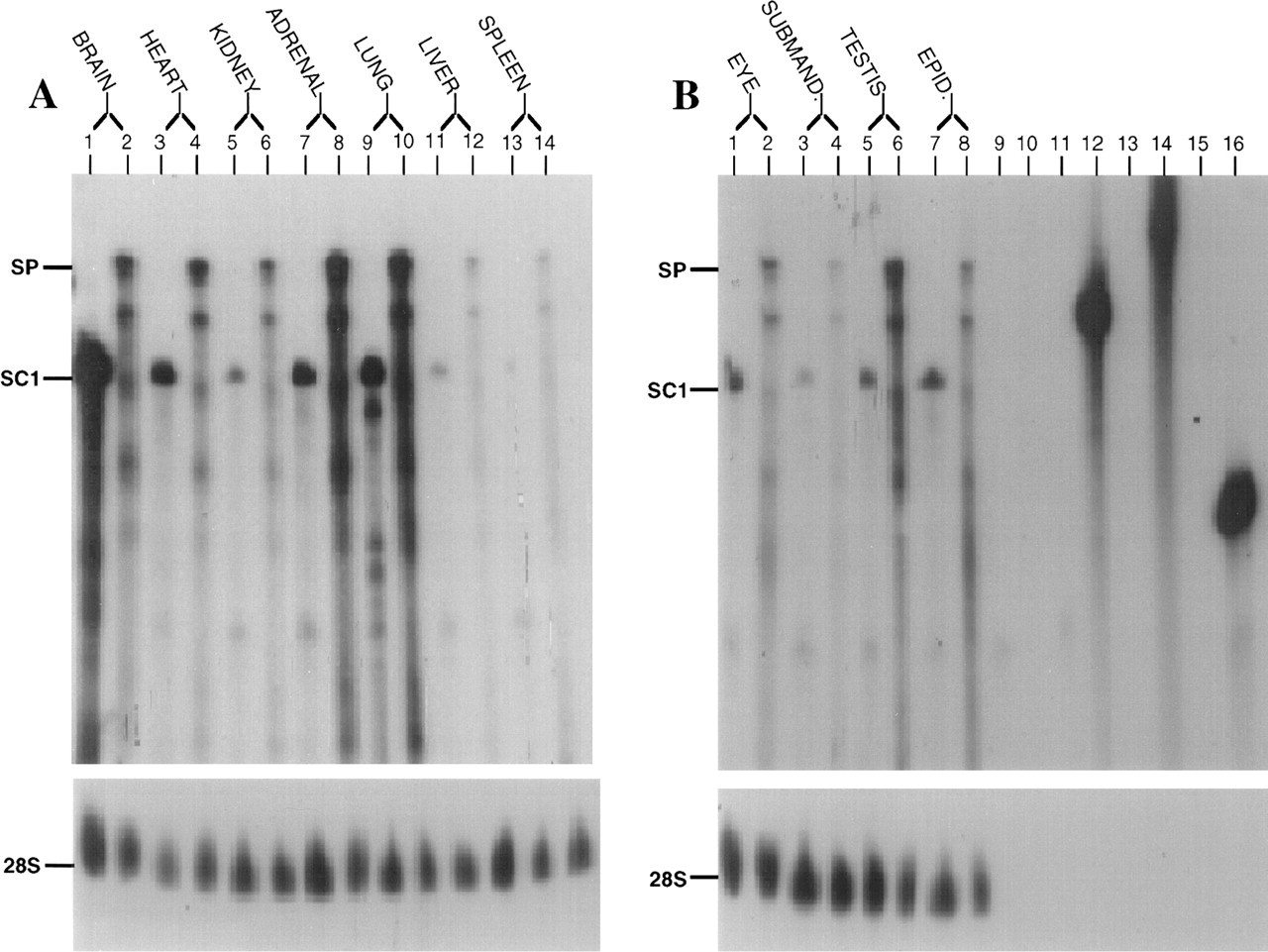
SC1 mRNA is expressed by a variety of murine tissues. (
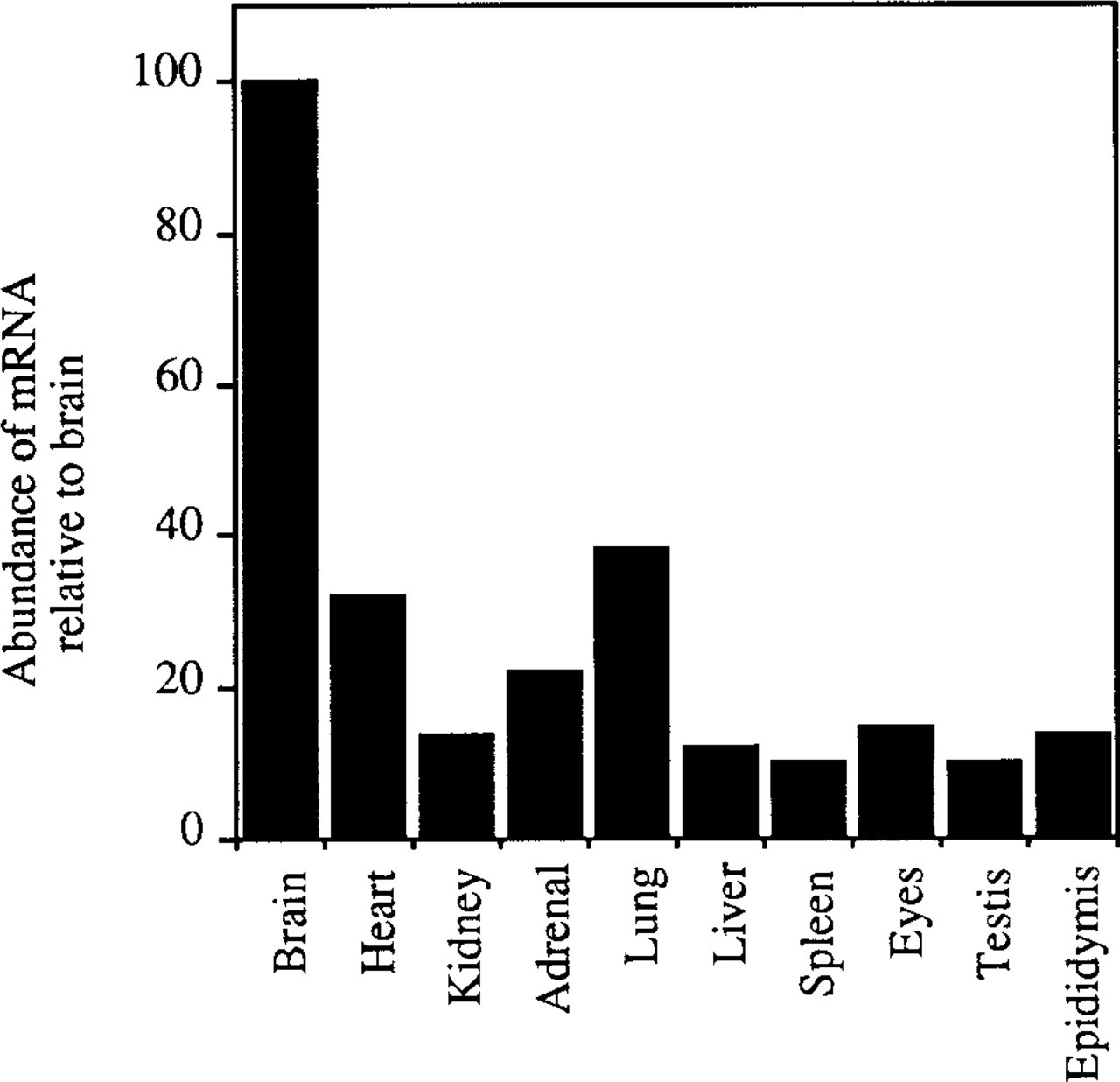
Murine tissues exhibit a moderate to low level of SC1 mRNA in comparison to that in the brain. Percentages were calculated based on densitometric scanning of the ribonuclease protection autoradiographs shown in Figure 4. The SC1 signal for each tissue was normalized to the corresponding 28S signal for that lane.
In Situ Hybridization Assays in Mouse Tissues
In situ hybridization was performed to determine the distribution of SC1 mRNA. Figure 6 shows in situ hybridization on rat kidney and adrenal gland and on mouse kidney. Figures 6A-6C represent tissues probed with anti-sense SPARC mRNA and Figures 6D-6F represent serial tissue sections probed with anti-sense SC1 mRNA. SPARC mRNA was found in the glomerular capillaries (Figures 6A and 6C) of the kidney, as well as in medium and large vessels. Expression of SPARC mRNA in the adrenal gland (Figure 6B) was predominant in the zona fasciculata of the adrenal cortex, with lesser amounts in the zona glomerulosa, zona reticularis, and medulla. Expression of SC1 mRNA in the kidney (Figures 6D and 6F) was also localized to the medium and large vessels of the kidney, whereas in the adrenal gland (Figure 6E), low levels of transcripts were detected throughout all zones of the cortex and in the medulla.
