
Abstract
To study the lifespan of immune cell populations in the LEW rat, 5-bromo-2-deoxyuridine (BrdU) was administered in the drinking water. After 12 weeks, the epithelium of gut and skin was completely BrdU+. In contrast, thymus, bone marrow, and germinal centers of Peyer's patches contained only a few BrdU+ cells, although most should have been labeled during this time. The lack of labeling was due neither to obvious toxic effects of BrdU on these organs nor to insufficient detection of incorporated BrdU. Analysis of the kinetic pattern of the appearance of BrdU+ cells in bone marrow, blood, spleen, and lymph nodes over 12 weeks revealed that the dosage of BrdU initially was high enough to label the proliferating cells in the bone marrow, but then became too low, although the BrdU uptake of the rats was similar over the entire time. This indicates that in the LEW rat the metabolism of orally applied BrdU changes over time, leading to a reduction in the amount of BrdU available for incorporation into the DNA below a level necessary for labeling all proliferating cells. This effect appears to be species- and strain-dependent, and should be considered when the BrdU technique is used.
Keywords
THE LIFESPAN of B- and T-lymphocytes and natural killer cells is an important factor in regulating the size of the respective population, and thereby determines to a great extent the capacity of the immune system. One possibility for studying the lifespan of cell populations is to label all newly formed cells over a defined period of time with BrdU, a DNA precursor (Gratzner 1982). The advantage of BrdU over [3H]-thymidine, the “gold standard,” is that it can be given continuously in drinking water, thereby avoiding stress induced by repeated IV injections (e.g., von Boehmer and Hafen 1993). This is important because it is known that stress alters, among other things, lymphocyte proliferation (e.g., Freitas et al. 1986). In addition, problems regarding the handling of radioactivity do not arise, especially when large animals are used, and time-consuming autoradiography is not necessary because the incorporated BrdU can be revealed by immunostaining in cytological (e.g., Westermann et al. 1989) and histological preparations (Westermann et al. 1996; Wynford-Thomas and Williams 1986). However, it is known that even low concentrations of BrdU may be toxic, teratogenic, and mutagenic (Goz 1978) and that chronic exposure to BrdU influences the proliferation rates of hepatic and renal cells in mice (Weghorst et al. 1991).
Therefore, in the present study not only the proliferation of B- and T-lymphocytes and natural killer cells in the LEW rat was determined but also great care was taken to detect possible adverse effects of long-term BrdU application in this species. LEW rats were given an often-used BrdU dosage in the drinking water over 12 weeks (1 mg/ml) (e.g., Kroese et al. 1995; Tough and Sprent 1994; Förster and Rajewsky 1990; Gray 1988), and on cytological and histological preparations both the phenotype and the incorporated BrdU were revealed. To control for toxic effects of BrdU, not only B- and T-lymphocytes and natural killer cells but also monocytes and granulocytes were analyzed in several lymphoid organs and at various time points after BrdU application. In addition, because the detection of BrdU in the DNA depends very much on the technique used (Hayashi et al. 1988; Moran et al. 1985), the method applied in the present study was checked by revealing incorporated BrdU in parallel in Balb/c mice, which have been often used for such studies (Tough and Sprent 1994; Rocha et al. 1990).
Materials and Methods
Animals and BrdU Application
Adult male LEW rats (n = 25) and Balb/c mice (n = 14) were kept under specific pathogen-free conditions until the beginning of the experiments. The animals had free access to food and water. Rats and mice were continuously administered BrdU (5-bromo-2-desoxyuridine; Sigma, Munich, Germany) for various times up to 12 weeks. BrdU (150 mg) was dissolved in 125 ml water and 25 ml orange juice (previous studies showed that the amount of BrdU water imbibed was reduced when the orange juice was lacking). The BrdU concentration was 1 mg/ml water according to established protocols (e.g. Kroese et al. 1995; Tough and Sprent 1994; Förster and Rajewsky 1990; Gray 1988). The drinking water was protected from light by aluminum foil and was changed twice each week, the amount of water drunk was recorded, and the BrdU uptake was calculated. Rats and mice serving as control animals were kept under the same conditions, with the exception that the drinking water did not contain BrdU.
Cell Preparations and Histological Sections
Rats were exsanguinated under ether anesthesia after 3, 17, 24, 38, 52, 59, 70, or 84 days (12 weeks) of BrdU administration. Balb/c mice were investigated after 7, 17, 24, or 70 days of BrdU administration. The number of leukocytes and the percentage of lymphocytes and granulocytes in the arterial blood were determined. The organ weights of the thymus, spleen, mesenteric lymph nodes, axillary lymph nodes, liver, right kidney, and right testicle were recorded and the organs were prepared for histology and cytology. Tissue samples of the thymus, skin, small intestine, and Peyer's patches were quickly frozen in liquid nitrogen and cryosections 5 μm thick were made. After air-drying the slides were wrapped in aluminum foil and stored at −20C. Thymus, spleen, and mesenteric lymph nodes were minced and washed with RPMI 1640 (Seromed; Berlin, Germany) at 400 X g for 10 min. The red blood cells were removed by osmotic shock. Afterwards, the cells were centrifuged through 2 ml of fetal calf serum (Seromed) at 400 X g for 10 min and resuspended in RPMI 1640 with 0.5% bovine serum albumin (BSA; Serva, Heidelberg, Germany). To obtain the bone marrow cells, the femur was flushed with RPMI 1640 containing 0.5% BSA and the cells were treated as described above. The blood was diluted 1:4 with RPMI 1640 containing 0.5% BSA. After centrifugation in Ficoll Isopaque (Pharmacia; Uppsala, Sweden) (= 1089 g/cm3 at 400 X g for 30 min) the interface layer was harvested and washed (800 X g for 10 min). Then cells were treated as described above. Cytocentrifuge preparations were made (Shandon, Frankfurt, Germany; 150 X g, 8 min). To prevent clotting, 0. 1 × 106 cells were diluted with 100 μl 0.9% NaCl (containing 2.7 mM Na-EDTA, 5% BSA, pH 7.2). The slides were air-dried overnight, wrapped in aluminum foil, and stored at −20C.
