
Abstract
The spatial expression patterns of the two α-carbonic anhydrase genes, CA VII and CA-RP VIII (called Car7 and Car8 in the mouse) were examined in the mouse brain by in situ hybridization. These two genes are the most highly conserved evolutionarily among the mammalian α-CAs. Both genes showed a similarly wide expression pattern in the brain. In the cerebrum, mRNA expression was detected in the pia, choroid plexus, and neurons of the cortical layer, thalamus, and medial habenulae. A high level of expression appeared in the pyramidal and granular cells of the hippocampus. In the cerebellum, both Car7 and Car8 were transcribed to different degrees in the Purkinje cells, and a lower expression level occured in the molecular and granular cell layers. Transcription signals for both genes were excluded from the white matter regions.
E
CA activity was demonstrated in neural tissues as early as the 1940s. This was further confirmed by many subsequent studies carried out by many different investigators. Most of these studies used enzymatic cytochemical approaches that detect CA activity without identifying the specific isozyme. These studies localized the CA activity extraneuronally to the oligodendroglial and choroid plexus cells (Kazimierczak et al. 1986; Ridderrstralle and Hanson 1985; Giacobini 1962), pericytes (Kazimierczak et al. 1986), ganglionic cells of the peripheral nervous system (Kazimierczak et al. 1986; Korhonen and Hyppa 1967), and the capillaries and axons of the CNS (Nogradi and Mihaly 1988; Ridderstralle and Hanson 1985; Riley et al. 1984). Only a few studies indicated the presence of carbonic anhydrase in the neuronal cell bodies of the CNS (Nogadi and Mihaly 1992; Nogradi et al. 1989; Aldskogius et al. 1988; Wong et al. 1983, 1987). Alternatively, some investigations localized specific isozymes in the mammalian CNS, e.g., CA II in oligodedrocytes (Ghandour et al. 1980), retinal Muller cells (Linser al. 1984), CA III in ameboid microglia cells and choroid plexus epithelial cells (Nogradi et al. 1993; Lyons et al. 1991), and CA IV in capillaries and myelin (Brion et al. 1994; Ghandour et al. 1992). The majority of these studies detected proteins using specific antibodies.
In this study, we examined by in situ hybridization the mRNA expression of one catalytic CA (CA VII) and one acatalytic CA (CA-RP VIII) in the mouse brain. The amino acid sequences of their protein products are the most highly conserved of the α-CAs between the human and mouse homologues, the percent sequence identity being 95% for the CA VIIs and 98% for the CA VIIIs (Ling et al. 1995; Skaggs et al. 1993), which suggests an important biological function for both genes.
The CA VII gene was isolated and characterized from a human genomic library (Montgomery et al. 1991). The derived amino acid sequence showed that its critical catalytic site residues have remained strictly conserved, suggesting that this gene codes for a catalytic enzyme. Recently, mouse CA VII was expressed in E. coli and was demonstrated to possess CO2-hydrase activity (Lakkis et al. 1996). CARP VIII, initially termed CA-related protein (CARP), was first identified in a mouse brain cDNA library (Kato 1990). However, its deduced amino acid sequence indicated changes in the critical catalytic site residues that would be expected to abolish the CO2-hydrase activity characteristic of all known CAs. Initially, it was believed to be exclusively expressed in the Purkinje cells of the mouse cerebellum (Kato 1990). However, it was later found to be expressed in human testis, salivary glands, placenta, and lung as well (Ling et al. 1994; Skaggs et al. 1993).
Throughout this report, the mouse CA VII and CA-RP VIII genes will be given the designations Car7 and Car8, respectively, in keeping with nomenclature policy for the naming of murine CA genes.
