
Abstract
Immunocytochemical detection of bromodeoxyuridine (BrdU) labeling can be hampered by low BrdU incorporation levels. We describe here an amplification method for weak BrdU immunosignals. The tyramide signal amplification method based on catalyzed reporter deposition (CARD) uses fluorescein-labeled tyramide as a substrate for horseradish peroxidase. The enzyme catalyzes the formation of highly reactive tyramide radicals with a very short half-life, resulting in the binding of fluorescein-conjugated tyramide only at the site of the enzymatic reaction. MCF-7 cells were grown in vitro in medium containing charcoal-stripped fetal bovine serum supplemented by growth factors. Under these culture conditions, the BrdU immunosignal was hard to detect but could be enhanced specifically by the tyramide signal amplification system, resulting in clear-cut differences between BrdU-negative and BrdU-positive cells. This enabled rapid and objective quantification of the BrdU labeling index without the risk of underestimating the number of cells in S-phase. Therefore, this amplification of BrdU immunosignals might also prove valuable for in vivo cancer prognosis, cell kinetics studies, and computer-assisted image analyses.
A
However, the immunocytochemical detection of BrdU labeling can be hampered by low BrdU incorporation levels. Low incorporation levels of labeled nucleotides can occur as a consequence of altered cell growth rate (Aakvaag et al. 1990), which can be the result of the culture conditions used, such as the use of charcoal-stripped serum instead of untreated serum (Roman et al. 1993). This would result in weakly immunostained nuclei. In such cases, amplification of the BrdU signal is necessary to eliminate the considerable risk of underestimating the number of cells in S-phase and to ensure an objective quantification of the BrdU labeling index.
Catalyzed reporter deposition (CARD) is a signal amplification method that has been described by Bobrow et al. (1989,1991). The amplification is achieved by biotin- or fluorescein-labeled tyramides that act as substrates for horseradish peroxidase. The horseradish peroxidase catalyzes the formation of highly reactive tyramide radicals that bind covalently to tyrosine residues (Bobrow et al. 1989,1991). Because of the extremely short half-life of the tyramide radicals, only tyrosine residues in the direct surroundings of the horseradish peroxidase will bind tyramide (Bobrow et al. 1989,1991). The biotin-labeled tyramide can then be visualized by fluorochrome- or enzyme-labeled streptavidin.
Although originally developed for ELISA, this technique has been adapted for immunohistochemistry on tissue sections (Merz et al. 1995; Berghorn et al. 1994; Adams 1992) and for in situ hybridization (Kerstens et al. 1995; Raap et al. 1995).
In this study, MCF-7 human breast cancer cells were used, grown in vitro in medium containing charcoal-stripped fetal bovine serum supplemented with growth factors. Under these culture conditions, these cells are known to have an increased cell doubling time (Roman et al. 1993; Aakvaag et al. 1990; Berthois et al. 1986), resulting in low BrdU incorporation levels and thus weak BrdU immunosignals. Fluorescein-labeled tyramide (Bobrow et al. 1991) was used as a substrate for horseradish peroxidase to amplify specifically the weak immunocytochemical signal of BrdU labeling. The tyramide signal amplification method enabled a rapid and objective quantification of the BrdU labeling index without the risk of underestimating the number of cells in S-phase.
Materials and Methods
Materials
Dulbecco's modified Eagle's medium (DMEM) with 4.5 g/liter glucose, phenol red-free DMEM with 1 g/liter glucose and 1 mM sodium pyruvate, fetal bovine serum (FBS),
Dextran T500 was obtained from Pharmacia (Uppsala, Sweden) and activated charcoal from Sigma (St Louis, MO). Charcoal-stripped FBS was prepared by treating FBS with dextran-coated charcoal (DCC-FBS), according to the protocol of Horwitz and McGuire (1978).
Monoclonal antibody to BrdU (clone BU-1) containing nuclease activity was purchased from Amersham (Poole, UK). Secondary biotinylated goat anti-mouse IgG was obtained from DAKO A/S (Prosan; Ghent, Belgium) and secondary fluorescein isothiocyanate (FITC)-labeled goat anti-mouse IgG from Southern Biotechnology Associates (Birmingham, AL). The Tyramide Signal Amplification (TSA)-Direct kit (Green) was obtained from DuPont NEN Products (Brussels, Belgium) and contained blocking reagent, horseradish peroxidase-conjugated streptavidin, 2 X concentrated amplification diluent, and fluorescein-labeled tyramide.