Characterization of the Phenotype and Detection of Incorporated BrdU by Immunostaining
The cell surface was stained by the alkaline phosphatase-anti-alkaline phosphatase (APAAP) technique (van der Loos et al. 1993). At first the slides were fixed for 10 min in equal parts of methanol and acetone. After washing in Tris-buffered saline (TBS) containing 0.05% Tween 20 (Serva), the slides were incubated for 30 min at room temperature in a moist chamber with the primary antibodies shown in Table 1. Then the slides were incubated with the second antibody (rabbit anti-mouse, Z259; Dako, Hamburg, Germany) and the APAAP complex (D651, Dako) for 30 min. The last two steps were repeated for 15 min each. The color reaction was carried out in red by 2 mg Fast Red (Sigma) mixed with 3 ml APAAP substrate (9.8 ml 0.1 M Tris buffer, pH 8.2, containing 2 mg naphthol AS-MX phosphate, 200 μl N,N-dimethylformamide, and 10 μl 1 M levamisole). The slides were incubated in this mixture for 25 min, then fixed in 70% ethanol and air-dried overnight. To detect BrdU, DNA was denatured with formamide (Sigma) and NaOH. Formamide (190 ml) and NaOH (1 N, 10 ml) were separately warmed to 70C and then mixed for 8 min. The slides were immersed in this solution for 30 sec. After washing with TBS, the slides were again immersed in a mixture of formamide and 0.15% 0.15 M trinatrium citrate (70C) for 15 min. After this, the cells were washed in ice water and fixed in 1% formaldehyde (30 min) and 0.2% glutaraldehyde (10 min). Slides were incubated overnight with the monoclonal antibody anti-BrdU which was dissolved in TBS/Tween (Becton Dickinson; Heidelberg, Germany). The next day, after washing with TBS, the slides were incubated with the second antibody (rabbit anti-mouse, Z251; Dako) and the APAAP complex for 30 min. After repetition of the last two steps (each one for 15 min), the color reaction was carried out in blue. For this, 2 mg Fast Blue (Sigma) was mixed with 4 ml APAAP substrate, filtered through a filter paper, and added to the slides for 20 min. The slides were counterstained with hematoxylin (Fluka; Buchs, Switzerland) and mounted in glycergel (Dako). Immunohistology was performed as described above, with the exception that BrdU was detected by the PAP technique and the color reaction was carried out with benzidine (Westermann et al. 1996).
Monoclonal antibodies used in the experiments
Evaluation
The percentages of the various phenotypes, the BrdU+ cells, and the double-positive cells (phenotype+ and BrdU+) were determined by counting at least 100 positive cells. Means and standard deviations were calculated.
Results
Long-term BrdU Labeling Did Not Cause Obvious Toxic Effects
Over the 12-week period, the daily BrdU uptake was in the range of 20 mg, leading to a total amount of orally administered BrdU of 1439 ± 103 mg (n = 10). Obvious toxic effects of the incorporated BrdU were not seen. The animals behaved normally and showed no clinical signs of illness. The histological appearance of thymus and Peyer's patches was not altered compared to control animals (Figure 1), nor was the functional capacity of the bone marrow, as indicated by a normal number of granulocytes in the blood (Table 2). In addition, the body weight, the organ weights, the blood lymphocyte numbers (Table 2), and the cellular composition of bone marrow, blood, spleen, and mesenteric lymph nodes were not altered by BrdU application compared to those of control animals (Table 3).
The Epithelium of Gut and Skin Was Completely Labeled by BrdU but only a Few Cells in Thymus and Bone Marrow Were BrdU+
As expected, sections of the gut and the skin of the rats showed that all epithelial cells had incorporated BrdU after 12 weeks of administration (Figure 2). On cytological preparations of blood, spleen, and mesenteric lymph nodes, B- and T-lymphocytes and natural killer cells could clearly be identified and incorporated BrdU revealed (Figure 3). After 12 weeks of BrdU administration, 30.3 ± 0.7% (n = 5), 28.9 ± 0.9% (n = 5), and 20.2 ± 0.6% (n = 5) BrdU+ cells were found in blood, spleen, and mesenteric lymph nodes, respectively. The percentage of BrdU+ cells among the different subsets varied considerably, being 15% for T-lymphocytes in the mesenteric lymph nodes and 75% for natural killer cells in the blood (Figure 4). B- and T-lymphocyte subpopulations (IgM+ and IgD+ cells, and CD4+ and CD8+ cells, respectively) showed a much smaller heterogeneity (15–25% of the cells of the respective subset being BrdU+). However, surprisingly few cells in the rat thymus (0.7 ± 0.4%, n = 5) and bone marrow (12.8 ± 1.0%, n = 5) were BrdU+ after 12 weeks. In the thymus, BrdU+ cells were located in the medulla (Figure 1). Cytological preparations of the rat bone marrow showed that after 12 weeks of BrdU administration 97.8 ± 0.4% (n = 5) of the monocytic cell lineage (ED1+) and 97.6 ± 2.4% (n = 5) of the neutrophilic granulocytes (RP1+) had not incorporated BrdU. In addition, in germinal centers of Peyer's patches, only a few lymphocytes were BrdU+, whereas the corona contained many BrdU+ cells (Figure 1). To check whether the low number of BrdU+ cells in thymus, bone marrow, and germinal centers of Peyer's patches in the present study might be due to insufficient BrdU detection by immunostaining, our technique was applied to Balb/c mice given BrdU (1 mg/ml in the drinking water) and was compared in the same experiment to similarly treated rats. It was obvious that most cells in the thymus and bone marrow of Balb/c mice were BrdU+ after 2 and 10 weeks of labeling (Figure 5). After 3 weeks, 90 ± 2% (n = 4) BrdU+ cells were found in the mouse thymus, as determined on cytospin preparations. After 2, 3, or 10 weeks of BrdU application 87 ± 5% (n = 2), 92 ± 2% (n = 4), and 94 ± 2% (n = 4) had incorporated BrdU into mouse bone marrow. No labeling at all could be detected when BrdU was omitted from the drinking water (Figure 5). In LEW rats, labeled cells were detected in the thymic medulla but not in the cortex (both after 2 and 10 weeks of labeling) (Figure 5). In the bone marrow of LEW rats many BrdU+ cells were found after 2 weeks of labeling (61 ± 9%) but only very few after 10 weeks (20 ± 15%; Figure 5). As in mice, omitting BrdU from the drinking water led to no BrdU staining at all (Figure 5).
