
Abstract
Using various sizes and dilutions of hapten-conjugated DNA probes, we compared catalyzed reporter deposition (CARD) to fluorochrome-conjugated antibody layering (immunological method) for amplifying FISH signals. Cosmid and phage probes that contained human DNA inserts of 40
FLUORESCENCE in situ hybridization (FISH) techniques are becoming increasingly useful in both basic research and clinical diagnostics, particularly in the areas of gene mapping, gene expression, tumor biology and clinical cytogenetics (van Ommen et al. 1995; Joos et al. 1994; Lichter et al. 1991; McNeil et al. 1991; Trask 1991; Lichter and Ward 1990; Pinkel et al. 1986).
Recently, advances have been made in the area of signal amplification (Bobrow et al. 1989,1991,1992). The technique, known as catalyzed reporter deposition or CARD, was originally developed for solidphase immunoassays and is based on the enzymatic deposition of biotinylated or fluorochrome-labeled tyramide at the probe site (Bobrow et al. 1989). Over the past several years, CARD has been successfully incorporated into immunohistochemical and FISH procedures (Kerstens et al. 1995; Raap et al. 1995; Adams 1992).
The present study was undertaken with the intent of comparing the CARD amplification method to the indirect immunological method commonly used for detection of FISH probe signals. By using probes of varying sizes and at concentrations lower than those typically applied, the detection capability of these methods was evaluated.
Materials and Methods
Probe Preparation
A cosmid cocktail containing two approximately 40-
Target Chromosome Preparation
Metaphase chromosomes from human peripheral blood lymphocytes were prepared according to standard cytogenetic protocols (Barch 1991). Metaphase cell suspensions were spread onto glass slides and stored at room temperature (RT; if used within 1 week) or at −4C (for up to 3 weeks from preparation). Before in situ hybridization the slides were dehydrated through a series of ethanol washes, air-dried, and denatured at 70C in 70% formamide in 2 X SSC.
In Situ Hybridization
The probes, in hybridization buffer, were applied to the chromosomal spreads and incubated overnight at 37C. They were then subjected to a posthybridization stringency wash (2 X SSC at 72C), followed by a brief wash in PBS (pH 7.4) containing 0.1% NP-40 (PBD) before signal amplification.
Detection and Amplification
Both immunological and CARD methods were used for the detection and amplification of the FISH signals. The immunological method involved incubation of the hybridized slides with a rhodamine-conjugated sheep anti-digoxigenin antibody (sh α dig-rhd) (Boehringer-Mannheim; Indianapolis, IN), followed by rabbit anti-sheep IgG (rb α sh) (Jackson ImmunoResearch; West Grove, PA), and finally rhodamine-conjugated goat anti-rabbit antibody (gt α rb-rhd) (Jackson ImmunoResearch), each for 15 min at RT with intervening PBD washes.
Enzymatic amplification of the hybridization signal (CARD) was based on the technique developed by Bobrow et al. (1989). Immediately before amplification the slides were washed in Tris-buffered saline (TBSB) containing 10% horse serum (Sigma Chemicals; St Louis, MO) and 0.05% tyramine HCl (Sigma) for 1–2 hr at RT. Slides were then subjected to a series of antibody incubations as follows: (a) horseradish peroxidase-conjugated sheep anti-digoxigenin (sh α dig-hrp); (Boehringer-Mannheim) for periods ranging from 15 min to overnight; (b) biotin-conjugated donkey anti-sheep IgG (Jackson ImmunoResearch) for 15 min; (c) horseradish peroxidase-conjugated streptavidin (SA-Hrp) (DuPont NEN; Wilmington, DE) for 15 min; and (d) fluorochrome-conjugated tyramide (DuPont NEN) for 5–15 min. Each antibody incubation was followed by a short wash in TBSB.
For experiments involving the biotin-labeled phage probe, the detection and amplification procedures used different reagents, but the general scheme remained the same. For the immunological method, amplification consisted of a cyanine dye conjugated to streptavidin (SA-Cy3) (Sigma) as the first layer, followed by a biotin-conjugated goat anti-streptavidin (gt α SA-biotin) (Vector Laboratories; Burlingame, CA) and ending with another layer of SA-Cy3. In performing the enzymatic amplification (CARD method), the layers consisted of SA-Hrp, gt α SA-biotin, SA-Hrp, and fluorochrome-conjugated tyramide. As previously described, the preparations were washed briefly between administration of each layer.