Materials and Methods
Tissue Preparation
Healthy 18-day-old female mice (C57 B1/6J) were sacrificed according to the guidelines of the University of Michigan Commitee on the Use and Care of Animals (UCUCA), and their brains were rapidly isolated. The cerebrum and the cerebellum were separated, frozen in isopentane cooled to – 30C and stored frozen at –80C. They were then equilibrated to cryostat temperature (– 18C), mounted in OCT medium (Lab-Tek Division; Miles, Elkhart, IN), and frozen as solid blocks, which were later cryosectioned at 12-μm thickness. Sections were thaw-mounted on poly-
Riboprobe Preparation
Specific probes for each gene were prepared by reverse transcription-polymerase chain reaction (RT-PCR) of mRNA isolated from the mouse brain. This mRNA was first reverse-transcribed into cDNA by superscript II (Life Technologies; Bethesda, MD). The cDNA probes for both Car7 and Car8 genes were made by PCR of this cDNA template using Car7-and Car8-specific sets of primers. The amplified cDNAs for Car7 (446
In Situ Hybridization and Autoradiography
The in situ hybridization procedure was an adaptation of previously described methods (Wilkinson and Nieto 1993; Cox et al. 1984). The frozen mouse brain sections were thawed at room temperature and then fixed in 4% paraformaldehyde (Sigma; St Louis, MO) for 30 min, washed in sterile dH2O, then treated with 0.2 N HCl for 5 min, washed in dH2O, and treated with proteinase K (1 μg/ml) at room temperature for 5 min. These sections were then acetylated with 0.1 M triethanolamine containing 0.25% acetic anhydride (both from Sigma) for 10 min. Hybridization was performed by applying 50 μl of hybridization solution [50% formamide, 0.6 M NaCl, 10 mM Tris (pH 7.5). 1 mM EDTA, 50 μg/ml heparin, 10 mM DTT, 0.5 mg/ml carrier DNA, 0.5 mg/ml tRNA, 10% PEG 8000, 1 x Denhardt's], containing 1 x 104 cpm/μl 35S-labeled probe directly on the well-drained sections and incubating them at 55C overnight. The slides were then washed twice in 2 x SSC and 50% formamide, 2 x SSC at 65C for 30 min each, followed by treatment with RNAse A (200 μg/ml) at 37C for 30 min and a final wash in 0.1 x SSC at 55C for 30 min. Autoradiography was then performed by dipping the slides in Kodak NTB2 emulsion. Exposure time was about 8 days, after which slides were developed and fixed in Kodak developer and fixer, respectively. Finally, slides were stained with 5% aqueous Giemsa stain (Sigma) for 2 min, dried, and cover-slipped. Photography was carried out using Kodak TMAX 100 black-and-white film under darkfield illumination.
Results and Discussion
In this study, we used in situ hybridization of 35S-labeled anti-sense riboprobes, specific for Car7 and Car8, to examine the regional distribution in the mouse brain of mRNAs encoded by these two functionally different CA genes. The enzyme histochemical studies used earlier for detecting CA expression in animal tissues did not demonstrate the specificity of the isozyme. These methods are unable to detect the acatalytic isozymes, such as CA-RP VIII (Car8).
The two riboprobes specific for each of these two genes were transcribed from two cDNA clones isolated from mouse brain by RT-PCR. The specificity of the hybridization signal was verified by using sense riboprobes for both genes that produced (in both cases) only a weak background random scatter of grains (Figures 1A, 1B, and 2A-2C).
The transcripts of the two genes displayed slightly different anatomic distributions in the cerebellum (Figure 1). Both were expressed at a relatively high level in the Purkinje cells, although the Car8 transcription signal (Figures 1E and 1F) was slightly stronger. A transcription signal for both genes appeared in the granular layer, but in this case the signal for Car7 (Figures 1C and 1D) appeared to be stronger than that of Car8. In addition, a low level of mRNA expression for both genes was present in the molecular layer of the cerebellum. It is not clear whether this positive signal is due to late-migrating granular cells or to scattered basket and stellate cells present in the molecular layer. There was no detectable signal for either gene in the white matter.