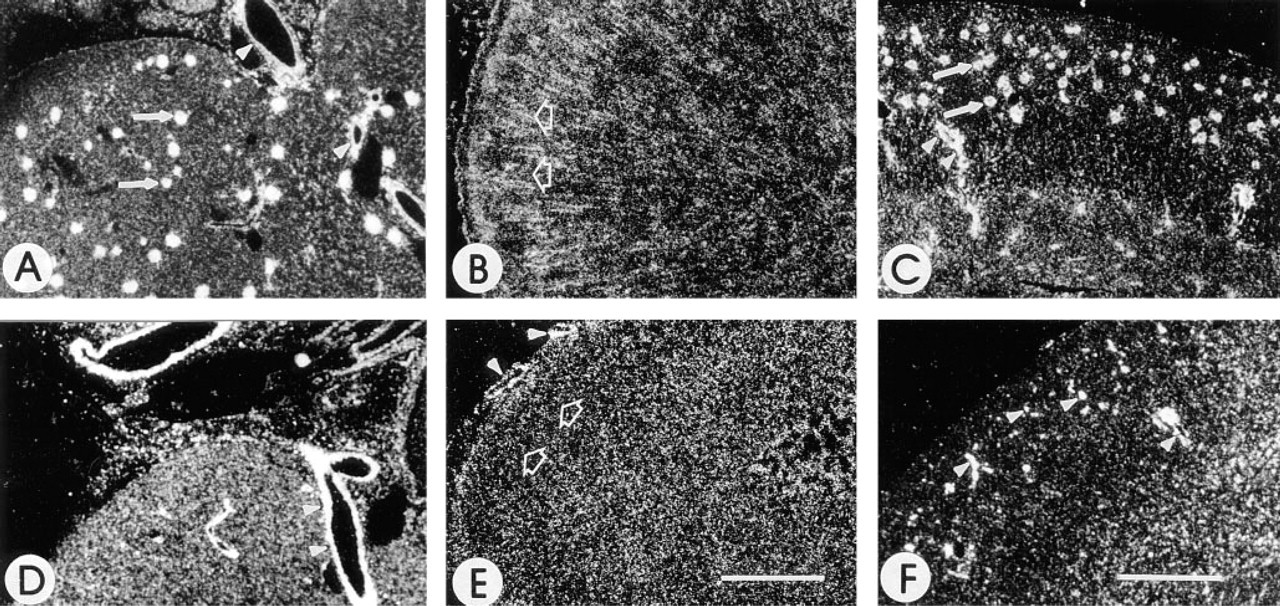
The distribution of SC1 is different from that of SPARC in the kidney and adrenal gland. Shown are darkfield photographs of rat kidney (
Figure 7 shows SC1 mRNA in heart, lung, liver, spleen, and epididymis. In the heart (Figure 7A), transcripts are noted in medium and larger-sized vessels coursing through the cardiac muscle and in a localized area of the apex consistent with terminal fibers of the conducting tissue of the heart. SC1 mRNA in the lung (Figure 7B) was detected in medium and large vessels and in the bronchi and bronchioles. The liver (Figure 7C) showed expression of SC1 mRNA almost exclusively in medium and larger vessels. Transcripts were not detected in the sinuses of the liver (Figure 7C) or the spleen (Figure 7D). In the spleen, SC1 mRNA was scattered throughout the red pulp. Transcripts were absent in the white pulp except for the expression of SC1 mRNA in medium-sized blood vessels (Figure 7D). In the epididymis (Figure 7E), transcripts were present in the vessels surrounding the tubules and in cells of the interstitia. Figure 7F represents the liver hybridized with the sense SC1 riboprobe.