Visualization and Scoring of Hybridization Signals
After the final amplification layer was reached for either the immunological or the CARD method, slides were washed briefly in Tris-buffered saline containing 0.05% Tween-20 (TBST) and mounted in an anti-fade mounting medium (p-phenylenediamine, pH 8) containing 50 ng/ml 4',6-diamidino-2-phenylindole hydrochloride (DAPI) as a counterstain to visualize the chromosomes.
Hybridization signals were detected using an Axioskop equipped with a triple pass (DAPI, rhodamine, FITC) filter (Chroma Technology; Brattleboro, VT). When successfully hybridized, the cosmid cocktail revealed signals in both the proximal and distal long arms of both no. 15 chromosomes in each cell, corresponding to the 15q11.2 and 15q22 loci, respectively. For the phage probe, successful hybridization revealed signals only in the proximal long arm of the no. 15 chromosomes, corresponding to the 15q11.2 locus. Hybridization reactions were evaluated for the presence and intensity of these locus-specific signals and also for the presence and intensity of nonspecific fluorescent background signals.
Results and Discussion
Table 1 and Figures 1A-1E show the results from the comparison of the two detection methods used in this study. For the cosmid probes at standard probe concentrations of 10 ng/μl, the two detection methods were comparable in signal intensity and background staining. Figure 1 A is representative of the fluorescence signals routinely obtained from the immunological detection system with a probe concentration of 10 ng/μl. Note the appearance of the strong signals representing the 15q11.2 and 15q22 cosmids. With this method, strong signal intensities for the cosmids were maintained at a probe concentration of 5 ng/μl. However, diluting the probe further resulted in a loss of signal intensity (Table 1).
Comparison of FISH signal detection using two amplification methods
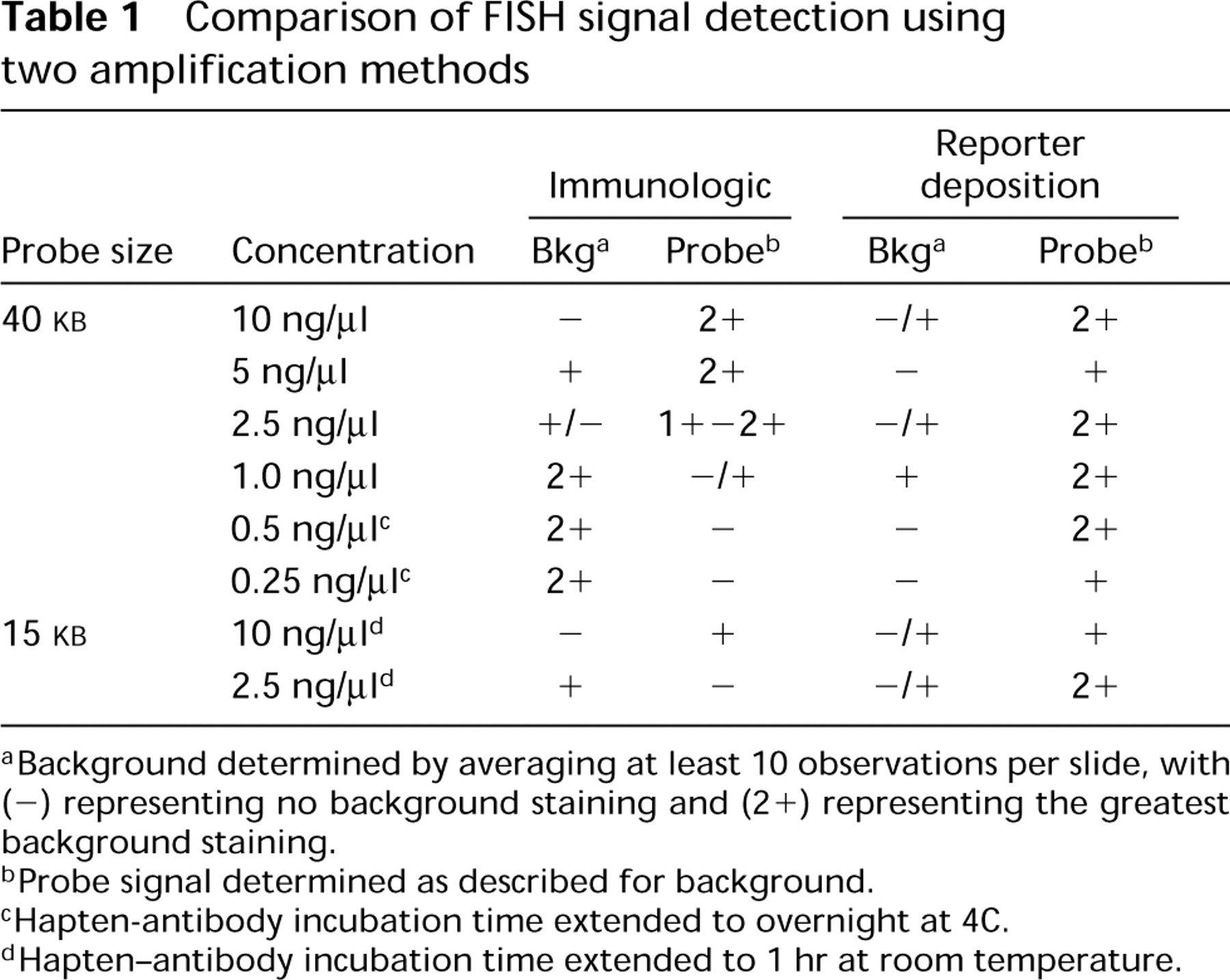
aBackground determined by averaging at least 10 observations per slide, with (-) representing no background staining and (2+) representing the greatest background staining.