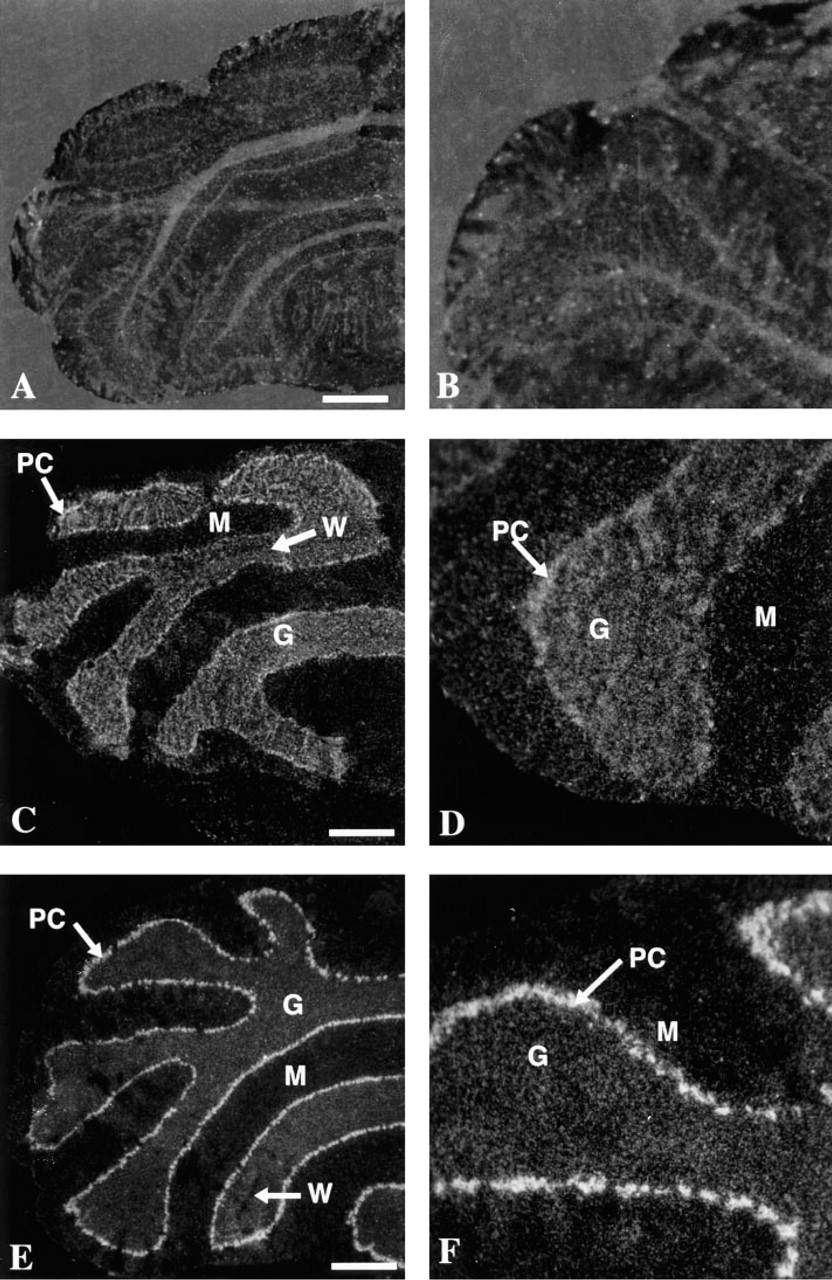
Darkfield photomicrographs illustrating the in situ hybridization of Car7 and Car8 on cross-sections of mouse cerebellum. (
The mRNA from both genes appeared to be distributed throughout the cerebrum in a fairly similar pattern (Figure 2). An mRNA expression signal appeared at the pial surface and in the large neurons throughout the cortical layers. A very strong transcription signal was present in the hippocampal formation, specifically in the pyramidal cells of Ammon's horn and in the granular cells of the dentate gyrus. In addition, a moderately strong signal for both genes was present in the medial habenulae and neurons of the thalamus. The signal appeared to be absent in the glial end-feet (glial limitans) at the pial surface and in the subventricular zone. In general, signal was also absent in the white matter layer. No expression was seen in blood vessels and capillaries. However, signal appeared in the choroid plexus and cerebrospinal fluid-containing channels (not shown).