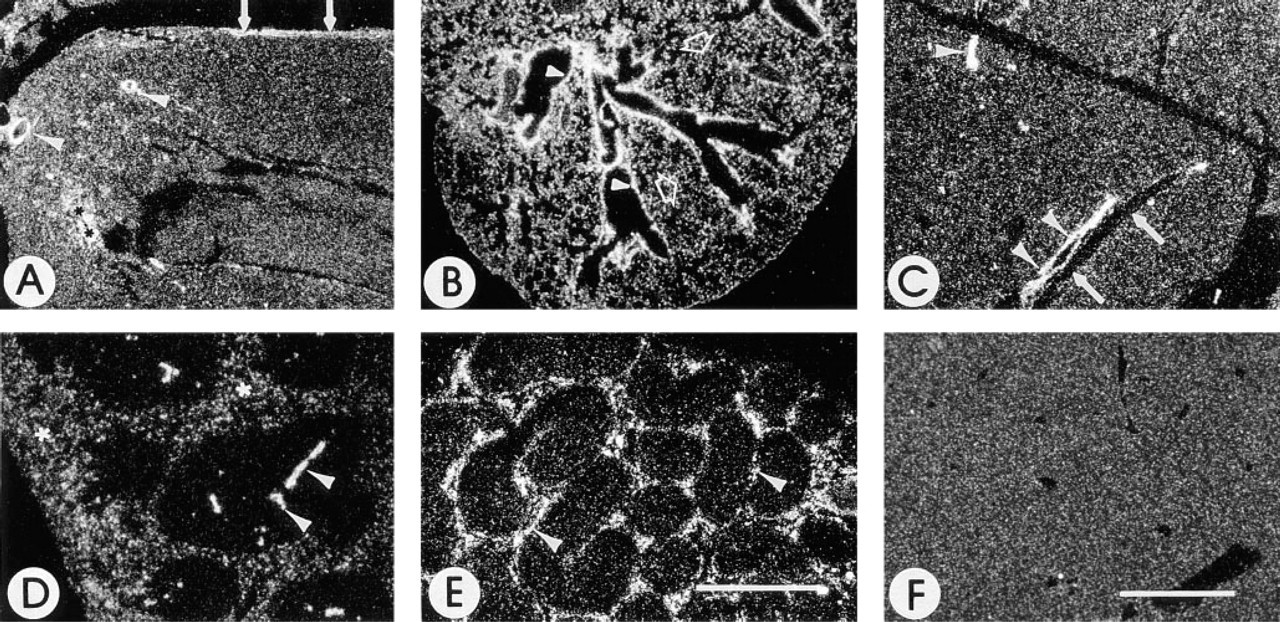
Expression of SC1 in organs of adult mouse. Shown are darkfield photographs of sections of tissues of an adult mouse after in situ hybridization with an anti-sense (
Highly vascular organs were studied to examine further the expression of SC1 in vessels of different sizes. In all tissues examined, expression of SC1 mRNA in spleen and liver was observed in the media and adventitia of medium and large vessels but not in the endothelial cell layer (Figure 8). Figure 8A shows that, in the spleen, the endothelial cells were devoid of SC1 transcript, but the media and extracellular matrix surrounding arterioles contained relatively high levels of mRNA. Moreover, no transcripts were detected in the sinus adjacent to the arteriole. Figure 8B shows the concentration of signal in the media and adventitia of the vessel but not in the endothelial cell (intimal) layer or the adjacent sinus of the liver. In contrast, expression of SC1 mRNA was generally negative in small arterioles and venules of the liver (Figure 8C). SC1 transcripts were present at low levels throughout the capsule, medium-sized vessels, and parenchyma of lymph nodes (data not shown). Small vessels of the lymph node, such as the high endothelial venules, were not distinguishable, and localized transcripts were, therefore, difficult to detect.

Distribution of SC1 in vessels. Shown is a combination bright- and darkfield photograph of a large sinus (left) and medium-sized artery (right) in the spleen (
Mouse embryos from various stages of development were examined to determine whether the distribution of SC1 was regulated in a differential manner during embryogenesis (Figure 9). At Day 12 (Figure 9A), SC1 mRNA was diffuse throughout the central and peripheral nervous system and in developing bone. Similar levels of expression were noted in the Day 13 embryo (Figures 9B-9C). Figure 9D represents the brain of a Day 15 embryo. As noted during other time points of development, medium and large-sized vessels as well as the meninges of the brain expressed SC1 mRNA. Transcripts for SC1 were also noted throughout the brain parenchyma. At Day 16, SC1 transcripts were also prominent in the lung, interstitial tissue, and again in medium and large vessels (Figure 9E). Expression in many organs, such as the heart and liver, was limited to the capsules surrounding the organs and larger blood vessels (Figure 9E). In the gut, abundant transcripts were noted in the mesentery, the vasculature, and in many of the epithelial cells (Figure 9E). As noted earlier in development, there were marked levels of expression in the spinal cord and in the developing bone of the vertebrae (Figure 9F). In the Day 18 embryo (Figures 9G-9I), SC1 mRNA was abundant in the area of the sensory nerve supply and surrounding connective tissue of the vibrissae (Figure 9G), in the spinal cord, and in the gut (Figure 9H). Bone islands, such as those of the foot, continued to express abundant transcripts for SC1 (Figure 9I).