bProbe signal determined as described for background.
cHapten-antibody incubation time extended to overnight at 4C.
dHapten-antibody incubation time extended to 1 hr at room temperature.
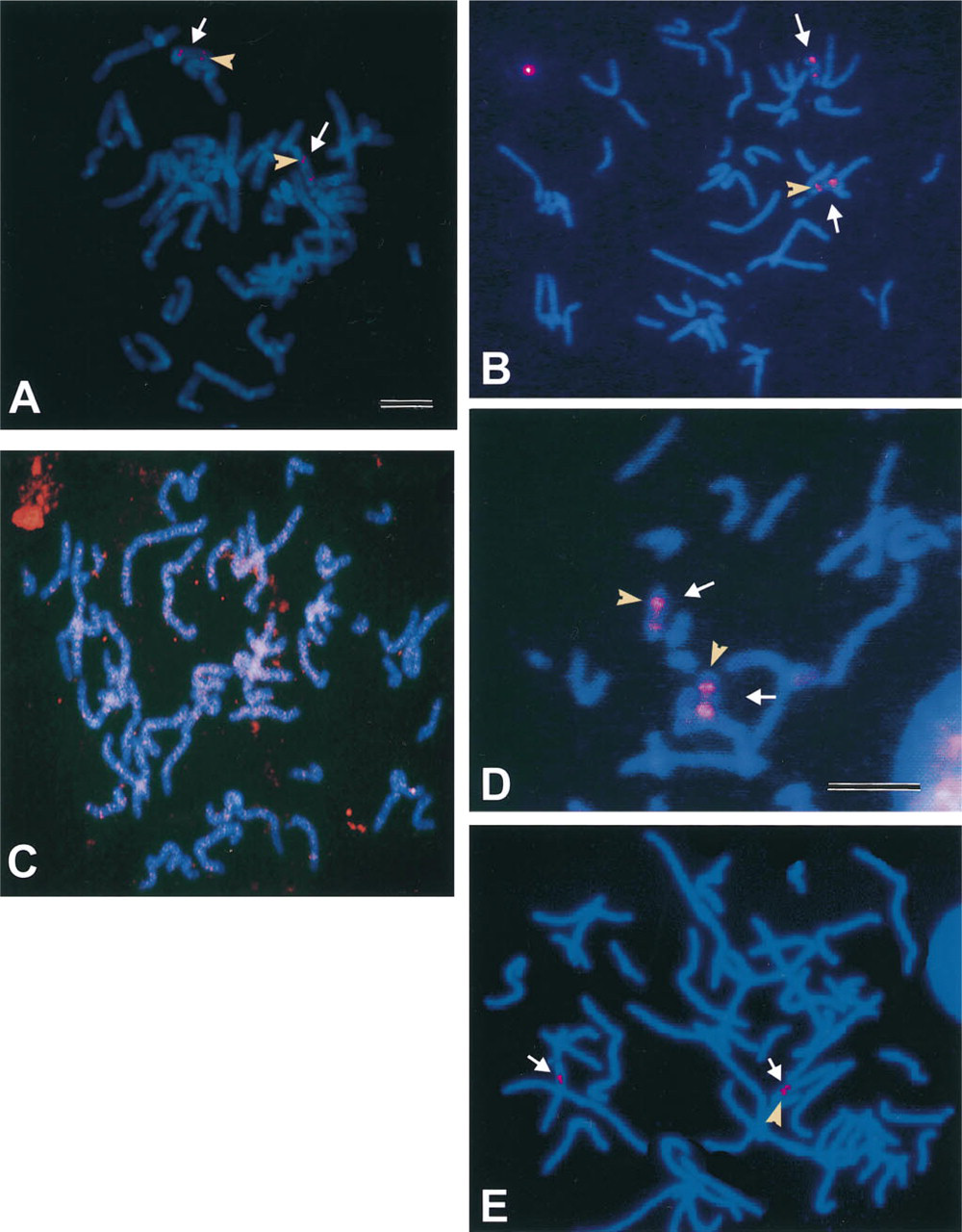
Comparison of fluorescence in situ hybridization signals obtained from two amplification methods using a 40-
In contrast, the CARD amplification method resulted in intense probe signals even at probe concentrations as low as 0.25 ng/μl (Table 1) while maintaining the resolution of these signals. Figure 1 B shows the results obtained using the CARD method and a probe concentration of 1.0 ng/μl. By comparing Figures 1A and 1B, it is clear that the CARD method improves the detection capabilities of in situ hybridization with diluted probe. Even though the probe has been diluted 10-fold, the intensity of the signal is equal to or greater than that seen with the immunological detection method. Figure 1 D further emphasizes the capabilities of the CARD method by showing the intense signal obtained even at a probe dilution of 1:40 (0.25 ng/μl). An increase in sensitivity was accomplished by overnight incubation with the sh α dig-rhd antibody followed by subsequent incubation in Tyramide-rhd, but it should be noted that a slight reduction in resolution occurred. The two signals for cosmid 15q11.2 in 1D are distinguishable but appear fused, owing to an overlap of peripheral fluorescent halos that most likely resulted from a slightly prolonged incubation in the tyramide signal amplification step. The temporal duration of the tyramide amplification step is critical because (a) excessive incubation may lead to enlarged areas of signal, leading to a decrease in resolution although sensitivity may be increased, and (b) if this step is not well-monitored, over-incubation can lead to substantial increase in background that can interfere with signal interpretation.
With both the immunological and the CARD method, nonspecific fluorescence signals became a problem at very low probe concentrations or extended hapten-antibody incubation times (4 hr to overnight). Addition of large polymeric substances or normal serum to the buffers used in immunofluorescence methods is known to improve spatial resolution (Van Gijlswilk et al. 1996; Van Noorden and Vogels 1989) and decrease background staining (Jeppesen 1994) of these techniques respectively. Improvement in signal detection