Cell Culture
Routinely, MCF-7 human mammary carcinoma cells, purchased from the American Type Culture Collection (Rockville, MD), were cultured in DMEM with 4.5 g/liter glucose supplemented by 10% FBS, 2 mM
For the growth experiments, cells were cultured for 6 days in phenol red-free DMEM containing 5% DCC-FBS, 4.5 g/liter glucose, 2 mM
Control cells, growing in medium containing 10% untreated FBS, were seeded from stock cultures onto PLL-coated Chamber Slides at a concentration of 2500 cells/chamber. Cells were grown under these conditions for 7 days, with medium changes 3 and 6 days after seeding.
BrdU Labeling and Immunodetection
Cells grown for 7 days under the different culture conditions described above were incubated for 2.5 hr with an excess (100 μM) of BrdU at 37C in an incubator. Thereafter, cells were washed twice with serum-free culture medium and fixed in methanol (5 min, −2C) and acetone (10 sec, −2C), and air-dried. Cells were then ready to use for BrdU detection according to the following Protocols A or B.
Protocol A: BrdU Detection by Conventional Immunofluorescence Staining
1. Slides were blocked in 5% normal goat serum in 10 mM PBS (pH 7.4) for 1 hr in a dark, moist chamber at room temperature (RT).
2. Slides were rinsed three times for 10 min with PBS at RT.
3. Slides were incubated with monoclonal antibody against BrdU containing nuclease (diluted 1:5 in PBS) for 5 hr at RT in a dark, moist chamber. Omission of the primary antibody did not reveal any signal. Different incubation times (1 hr, 3 hr, 4 hr, 5 hr, and 16 hr) and antibody dilutions (1:2, 1:5, 1:8, 1:10) were tested. The best results were obtained with an incubation time of 5 hr and an antibody dilution of 1:5.
4. Slides were rinsed three times for 10 min with PBS at RT and stored overnight.
5. Slides were incubated with FITC-labeled goat anti-mouse IgG (diluted 1:60 in PBS) for 1 hr at RT in a dark, moist chamber.
6. Slides were rinsed three times for 10 min with PBS at RT.
7. Slides were postfixed for 10 min in methanol at RT.
8. Slides were postfixed for 10 min in 4% paraformaldehyde in PBS at RT.
9. Slides were rinsed three times for 10 min with PBS at RT.
10. Slides were rinsed three times for 10 min with distilled water at RT.
11. Slides were air-dried and mounted in Mowiol containing propidium iodide (PI, 2 μg/ml final concentration) and RNAse (20 μg/ml final concentration) to stain nuclei.
Protocol B: BrdU Detection by Tyramide Signal Amplification
1. Endogeneous peroxidase activity was blocked by incubating slides in 0.03% H2O2 diluted in PBS for 30 min at room temperature (RT).
2. Slides were washed twice for 5 min in PBS at RT.
3. Slides were blocked in 5% normal goat serum in 10 mM PBS (pH 7.4) for 1 hr at RT in a dark moist chamber.
4. Slides were rinsed three times for 10 min with PBS at RT.
5. Slides were incubated with monoclonal antibody against BrdU containing nuclease (diluted 1:8 in PBS) for 5 hr at RT in a dark, moist chamber. Omission of the primary antibody did not reveal any signal. Different incubation times (1 hr, 3 hr, 4 hr, and 5 hr) and antibody dilutions (1:2, 1:5, 1:8, 1:10) were tested. The best results were obtained with an incubation time of 5 hr and an antibody dilution of 1:8.
6. Slides were rinsed three times for 10 min with PBS at RT and stored overnight.
7. Slides were incubated with biotinylated goat anti-mouse IgG (diluted 1:300 in PBS) for 30 min at RT in a dark, moist chamber.
8. Slides were rinsed three times for 5 min with PBS at RT.
9. Slides were blocked with 0.5% DuPont Blocking Reagent in 0.1 M Tris-HCl (pH 7.5) containing 0.15 M NaCl (TBS) for 30 min at RT in a dark, moist chamber.
10. Slides were incubated for 30 min with streptavidin-HRP diluted 1:500 in TBS containing 0.5% DuPont Blocking Reagent at RT in a dark, moist chamber.