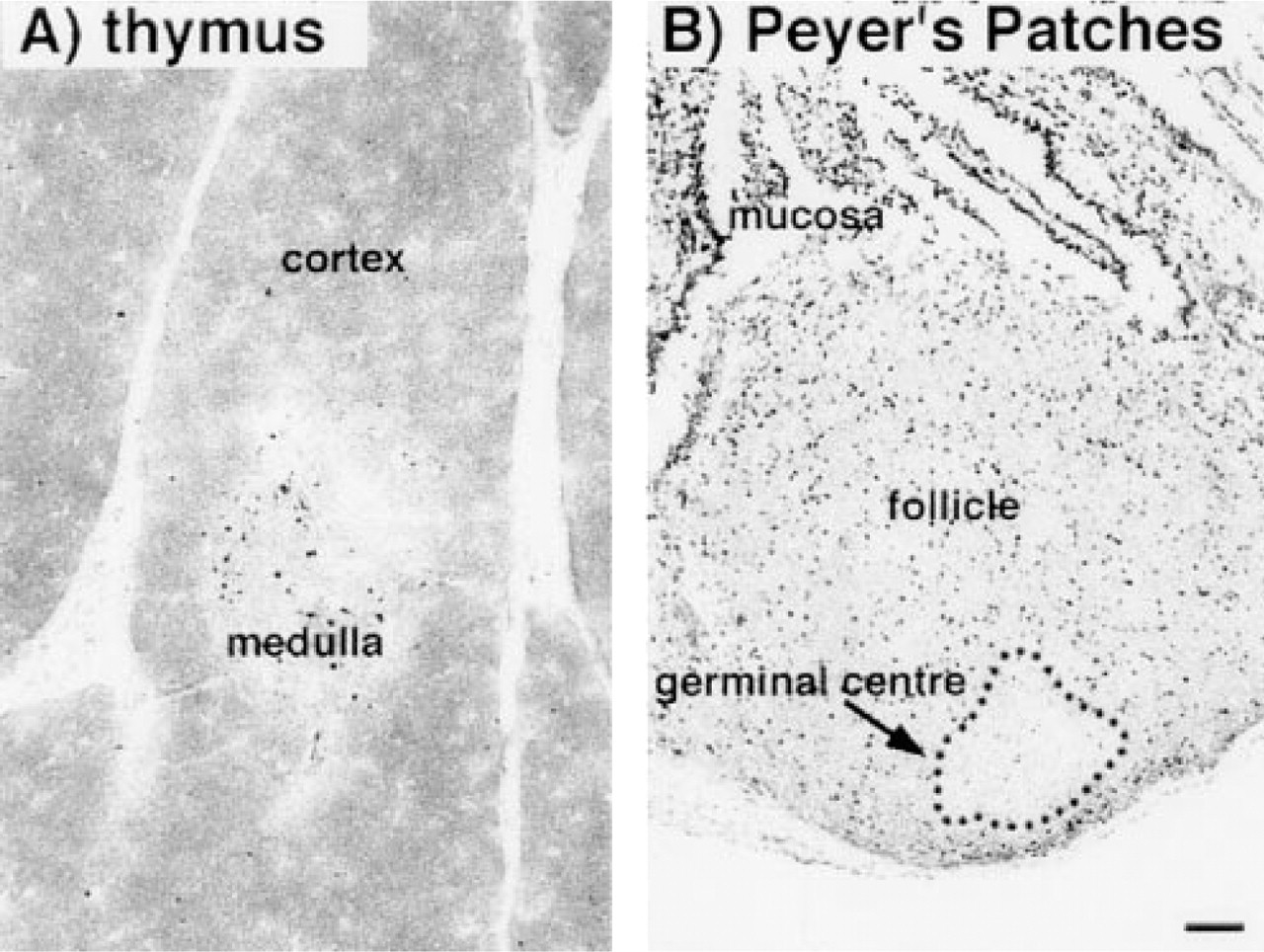
Cryostat sections of rat thymus (
Comparison of body and organ weights and the number of blood leukocytes in control animals and BrdU-treated rats
Comparison of different subpopulations in various organs between control and BrdU-treated animals
aPercentage of positive cells (including BrdU-positive and -negative cells), single data.
bPercentage of positive cells (including BrdU-positive and -negative cells); n = 5; mean ± SD.
cThe antibody staining CD4+ T-cells also detects macrophages and monocytes because they express the CD4 antigen. In addition, apart from CD8+ T-cells, many NK cells are also CD8+. This is why the percentage of CD4+ and CD8+ cells is higher than that of total T-cells.
dThe percentage of macrophages is so low because the tissue was disrupted by mincing it.
eThe percentage of granulocytes is so low because blood cells were isolated on a Ficoll gradient.