and reduction in background staining was obtained when normal horse serum (10%) and unlabeled tyramine-HCl (0.05%) were added to the buffers/wash solutions. These efforts to reduce the background staining clearly helped to facilitate detection of the probe signal with the CARD amplification technique but had little effect on the immunological amplification technique, as seen in Table 1 and by comparing Figures 1B and 1C. Figure 1 B represents the CARD amplification of the FISH signal of a 40-
Perhaps the most exciting prospect of using CARD amplification techniques can be found in their ability to detect in situ hybridization signals from smaller probes. The CARD amplification technique was useful in detecting the fluorescence signals resulting from hybridization with the smaller 15-
Dilution of the probe beyond the standard concentrations currently used in most laboratories resulted in detectable signals with the CARD method, but it should be noted that, with CARD, the intensity of the signals could be enhanced by extending the exposure time between hapten and antibody. Although, as noted previously, this created some background noise, the specific signal was still distinguishable. In contrast, extending the exposure time between hapten and antibody when the immunological protocol was used increased background without concomitant improvement of either sensitivity or resolution of the probe signal.
The results of this study further emphasize the potential usefulness of the CARD amplification technique in detecting FISH signals. The demonstrated ability of the CARD amplification technique to detect probes present at very low concentrations while maintaining signal intensity and resolution may be particularly valuable in applications with both small probes and targets. In this regard, work is now being conducted in which the CARD amplification technique is successfully being used to detect FISH signals in the nuclei of parasitic protozoan Giardia (data not shown). Given the fact that a single nucleus in Giardia is approximately 1–1.5 μm in diameter (Erlandsen and Rasch 1994), the ability to detect a signal on a single chromosome within this nucleus suggests that the value of this technique has not yet been fully realized.
Footnotes
Acknowledgements
Supported in part by US Environmental Protection Agency (USEPA) Cooperative Agreement CR816637 and by USEPA grant R82140401.