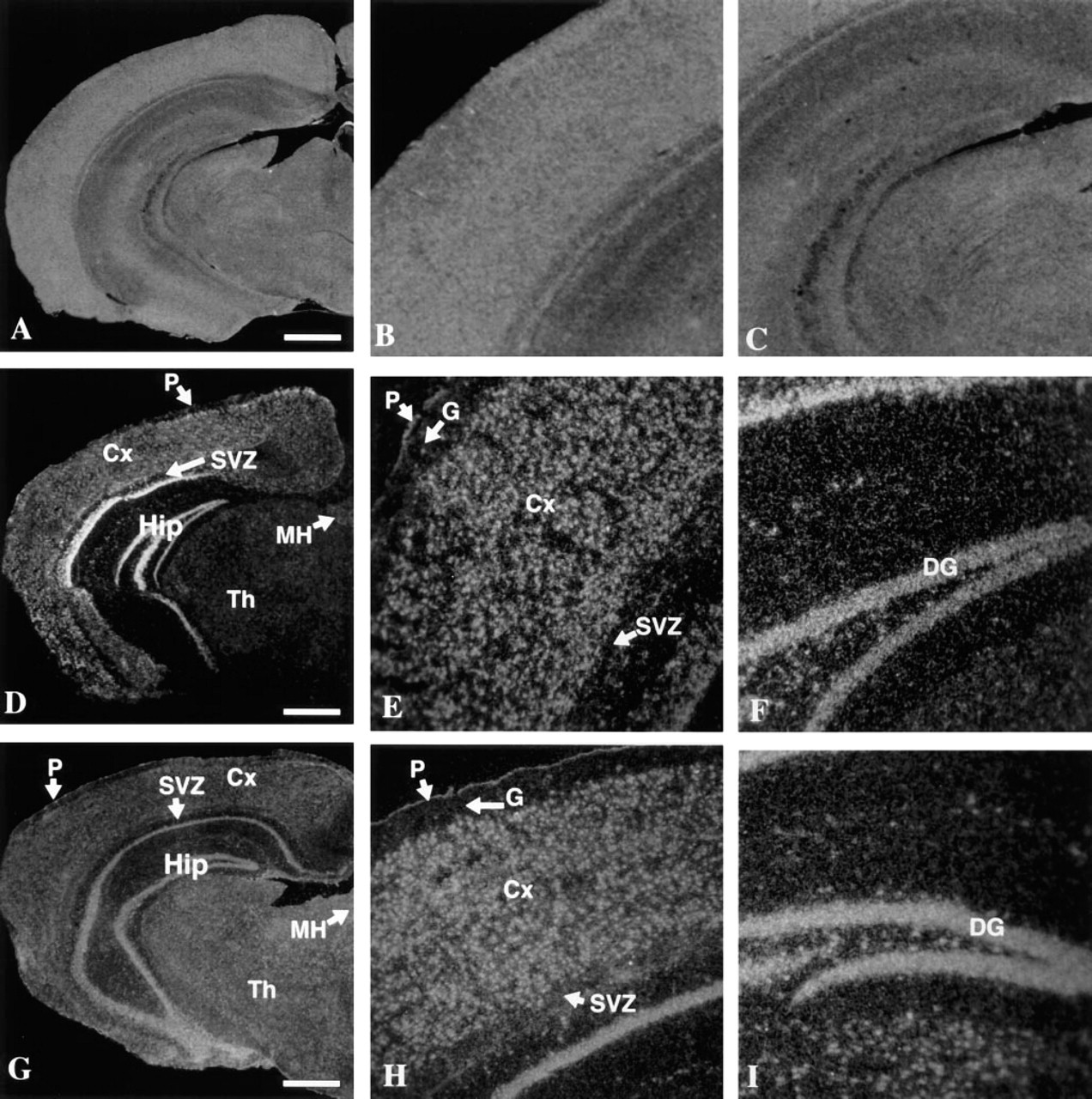
Darkfield photomicrographs illustrating in situ hybridization of Car7 and Car8 to cross sections of the mouse cerebrum. (
The significance of the expression pattern of these two genes is not clear. Even though their expression appeared to be overlapping in the cerebrum, there was a slightly different pattern of expression in the cerebellum. Cross-hybridization is very unlikely under the experimental conditions used because the two sequences shared no more than 52% identity in their nucleotides, in addition to the insertion or deletion of 10 nucleotides between the two sequences. These results demonstrate a wide distribution of mRNA for both genes, suggesting a nonspecific but rather generalized function for the products of these genes in the brain. Their expression is characteristically strong in neurons. Clearly, because CA-RP VIII lacks CO2-hydrase activity, the two gene products are expected to have very different functions.