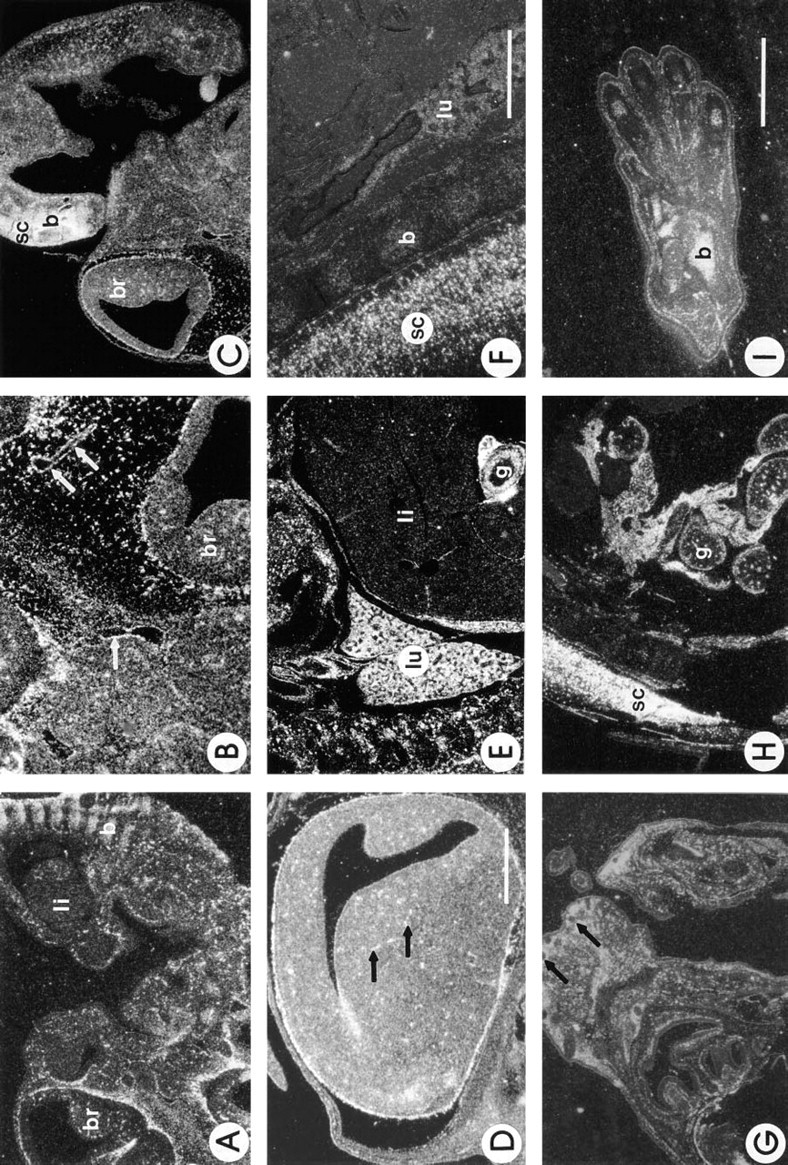
Expression of SC1 during murine development. Shown are darkfield photographs of murine embryos at 12–18 days of development after in situ hybridization with an anti-sense probe for SC1. At 12 days (
Discussion
Members of the SPARC family include albondin, follistatin-like protein (TSC-36), testican, QR1, hevin, and SC1 (Maurer et al. 1995). SPARC inhibits cell spreading on collagen and induces cell rounding in cultured endothelial cells and fibroblasts (Sage et al. 1989). SPARC also disrupts focal adhesions (Murphy-Ullrich et al. 1991) and was shown to alter the distribution of cytoskeletal elements as well as the permeability of endothelial cell monolayers (Goldblum et al. 1994). SPARC, testican, QR1, hevin, and SC1 all exhibit four distinct regions or domains which, in the case of SPARC, function independently or cooperatively to modulate cell-matrix interactions. Of the proteins mentioned above, SC1 displays the highest amino acid similarity with SPARC.
