
Abstract
We have devised a simple method that combines retrograde labeling of projecting neurons and in situ hybridization histochemistry to examine mRNA expression in the retrogradely labeled neurons. First, projecting neurons were retrogradely labeled in vivo by injection of the lipophilic neuronal tracer DiI. The fluorescence of the labeled neurons in the brain slices was photoconverted into stable DAB precipitate by green light illumination. The slices were cut into thinner sections and processed for detection of specific mRNA by in situ hybridization. Using this highly sensitive method, we demonstrate here that the corticospinal tract neurons in newborn rats express mRNA for the cell adhesion molecule L1. TAG-1 mRNA was not detected in these neurons. Therefore, the present method provides an important tool to study the molecular expression of projection neurons during the development of neuronal circuitry.
Keywords
T
For combining in situ hybridization histochemistry with retrograde labeling, fluorescent tracers were used in most cases to label neuronal somata (Burgunder and Young 1988). However, the fluorescence tends to disappear during the subsequent procedures required for in situ hybridization because of the repeated rinsing in buffers and organic solvents. When enzymes such as HRP and WGA-HRP were used as retrograde tracers, proteinase treatment caused the activity to deteriorate. When the enzymes were reacted for visualization before in situ hybridization, reaction procedures involving many rinses in buffers caused ribonuclease contamination and reduced the sensitivity for in situ hybridization. Therefore, to overcome these problems we developed a method that combines DiI retrograde labeling (Honig and Hume 1989), photoconversion into DAB reaction product, and in situ hybridization histochemistry. Our protocol was tested for detection of mRNA expression of cell adhesion molecules L1 and TAG-1 (Yoshihara et al. 1995; Wolfer et al. 1994; Dodd et al. 1988) in DiI-labeled Layer V pyramidal neurons in developing rat neocortex.
Materials and Methods
In Vivo Retrograde Labeling of DiI
Sprague-Dawley rats staged at postnatal Day 1 (P1; day of birth = PO) were deeply anesthetized by hypothermia. The dorsal surface of the spinal cord was exposed at cervical level 2-3 by removing vertebral arches, adhesive muscles, and adipose tissue. A small crystal of DiI (1, 1'-dioctadecyl-3,3,3',3'-tetramethylindocarbocyanine perchlorate; Molecular Probes, Eugene, OR) was injected into the median plane of the spinal cord (Honig and Hume 1989). Two days later the rats were given an overdose of sodium pentobarbital (100 mg/kg IP) and perfused transcardially first with 0.9% saline and then with fresh RNAse-free 4% paraformaldehyde in 0.1 M PBS, pH 7.4. All solutions for these and the following procedures were made using water treated with 0.1% diethylpyrocarbonate (DEPC; Sigma, St Louis, MO) and autoclaved to inactivate ribonuclease. The brains were removed and kept in the same fixative at 4C overnight.
Photoconversion
The brains were cut into slices 200μm thick with a microslicer. The slices were immersed in 3,3'-diaminobenzidine (DAB: 1 mg/ml in 0.1 M Tris, pH 7.4) solution and irradiated with green light to photoconvert the DiI fluorescence into DAB precipitation (Sandell and Masland 1988). Irradiation was performed with an inverted fluorescent microscope system IMT-2-RFC (Olympus; Tokyo, Japan) using a X 20 objective lens (SPLAN Apo-20), a 100-W mercury lamp (USH-102D), a power supply (BH2-RFL-T3-100), and a G filter (IMT-2-DMG; peak excitation wavelength 546 nm). During irradiation, DiI fluorescence faded completely and brown DAB precipitation appeared gradually. The slices were rinsed in PBS and immersed in 30% sucrose/PBS for several minutes for cryoprotection. The slices were spread on slide glasses, covered with OCT compound (Miles; Elkhart, IN), frozen on dry ice, and stored at −80C until use.
In Situ Hybridization
We essentially followed a standard protocol (Simmons et al. 1989) for in situ hybridization. Briefly, the slices were cut into thinner 15-μm sections with a cryostat. The sections were mounted on poly-
Results
Two days after DiI injection into the spinal cord (Figure 1 A), fluorescence was detected in the neocortex (Figure 1 B). The laminar position of the labeled neurons in the section indicated that they were Layer V pyramidal neurons. DiI labeling was observed as granules in the cell somata and dendrites (Tamamaki and Nojyo 1995). When the DiI fluorescence was photo-converted into DAB precipitate, brown granules appeared in the somata and dendrites of the pyramidal neurons under brightfield microscopy as shown in Figure 1 C.
The laminar expression patterns of TAG-1 and L1 mRNA were visualized by in situ hybridization in the P3 rat neocortex (Figures 2B and 2C). Comparison of the laminar position of the retrogradely labeled neurons (Figure 2 A) with laminar distribution of the mRNAs (Figures 2B and 2C) in adjacent sections suggests that the Layer V pyramidal cells express L1 but not TAG-1. However, with such a comparison it is not possible to determine whether or not the retrogradely labeled Layer V pyramidal neurons express L1 and TAG-1.