11. Slides were rinsed three times for 5 min with TBS containing 0.05% Tween 20 (TBST) by shaking at RT.
12. Slides were incubated with fluorescein-labeled tyramide diluted 1:50 in 1 X amplification buffer for 10 min at RT in a dark, moist chamber. Different incubation times (1 min, 5 min, and 10 min) and dilutions (1:25 and 1:50) were tested. An incubation of 10 min and a dilution of 1:50 gave the best results.
13. Slides were rinsed three times for 5 min with TBST by shaking.
14. Slides were rinsed three times for 5 min with distilled water at RT.
15. Slides were postfixed in methanol for 5 min at RT.
16. Slides were air-dried and mounted in Mowiol containing PI (2 μg/ml final concentration) and RNAse (20 μg/ml final concentration) to stain nuclei.
The BrdU labeling index, i.e. the percentage of cells in S-phase of the cell cycle, was determined by counting cells under a fluorescence microscope (Axiophot; Zeiss, Oberkocher, Germany) with a dual filter set for simultaneous visualization of both fluorescein and PI. About 800 cells were counted twice for each test condition per experiment. Results are presented as mean ± SEM (n = 3).
Statistical Analysis
Data were analyzed with the two-tailed Mann-Whitney U-test using Stat View II software (Abacus Concepts; Berkeley, CA). Significance was defined at the level of p<0.05.
Results
Control MCF-7 cells, grown in medium containing untreated FBS, showed a BrdU labeling index (Table 1) of 36.8 ± 1.3%, as described previously (Van heusden et al. 1996). On BrdU immunodetection, normal BrdU incorporation levels were observed, which resulted in strongly immunoreactive nuclei.
In contrast, cells grown in medium containing charcoal-stripped FBS supplemented with growth factors showed low BrdU incorporation levels. This was reflected by weak BrdU immunosignals, which were hard to detect using the conventional immunofluorescence staining procedure (Figure 1a). BrdU-positive nuclei were stained faintly and showed a heterogeneous punctate staining pattern (Figure 1a, inset). The BrdU labeling index (Table 1) was 27.9 ± 0.6%, which was statistically different (p<0.05) from that of cells grown in medium containing untreated serum.