SC1 is thought to play an important role in neural development. Mendis et al. (1996) have shown an increase in SC1 during development of the brain, with the highest levels in the adult associated with neurons and synaptosomes. These authors also found that SPARC was expressed in the adult brain by astrocytes in synapse-enriched regions and suggested a role for SPARC in neural plasticity (Mendis et al. 1995). Moreover, the cytokine-binding follistatin region in both of these proteins has been suggested to function in neural differentiation through the accumulation, protection, and regulation of the activity of growth factors (Mendis et al. 1996).
Hevin (human SC1) is a protein associated with the basal, lateral, and apical surfaces of human high endothelial venule (HEV) cells and is not expressed in the underlying basement membrane (Girard and Springer 1996). Purified hevin does not support adhesion of human umbilical vein endothelial cells in vitro; in fact, addition of soluble, exogenous hevin inhibited attachment and spreading of endothelial cells on fibronectin substrates (Girard and Springer 1996). These results are similar to those reported for the addition of exogenous SPARC to cells (Lane and Sage 1994). Peptides generated against the N-terminal and C-terminal EF hand regions of SPARC (peptides 1.1 and 4.2, respectively) were shown to inhibit cell spreading of endothelial cells and fibroblasts in a concentration-dependent manner (Lane and Sage 1990). From the similarities in the structure of these two proteins, we predict that the N-terminal region of hevin and/or its EF hand domain is responsible for this inhibition of cell attachment and spreading. SPARC, the tenascins, osteopontin, and thrombospondin 1 have been described as matricellular proteins, a group of modular extracellular proteins whose functions are achieved by binding to matrix proteins as well as to cell surface receptors, or to other molecules such as cytokines and proteases that also interact with cells (Bornstein 1995). On the basis of its similarity, in both structure and function, with SPARC, hevin (SC1) could also be described as a matricellular protein.
Both SC1 and SPARC are expressed in many organs in the mouse. Ribonuclease protection assays showed similarities in tissue expression but notable differences in relative abundance between these two mRNAs. Moreover, in situ hybridization demonstrated that transcripts for these two proteins were not expressed in the same regions of a particular organ or by the same cell type. For example, SC1 was expressed almost exclusively in the medium and large vessels of the kidney, whereas SPARC transcript was seen in the glomeruli as well as the vasculature of the kidney. During development, patterns of expression of SC1 were similar to those found in the adult mouse, with prominent levels in the brain and spinal cord, and in the media and adventitia of vessels. After the development of the gut, the mesenteric membrane of the small intestine showed abundant transcripts for SC1 mRNA. This supporting structure is composed of fibrous tissue containing a network of fibroblasts and mesothelial cells that synthesize a number of ECM proteins, including collagens and elastin. In spite of the high levels of expression in cells that provide structural support in vivo, cultured smooth muscle cells and fibroblasts isolated from mouse aorta had no detectable SC1 mRNA. Conditions conducive to transcription of SC1 mRNA, which might be lacking in vitro, include the presence of specific extracellular matrix components and/or growth factors and a three-dimensional network.
Although SC1 and SPARC share a similar pattern of tissue distribution, they are not found in the same structures within organs. However, the cell adhesion studies done by Girard and Springer (1996) have shown that these two proteins do exhibit similar functions with respect to the inhibition of attachment and spreading of endothelial cells in culture. In addition, the coincident expression of SC1 and SPARC mRNA in some organs (e.g., adrenal cortex and brain) supports the possibility that these proteins could compensate for each other functionally. It has been recently suggested, however, that thrombospondins 1 and 2 have distinct functions in the adrenal cortex, despite their homology and coincidence in this tissue (Lafeuillade et al. 1996). Studies on mice with targeted disruption of either the SC1 or SPARC gene should address the question of compensation and will provide a more complete understanding of their functions in vivo.
Footnotes
Acknowledgements
Supported by the National Institutes of Health (GM-40711).
We thank Dr L. Isom for guidance in the initial experimental design of this project, and S. Soderling and Dr K. Motamed for helpful discussions. We also thank Dr J. Boulter of the Salk Institute for his generous gift of cDNA libraries.