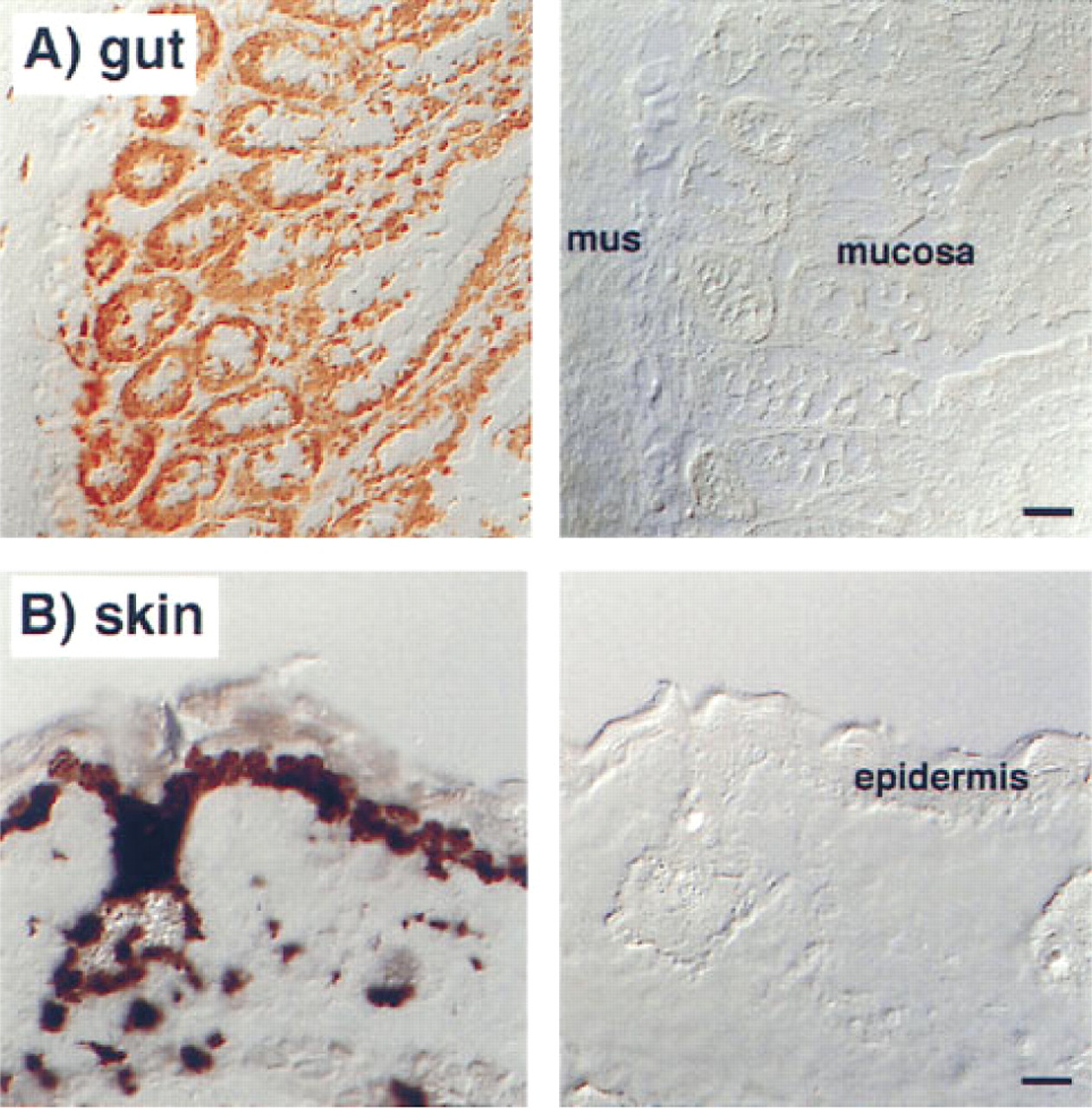
Cryostat sections of the small intestine (
Time-course Analysis Showed that the Number of BrdU+ Cells First Increased but then Decreased, Although the BrdU Uptake Was Constant
When the numbers of BrdU+ cells were studied in LEW rats at several time points in various organs during a 12-week labeling period, it became obvious that the number of BrdU+ cells did not increase constantly (Figure 6). The most dramatic changes were observed in the bone marrow (Figure 6 B). After 2 weeks of labeling, about 70% of all bone marrow cells were BrdU+. Then the number declined to reach a level of about 10% after 12 weeks of BrdU labeling. An increase and a subsequent decrease in the numbers of BrdU+ cells were also seen for blood, spleen, and lymph nodes, although less pronounced (Figures 6C and 6D). When the staining intensity of the gut and skin epithelium (which is proportional to the amount of incorporated BrdU) was investigated over time, similar changes were observed (data not shown). Surprisingly, during the entire time the BrdU+ uptake of the rats was constant (Figure 6 A).