Using the combination of retrograde labeling and in situ hybridization methods, we demonstrated that L1 mRNA was expressed by the retrogradely labeled pyramidal neurons. Silver grains of hybridization signals of L1 mRNA overlapped the areas marked by brown DAB precipitation, as shown in Figure 3 A. However, intense signals of TAG-1 mRNA were not observed in the retrogradely labeled pyramidal neurons.
In the same section, we compared the in situ hybridization signals between areas with and without photoconversion but could not detect any differences between them. In addition, DAB precipitation did not appear to decrease after the in situ hybridization procedure (Figures 3A-3D). Nonspecific binding of cRNA sense probes to the areas of DAB precipitation was not detected (Figures 3B and 3D).
Discussion
The method presented here combines DiI labeling of cortical pyramidal neurons and in situ hybridization histochemistry. One concern in combining the methods was the possibility that DiI labeling and/or DAB photoconversion might cause problems in the following in situ hybridization steps. Another concern was the possibility that the in situ hybridization method might cause disappearance of the retrogradely transported DiI. Although the photoconversion of fluorescent labeling into stable DAB precipitate was carried out before in situ hybridization, it involved only a single incubation in the DEPC-treated DAB solution. Messenger RNA is resistant to illumination with visible light, as demonstrated in the photobiotinylation of RNA (Forster et al. 1985). The results presented here showed that the sensitivity for detection of both retrograde tracer and mRNA was not markedly reduced by the present method.
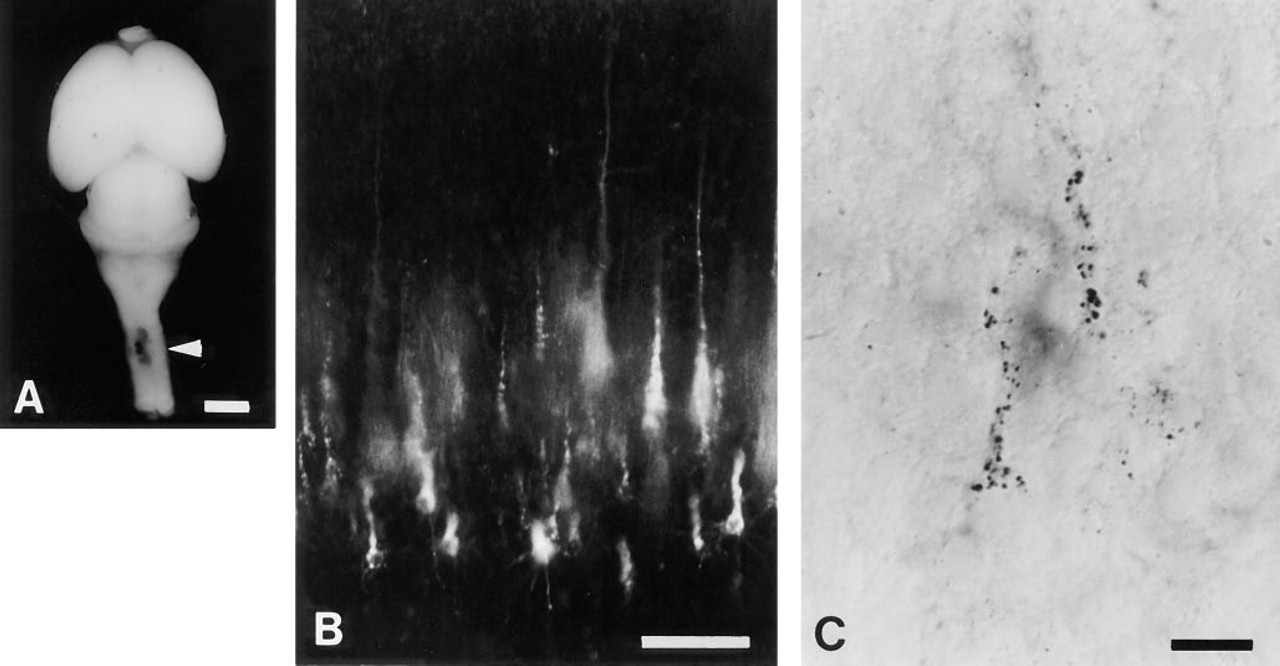
(A) A DiI injection site in the P3 rat spinal cord. The arrowhead indicates the injection site. Bar = 2 mm. (B) DiI-labeled Layer V neurons in the neocortex. Bar = 100 μm. (C) DAB precipitation in the neurons photoconverted from DiI labeling. Bar = 20 μm.