The tyramide signal amplification method strongly enhanced the BrdU immunosignal (Figure 1b) of cells grown in medium containing charcoal-stripped serum. The nuclei were now stained intensely and the majority of the cells were homogeneously BrdU-positive (Figure 1b, inset), whereas some cells partially retained their original punctate staining (Figure 1b, inset). On immunofluorescence microscopy, no significant fading of the fluorescein signal was observed, in contrast to the conventional immunofluorescence staining preparations, most probably owing to the intense amplification of the immunosignal.
After visualization with the tyramide signal amplification method, a BrdU labeling index (Table 1) of 26.9 ± 0.9% was obtained which was not statistically different (p = 0.14) from the labeling index (Table 1) of 27.9 ± 0.6% that was obtained with normal immunofluorescence staining. This means that no additional cells became labeled after the tyramide signal amplification.
BrdU labeling index for MCF-7 cells
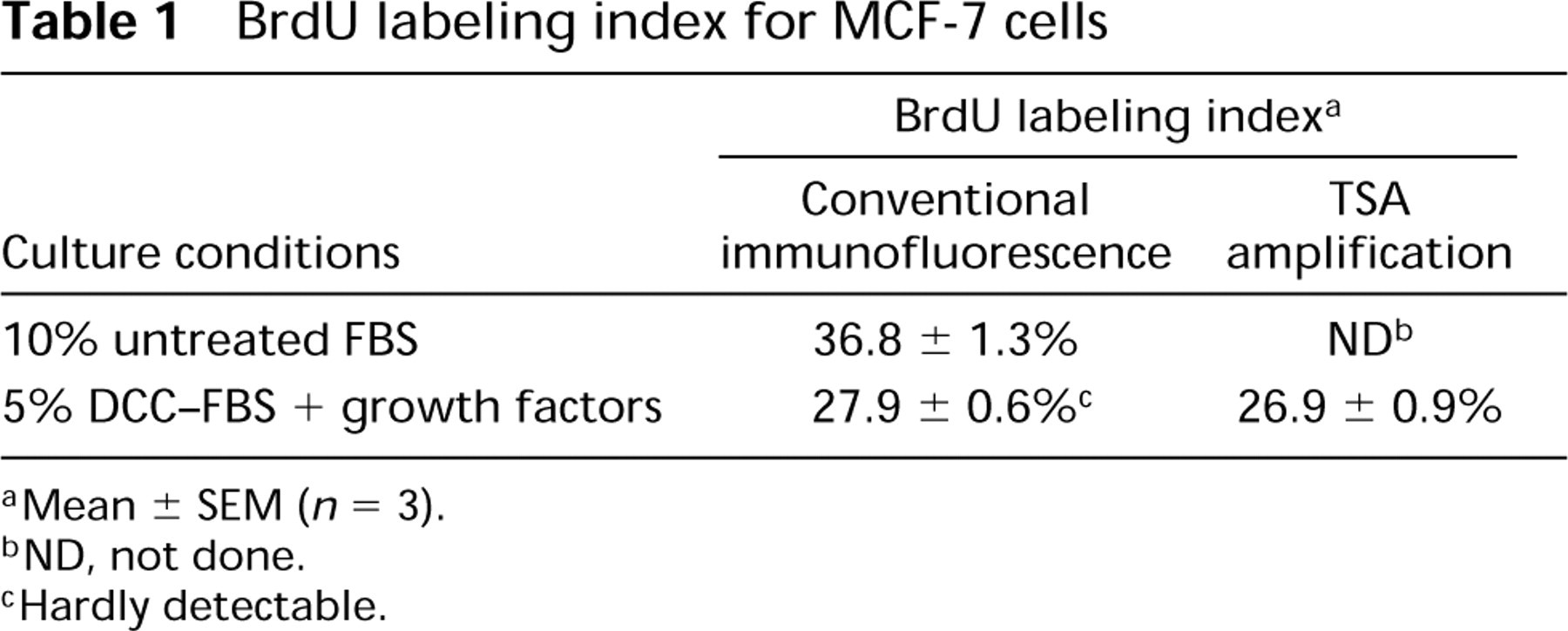
aMean ± SEM (n = 3).
bND, not done.
cHardly detectable.
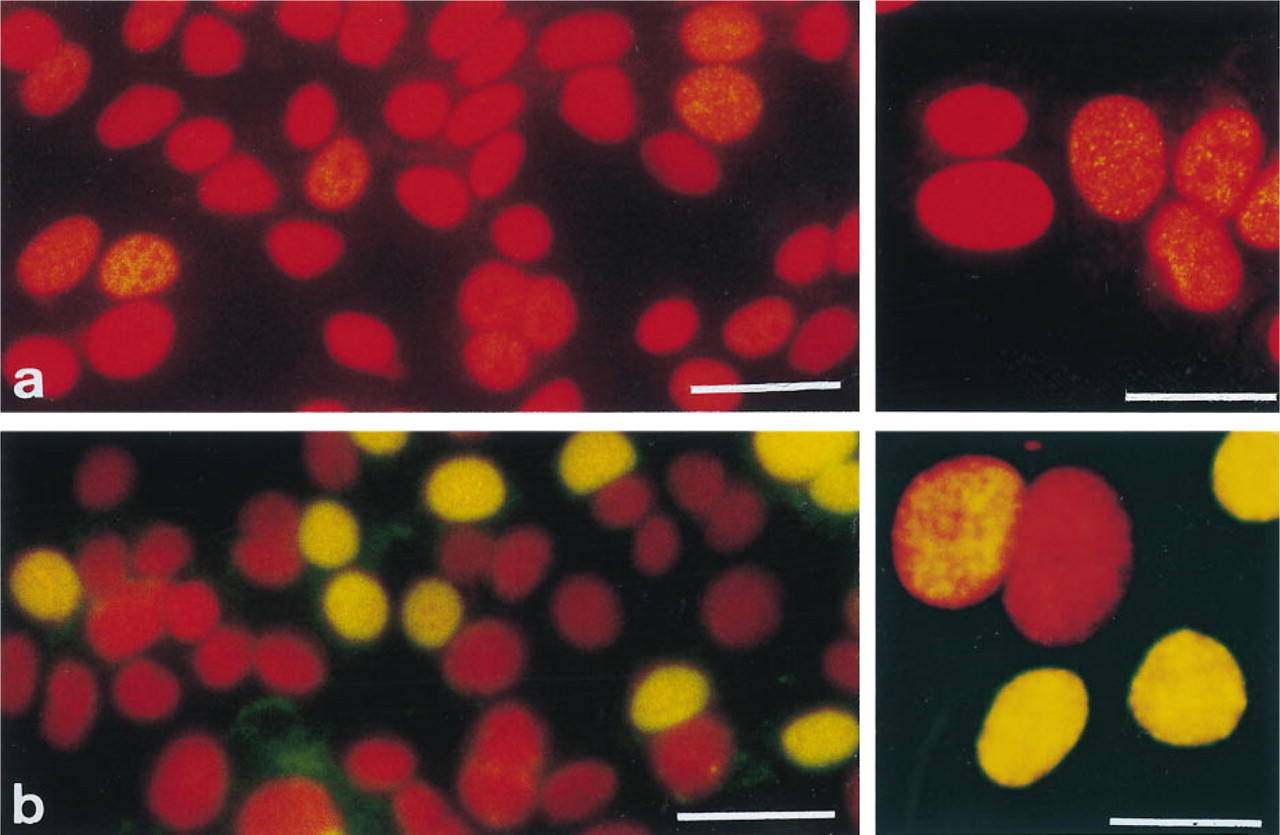
BrdU immunodetection in MCF-7 human breast cancer cells grown in medium containing 5% charcoal-stripped fetal bovine serum supplemented by growth factors. Visualization was achieved by (
Discussion
MCF-7 cells, grown in medium containing charcoal-stripped FBS supplemented by growth factors, showed a decreased BrdU labeling index compared to that of cells grown in medium containing untreated fetal bovine serum. This was not surprising because it is known that MCF-7 cells are estrogen-dependent (Aakvaag et al. 1990; Brooks et al. 1973) and the MCF-7 cells used in this study were cultured essentially in the complete absence of estrogens by the use of phenol red-free medium and charcoal-stripped serum (Berthois et al, 1986; Horwitz and McGuire 1978). Furthermore, MCF-7 cells grown in medium containing charcoal-stripped serum and insulin showed an increased cell doubling time and lowered thymidine incorporation in the absence of 17β-estradiol (Aakvaag et al. 1990).
As a consequence of these culture conditions low BrdU incorporation levels were observed, most probably due to an increased cell doubling time (Roman et al. 1993; Aakvaag et al. 1990; Berthois et al. 1986). This resulted in weak BrdU immunosignals that made quantification of the BrdU labeling index very laborintensive and time-consuming and, more importantly, a considerable risk remained for underestimating the number of cells in S-phase.