One can speculate on the possible function(s) of CA VII on the basis of what is known about other active CAs. CA activity is known to play a role in cerebrospial fluid production and in regulation of its ionic constituents and pH (Maren and Vogh 1980). Neuronal CA VII could also be important for the rapid elimination and transport of CO2 produced by glycolysis (Giacobini 1987; Van Gelder 1983), thus enhancing and maintaining metabolic activity of the neurons. It was also suggested that CA activity influences neuroexcitation and susceptibility to seizures in many different ways (Ghandour et al. 1992; Wistrand 1984). In addition, it may play an important role in maintnining different membrane transport processes by facilitating CO2 transport and by regulation of transmembrane fluxes and appropriate distribution of chloride ions (Carr et al. 1989; Giacobini 1987; Van Gelder 1983). Carr et al. (1989) reported some relationship between CA activity in the dorsal root ganglia and cytochrome oxidase activity, which is coupled to energy requirements and electrical activity of the neurons. Giacobini (1987) also suggested that high concentrations of CA are required during the early growth and maturation phases of neurons.
Peyronnard et al. (1986) reported that peripheral nerve injury reduces CA enzyme content or activity in normally CA-positive dorsal root ganglion cells. It is also possible that the neurological side effects observed in humans after administration of CA inhibitors (e.g., acetazolamide), which have been attributed to the inhibition of CA II (Wistrand 1984) or CA IV (Ghandour et al. 1992), could also be due to inhibition of the active CA VII.
It is more difficult to speculate on the possible function(s) of CA-RP VIII. The fact that 19 acidic amino acids (i.e., 15 Glu, 4 Asp) are found in the first 35 N-terminal residues of human and mouse CA-RP VIII (Skaggs et al. 1993) is consistent with the possibility that CA-RP VIII may be involved in transcription or translation. Similar acidic zones are known to be characteristic of transcriptional activators such as GAL4, GCN4, and Herpesvirus VP16 (Gill et al. 1990; Triezenberg et al. 1988; Hope and Struhl 1986). Also of interest is the observation that the neurologically defective mouse (lurcher) lacks CA-RP VIII in the cerebellum, suggesting a possible role in neurological function (Kelly et al. 1994).
In the longer term, it would be of interest to determine if alterations in expression of these genes can be correlated with different physiological conditions or activities of the animal. The nonspecific expression of these genes is not a characteristic only of the brain, because they are also expressed in a variety of other tissues during embryonic development (Lakkis et al., in press) as well as in adults, implying an important general role in the various tissues or perhaps very different roles.
Acknowledgments
Supported by NIH grant GM24681.
We thank the Morphology Core Facility for assistance in tissue sectioning and the Cell Biology Laboratory, particularly Bruce Donohoe and Thomas Komorowsky, at the University of Michigan, for assistance in microscopy and computer scanning.