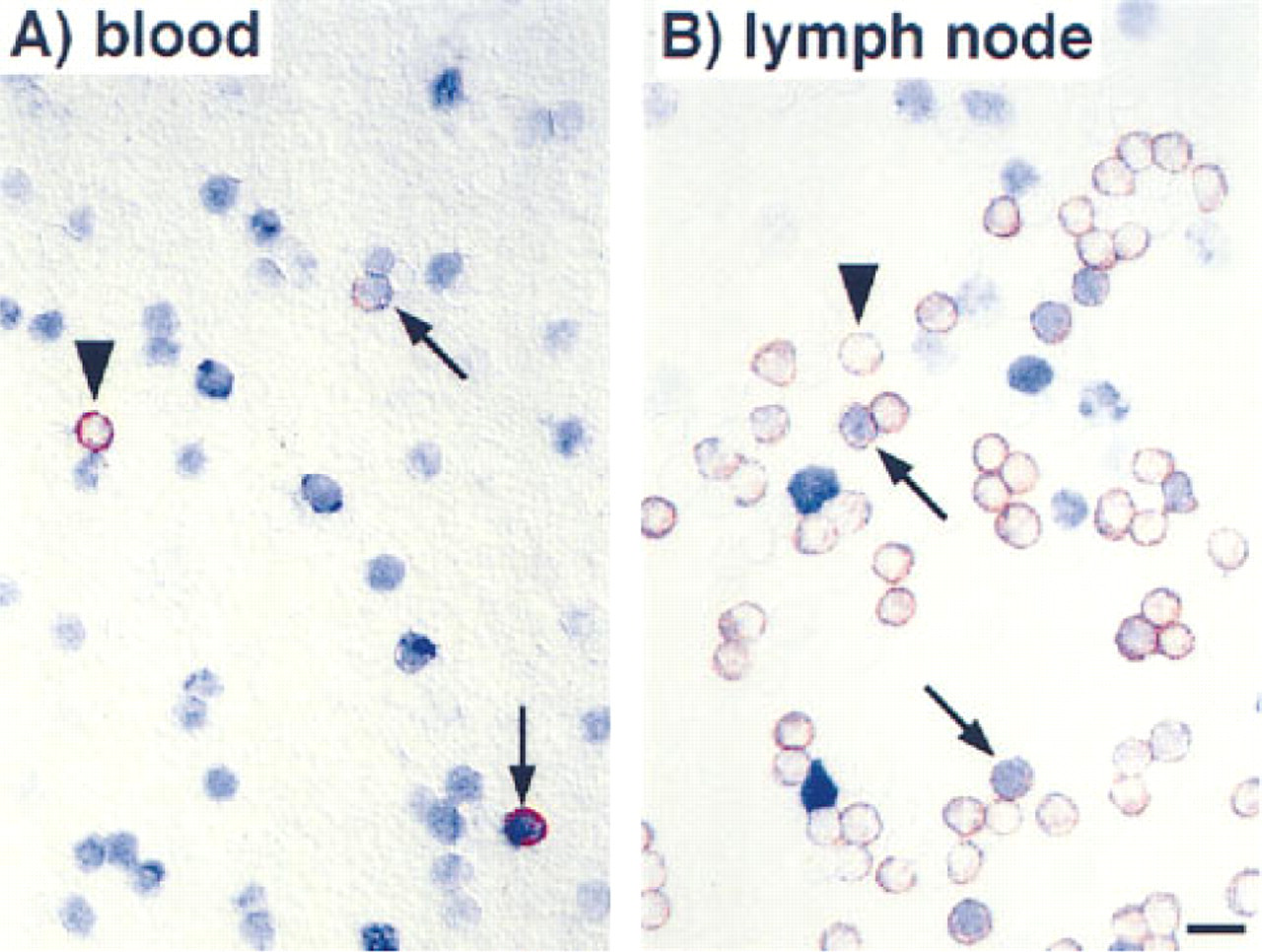
Cytology of rat blood (
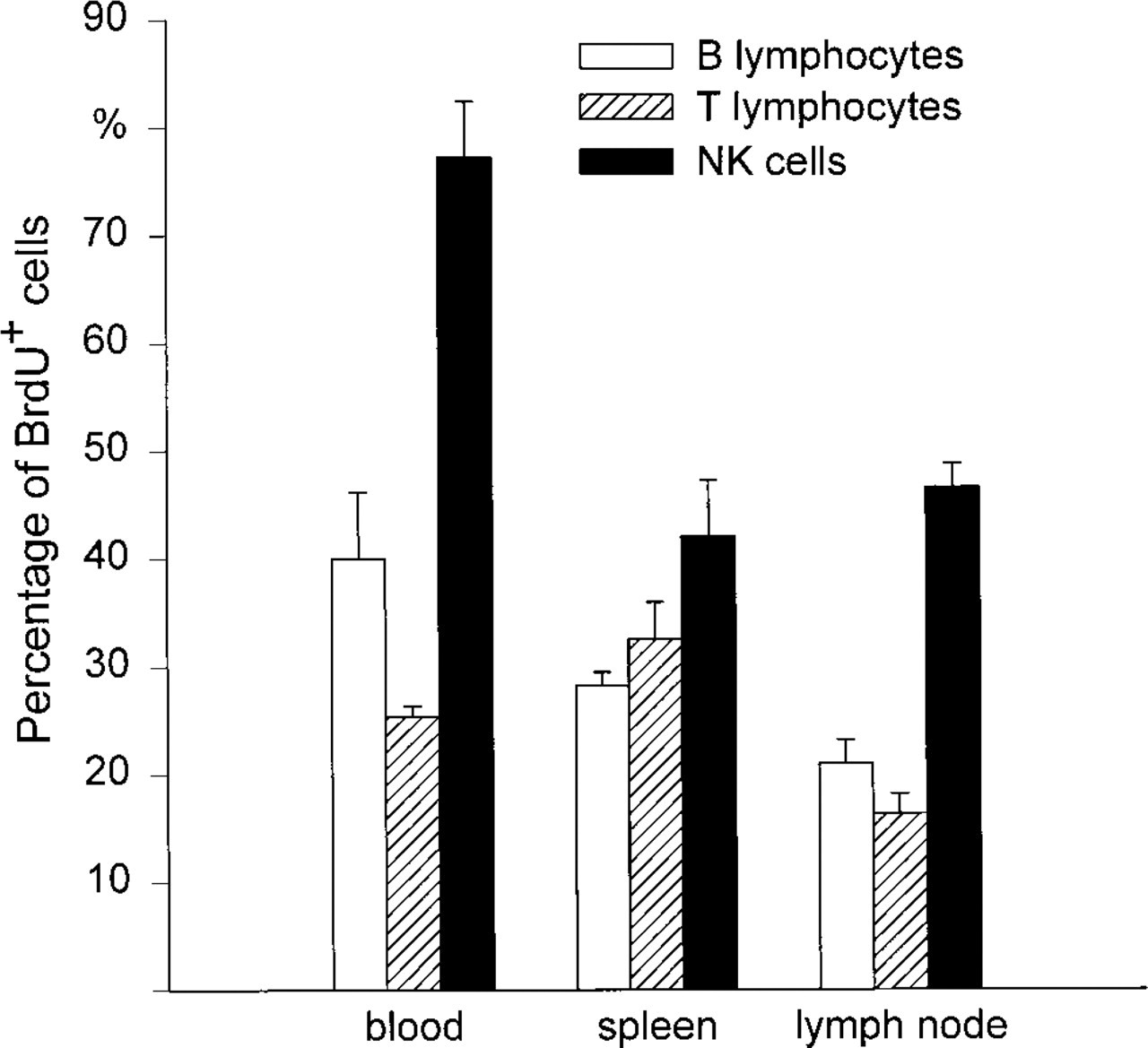
Percentage of BrdU+ cells among B- and T-lymphocytes and NK cells in blood, spleen, and mesenteric lymph nodes 12 weeks after oral BrdU administration. Means ± SD are indicated (n = 5).