Therefore, an immunofluorescence amplification procedure using fluorescein-labeled tyramide, based on the principle of CARD, was developed for cell cultures to enhance specifically the BrdU immunosignals. In contrast to the weak, heterogeneous BrdU staining using the conventional immunofluorescence protocol, the tyramide signal amplification method resulted in a much clearer and homogeneous BrdU immunosignal in the majority of the cells. The difference between BrdU-positive and BrdU-negative cells became clear-cut, which made quantification rapid, easy, and objective without the risk of underestimating the number of cells in S-phase. Some cells stained intensely but partially retained their punctate staining pattern, which is indicative of the specificity of the tyramide signal amplification in that it is restricted to the immediate surroundings of the HRP deposits. The BrdU labeling index determined after tyramide signal amplification was the same as that determined after conventional immunofluorescence staining. This suggests that the tyramide signal amplification method is specific and that no additional cells became positive.
The tyramide signal amplification for low BrdU levels might also lend itself to providing more accurate results for prognosis after in vivo BrdU administration. Another application of the tyramide signal amplification might be in cell kinetics studies. For that purpose, it would be necessary to adapt the current fluorescent amplification procedure so that cells could be stained in suspension for flow cytometric analysis. Furthermore, the tyramide signal amplification method might help in the development of computer-assisted image analysis. For this application, it is necessary to obtain consistently clear and stable immunostaining, resulting in high-contrast images that facilitate the identification of labeled and unlabeled nuclei.
In conclusion, we have described an amplification method using CARD to amplify specifically weak BrdU immunosignals due to low BrdU incorporation levels. Amplification was achieved by the use of fluorescein-labeled tyramide as a substrate for horseradish peroxidase. The tyramide signal amplification method holds the promise to be of great value for in vivo cancer prognosis, cell kinetics studies, flow cytometry, and computer-assisted image analysis.
Footnotes
Acknowledgements
We thank Lambert Leijssen and Hans Henderickx for the photographic layout.