DiI is an excellent anterograde and retrograde tracer with high sensitivity in both fixed and unfixed tissues. Therefore, DiI is suitable for investigation of neuronal circuits not only in embryos but also in the adult central nervous system (Tamamaki and Nojyo 1995; Koester and O'Leary 1994; Honig and Hume 1989; McConnell et al. 1989). In the present method, retrograde signals were observed as brown precipitate and in situ hybridization signals were seen as black dots (Figure 3). Overlap of these signals was easily observed under brightfield microscopy. Because the DAB precipitate is light-reflective, it can not be discriminated from the silver grains of in situ hybridization signals under darkfield microscopy. Recently, non-radioactive digoxigenin (DIG)-labeled cRNA probes have been widely used for in situ hybridization (Aigner and Pette 1990). The present method is applicable to the DIG in situ hybridization procedure. DIG-labeled signals are observed as dark blue precipitate under the condition of alkaline phosphatase reaction. Because the dark blue reaction products are non-reflective and DAB precipitate is reflective under darkfield microscopy, overlap of these two kinds of signals is detectable with a combination of dark- and brightfield illumination.
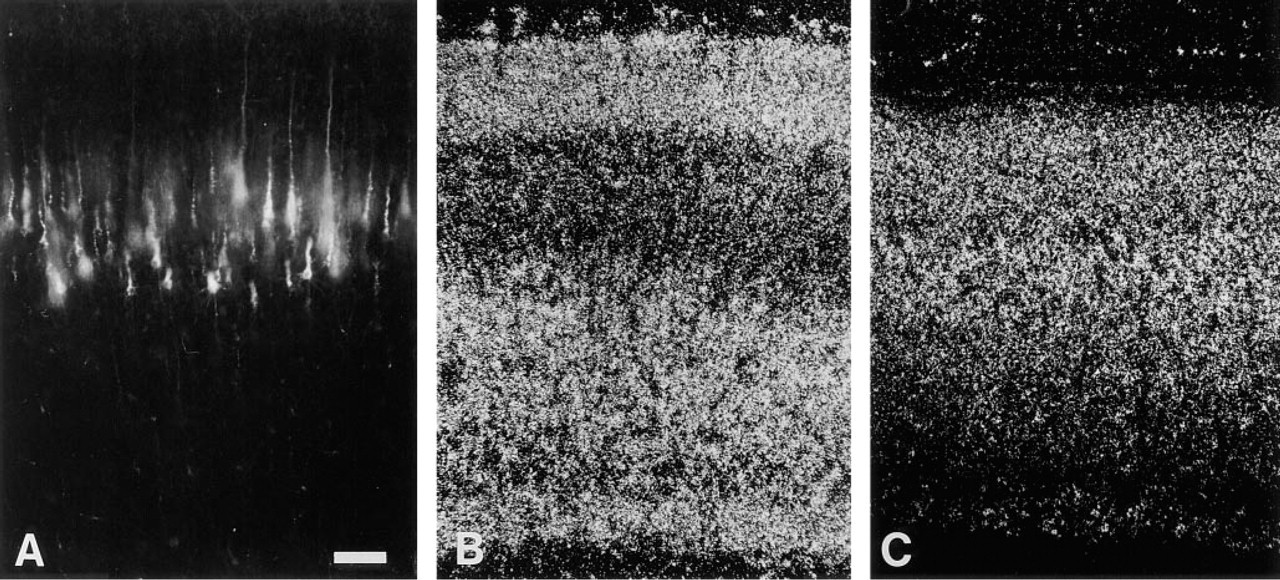
P3 neocortical sections under same magnification. DiI-labeled Layer V neurons. (A) In situ hybridization signals for TAG-1 mRNA (B) and L1 mRNA (C) with dark field illumination. The pia mater is located at the top of all figures. Bar = 100 μm.