Discussion
BrdU is commonly used to label proliferating cells during S-phase of the cell cycle (Gratzner 1982; Goz 1978). In this study, BrdU was administered orally for 12 weeks to study the lifespan of immune cell populations. Each day the rats received about 20 mg BrdU, which is in the range reported by other groups (e.g., Chan and MacLennan 1993). As expected, after this period the epithelium of the gut and the skin was completely labeled with BrdU. However, unexpectedly few cells in thymus, bone marrow, and germinal centers of Peyer's patches were labeled with BrdU, although most of them should have incorporated BrdU during the 12 weeks of administration. This indicates that not all cells passing through the S-phase of the cell cycle during the time of BrdU administration had been labeled, and clearly shows that the data obtained on the lifespan of B- and T-lymphocytes and natural killer cells in blood, spleen, and lymph nodes cannot be interpreted correctly.
Because the denaturation protocol is critical in revealing incorporated BrdU (Hayashi et al. 1988; Moran et al. 1985), and because the DNA of proliferating cells is less sensitive to denaturation (Darzynkiewicz et al. 1979), in a first step the possibility was excluded that the few BrdU+ cells in thymus, bone marrow, and Peyer's patches might be due to a technique inappropriate for detecting incorporated BrdU. When our technique was applied to the thymus and bone marrow of Balb/c mice orally administered the same concentration of BrdU, after a short period over 90% of the cells were BrdU+, and remained BrdU+, in agreement with the data reported by several groups (Tough and Sprent 1994; Deenen and Kroese 1993; Schittek et al. 1991).
The few BrdU+ cells in thymus, bone marrow, and the germinal centers of Peyer's patches are also not due to gross toxic effects of BrdU. Histological examination showed a normal architecture of thymus and Peyer's patches (Figure 1), and cytological examination a normal cellular composition of the bone marrow (Table 3). In addition, the numbers of granulocytes in the blood, which are predominantly produced in the bone marrow, were not affected by BrdU administration (Table 2). The unchanged body and organ weights, and the normal cellular composition of blood, spleen, and lymph nodes, also argue against toxic effects of BrdU in our experiments (Table 2). Even toxic effects of BrdU resulting in the death of only a small number of cells should be picked up by the parameters studied because, owing to the long BrdU administration period, such effects would accumulate with time and most likely would result in detectable damage. However, by largely unknown mechanisms (Tapscott et al. 1989), BrdU treatment can lead to functional alterations such as an increase in lung compliance (Nagai et al. 1993), a higher invasiveness of melanoma cells (Thomas et al. 1993), an arrest in the G2-phase of the cell cycle (Aarnás et al. 1990), or damage to lymphocyte precursors of the bone marrow (McGregor 1969), which cannot be completely ruled out by the methods used. In addition, this study does not assess the LD50 of BrdU.
Because our technique is as sensitive as techniques described by other groups (Tough and Sprent 1994; Schittek et al. 1991; Rocha et al. 1990) and because the dosage of BrdU administered is not toxic, the low numbers of BrdU+ cells in thymus, bone marrow, and germinal centers of Peyer's patches must be due to other reasons. Analysis of the time course of the number of BrdU+ cells during 12 weeks of BrdU administration showed that the number of BrdU+ cells in the bone marrow increased in the first 2 weeks to about 70% and then declined to about 10% during the remaining 10 weeks. An increase and a decline in the numbers of BrdU+ cells were also seen in blood, spleen and lymph nodes, although not as pronounced as in the bone marrow. This strongly suggests that the availability of BrdU changed during the experiment, reaching at some point a level that was too low to be detected. Because the amount of BrdU taken up by the animals stayed constant during the entire administration period, at least two mechanisms could lead to a lower level of BrdU in the circulation. The enterocytes might alter their absorption rate in response to chronic BrdU application and thus reduce the amount of circulating BrdU available for incorporation. However, this possibility cannot be the main cause, since the keratinocytes of the epidermis are all labeled after 12 weeks of BrdU administration, indicating that BrdU is reaching the circulation via the gut during the entire period in sufficient amounts. Another possibility could be a change in the metabolism of BrdU during the 12 weeks of administration, leading to a lower level of BrdU in the circulation. It has long been known that BrdU is rapidly dehalogenated in the liver and that only a small fraction is available for incorporation into the DNA (Kriss et al. 1963; Kriss and Revesz 1962). Continuous BrdU application might lead to enzyme induction in the liver, resulting in more efficient degradation of BrdU and thereby reducing the fraction of BrdU that is available for labeling cells in the S-phase of the cell cycle. Because of the competition between applied BrdU and endogenous thymidine (Hume and Thompson 1989), this reduction first becomes visible in organs such as thymus, bone marrow, and germinal centers of Peyer's patches, which show a high proliferation rate and contain huge amounts of endogenous thymidine (Heininger et al. 1971). For example, in human thymocytes 200 times more endogenous thymidine is present than in blood lymphocytes (Cohen et al. 1983). Therefore, it is not surprising that, after oral BrdU administration in mice, the amount of BrdU incorporated by cells of the thymus is much lower than that in the blood or lymph nodes (Tough and Sprent 1994). In addition, bone marrow cells need a higher arterial BrdU concentration for sufficient labeling than cells of peripheral tissues, as shown for rabbits (Stetson et al. 1988). In the LEW rat, it was not possible to overcome this effect by increasing the routinely used BrdU concentration (1 mg/ml) (Kroese et al. 1995; Tough and Sprent 1994; Förster and Rajewsky 1990; Gray 1988) because the LEW rats refused to drink at a BrdU concentration of 1.5 mg/ml.