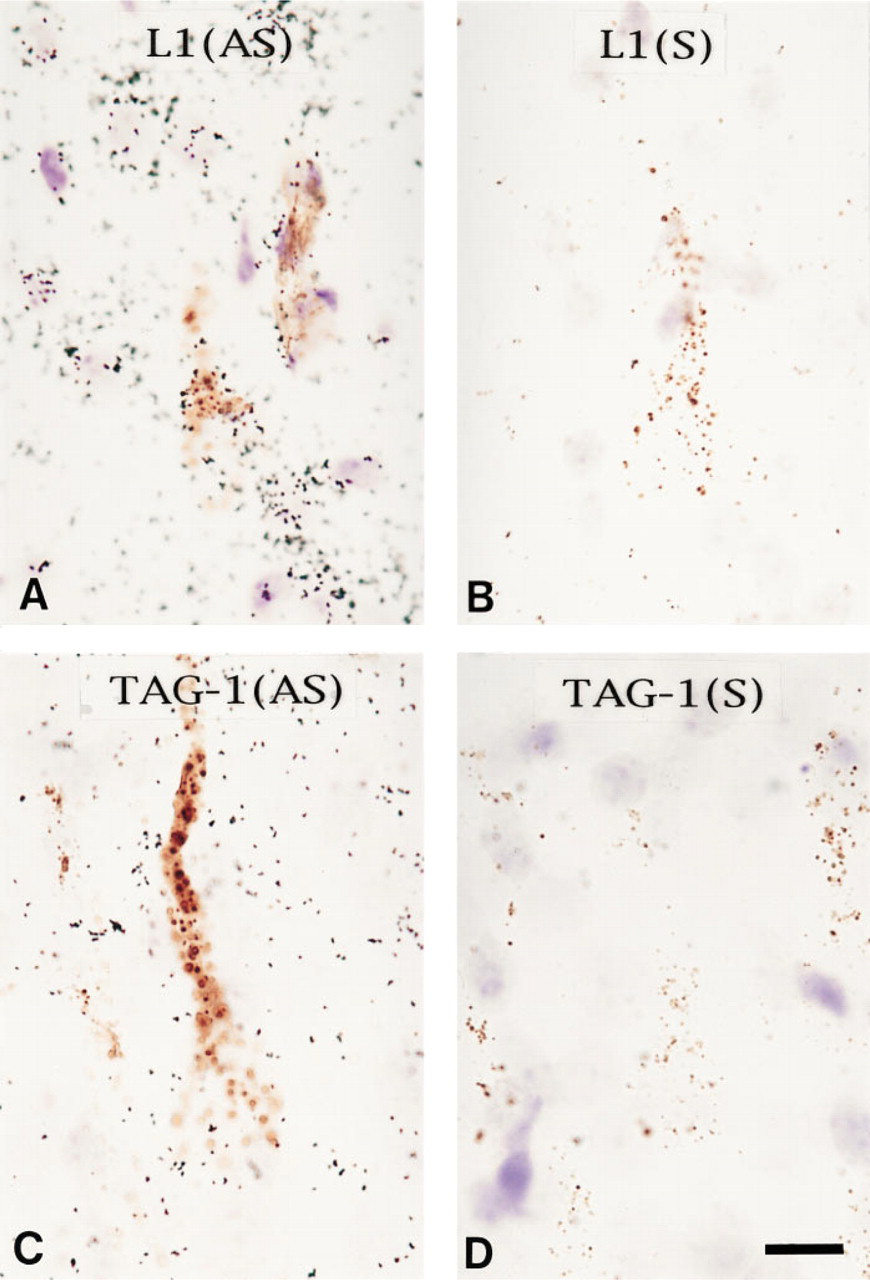
In situ hybridization analysis of retrogradely labeled Layer V neurons. Most retrogradely labeled layer V neurons expressed L1 at a high level, but not TAG-1. (A) In situ hybridization signals detected with L1 anti-sense cRNA probe on the retrogradely labeled Layer V neurons. Brown precipitation is DAB product photoconverted from DiI. Black dots are silver grains indicating [35S]-cRNA hybridization. (B) In situ hybridization with L1 sense probe. (C) In situ hybridization with TAG-1 anti-sense probe. (D) In situ hybridization with TAG-1 sense probe. Bar = 10 μm.
In this technique, there are several problems that should be overcome with repeated trials. First, single DiI injection into the spinal cord reveals only a fraction of the corticospinal projection neurons in the neocortex. The area of illumination through a X 20 objective lens was so small that only a few DiI-labeled pyramidal neurons were illuminated at one time. Therefore, to collect enough of double-labeled samples and to draw a reliable conclusion, many sections should be processed for in situ hybridization with each cRNA probe. Second, in autoradiography, the emulsion-dipped sections should not be exposed for too long. If the silver grains cover the DAB precipitation, it may be hard to detect overlapping of the two signals under bright-field microscopy.
To demonstrate the usefulness of the double-labeling method, we used it to reveal that the corticospinal projecting Layer V neurons express L1 mRNA but not TAG-1 mRNA in newborn rats. Recent immunohistochemical study revealed that the pyramidal tract in developing mouse has TAG-1 immunoreactivity (Wolfer et al. 1994). However, there is no discrepancy between their observation and our result because the pyramidal tract is not the tract only for the Layer V pyramidal neurons. TAG-1-immunoreactive axons in the tract may have their somata in other locations than Layer V of the cortex.
Because the present method is applicable to a wide variety of molecules involved in the control of axonal projection, it offers an important tool for the study of molecular mechanisms in formation of the neuronal circuitry.
Footnotes
Acknowledgements
Acknowledgment
Supported by grant-in-aid #08271215 for Scientific Research on Priority Areas on “Functional Development of Neural Circuits,” Ministry of Education, Science and Culture (to NT).