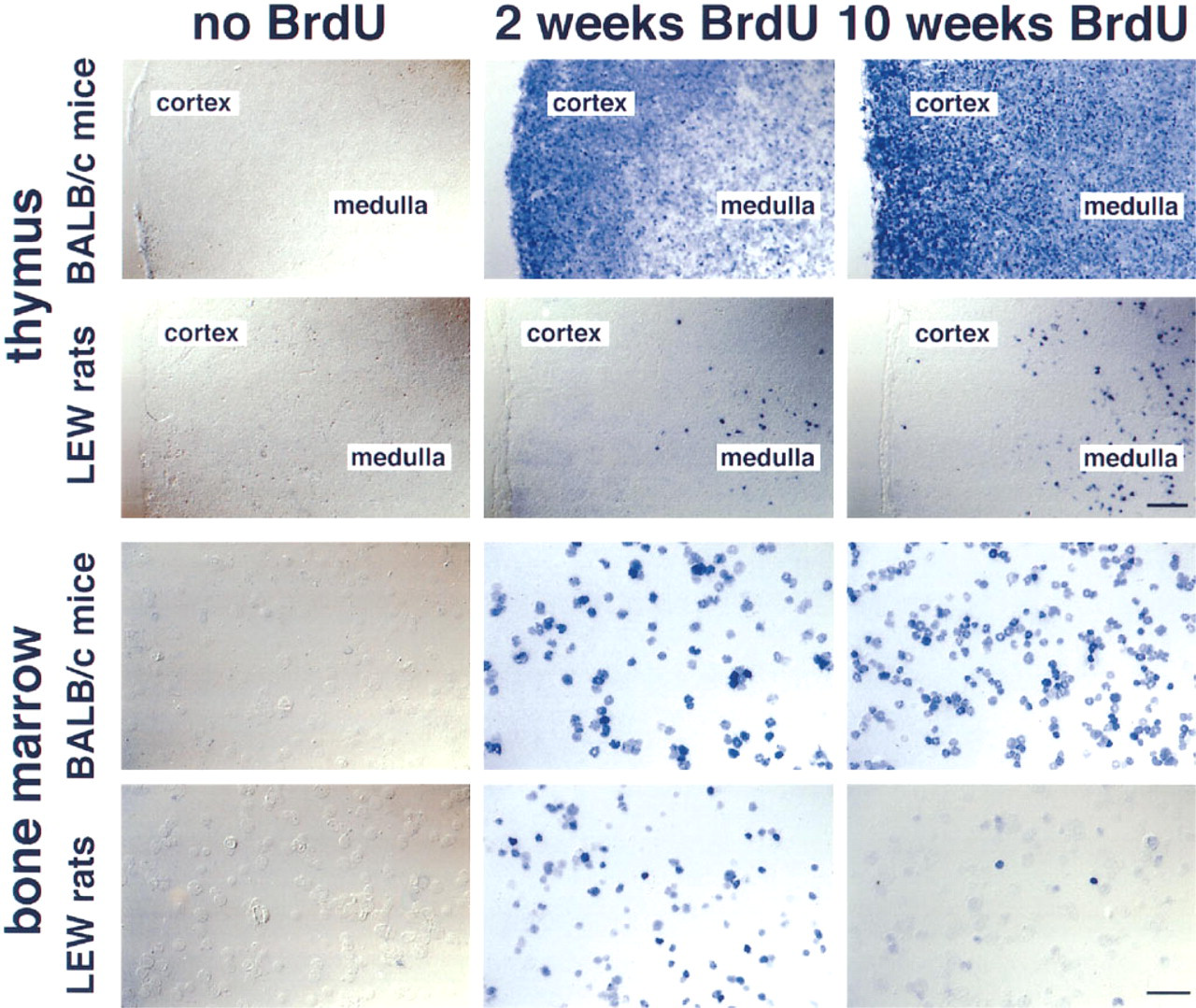
Cryostat sections of thymus and cytospots of bone marrow of Balb/c mice and LEW rats 2 and 10 weeks after oral BrdU administration. Control animals were treated accordingly, with the exception that BrdU was omitted. BrdU+ cells are revealed in blue. In Balb/c mice the expected increase of BrdU+ cells in the thymus is seen, whereas in LEW rats BrdU+ cells are found only in the thymic medulla. In addition, in Balb/c mice the expected increase of BrdU+ cells in the bone marrow is observed, whereas in LEW rats the number of BrdU+ cells first increases and then decreases over time. Alkaline phosphatase-anti-alkaline phosphatase technique, divergence-interference-contrast microscopy. Thymus, bar = 100 μm; bone marrow, bar = 50 μm.
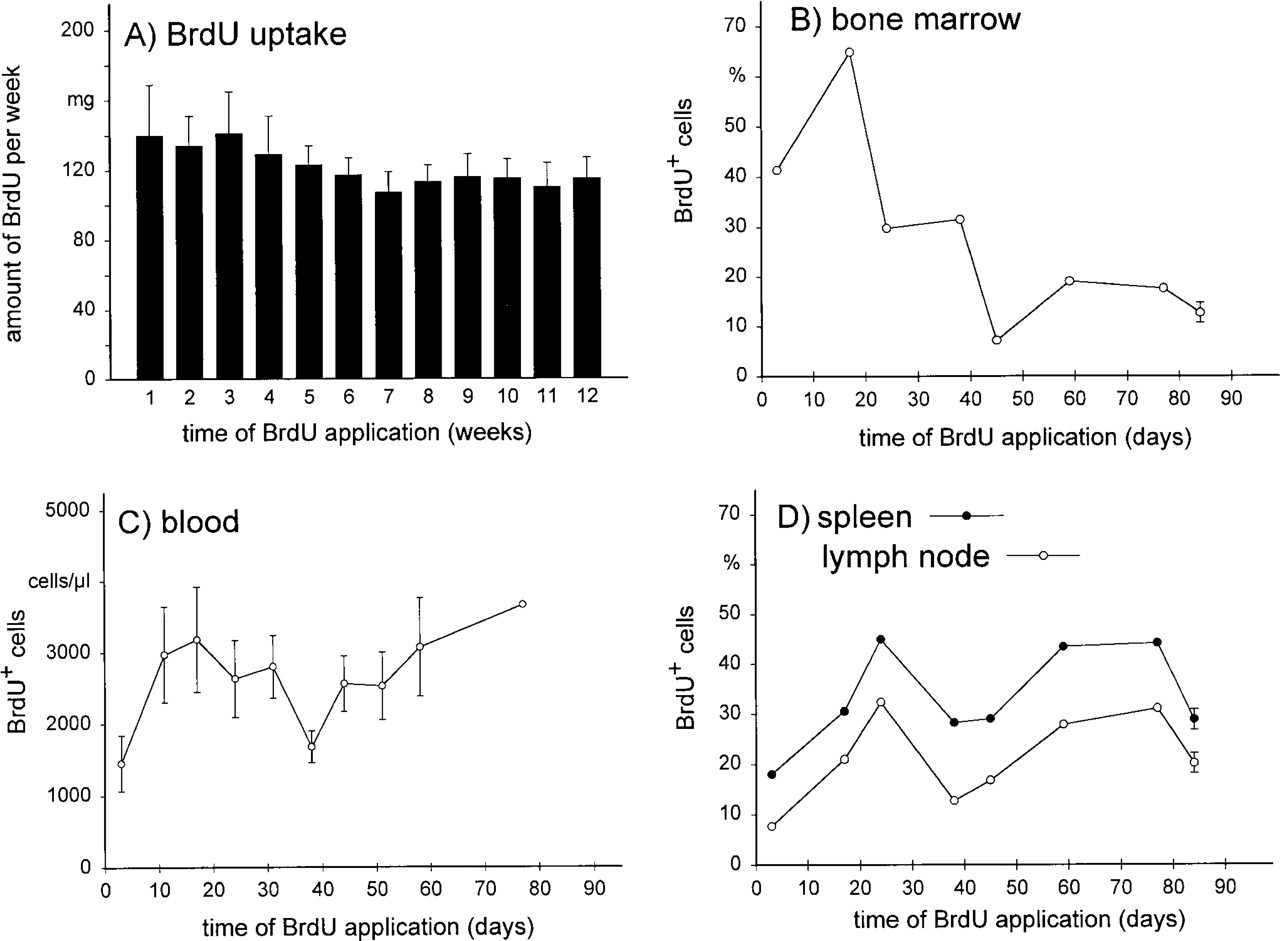
BrdU uptake per week (
One reason that such a change in the metabolism of BrdU has not been described thus far might be the fact that it depends on both the species and the animal strain. When, instead of LEW rats e.g., Balb/c mice (Tough and Sprent 1994) or AO/G rats (Deenen and Kroese 1993) were given BrdU for prolonged time periods, no decrease of labeling efficiency was seen. However, giving B6 mice BrdU under similar conditions leads to the death of these animals after 2 weeks (Rocha et al. 1990). Therefore, administering the same dosage of BrdU in the drinking water (about 1 mg/ml) gives incomplete labeling of proliferating cells in the LEW rat (present study), complete labeling in AO/G rats and Balb/c mice (Tough and Sprent 1994; Deenen and Kroese 1993), and results in death for B6 mice (Rocha et al. 1990). In addition, many studies concentrate on the proliferation of one cell type in one organ and might therefore miss effects such as described above. Taken together, unexpected problems of long-term BrdU treatment, as already described for liver and kidney (Weghorst et al. 1991), also occur when this technique is applied to estimate the lifespan of cell populations of the immune system of the LEW rat. Established BrdU labeling protocols should only very cautiously be applied to other experimental animals, and even a change from one animal strain to the other might cause severe problems.
Footnotes
Acknowledgements
Supported by the Deutsche Forschungsgemeinschaft (Je 201/1-1 and We 1175/4–1).
The generous donations of antibody 3.2.3 by Dr R. Herberman, (Pittsburgh Cancer Institute) and of antibody RP1 by Dr F. Sendo (Yamagata University) are gratefully acknowledged. The technical assistance of Ms P. Schwarz, Ms A. Herden, and Ms. S. Lopez-Kostka, the help by Ms D. Stelte in preparing the figures, and the correction of the English by Ms S. Fryk are deeply appreciated.