
Abstract
We examined five nucleic acid binding fluorescent dyes, propidium iodide, SYBR Green I, YO-PRO-1, TOTO-3, and TO-PRO-3, for nuclear DNA staining, visualized by fluorescence and laser confocal microscopy. The optimal concentration, co-staining of RNA, and bleaching speeds were examined. SYBR Green I and TO-PRO-3 almost preferentially stained the nuclear DNA, and the other dyes co-stained the cytoplasmic RNA. RNAse treatment completely prevented the cytoplasmic RNA staining. In conventional fluorescence microscopy, these dyes can be used in combination with fluorescence-labeled antibodies. Among the dyes tested, TOTO-3 and TO-PRO-3 stained the DNAs with far-red fluorescence under red excitation. Under Kr/Ar-laser illumination, TOTO-3 and TO-PRO-3 were best suited as the nuclear staining dyes in the specimens immunolabeled with fluorescein and rhodamine (or Texas red).
I
Two types of fluorescent dyes have been commonly used for immunofluorescence microscopy, i.e., fluorescein and rhodamine and their derivatives. Fluorescein and rhodamine emit fluorescence of green and red under blue and green excitation, respectively. Combined use of different fluorochromes enables the simultaneous identification of different cell components. One of the disadvantages of fluorescence microscopy is its inability to delineate cellular structures other than those that are immunostained. Simultaneous staining of nuclei and/or actin filaments with appropriate fluorescent dyes greatly facilitates the visualization of the location and shape of the cells. DNA in cells is usually stained with DAPI (4'6-diamidino-2-phenylindol) for fluorescence microscopy (Takata and Hirano, 1990). When stained with DAPI, the DNA appears as blue-white fluorescence under ultraviolet (
Recently, a variety of nucleic acid binding dyes have been developed, mostly for gel staining. In this study we examined five nucleic acid-specific fluorescent dyes to determine whether they would be suitable for histochemical staining and observation by laser confocal and conventional fluorescence microscopy. In addition, we evaluated a triple fluorescence staining method employing fluorescence-labeled antibody, fluorescence-labeled phalloidin for the F-actin, and a DNA-specific fluorescent dye.
Materials and Methods
Specimens
Male 6-week-old Sprague-Dawley rats were anesthetized with pentobarbital sodium and specimens of jejunum and pancreas were sampled. The specimens were cut into small pieces and embedded in OCT compound (Tissue Tek; Miles, Elkhart, IN) and frozen with liquid nitrogen.
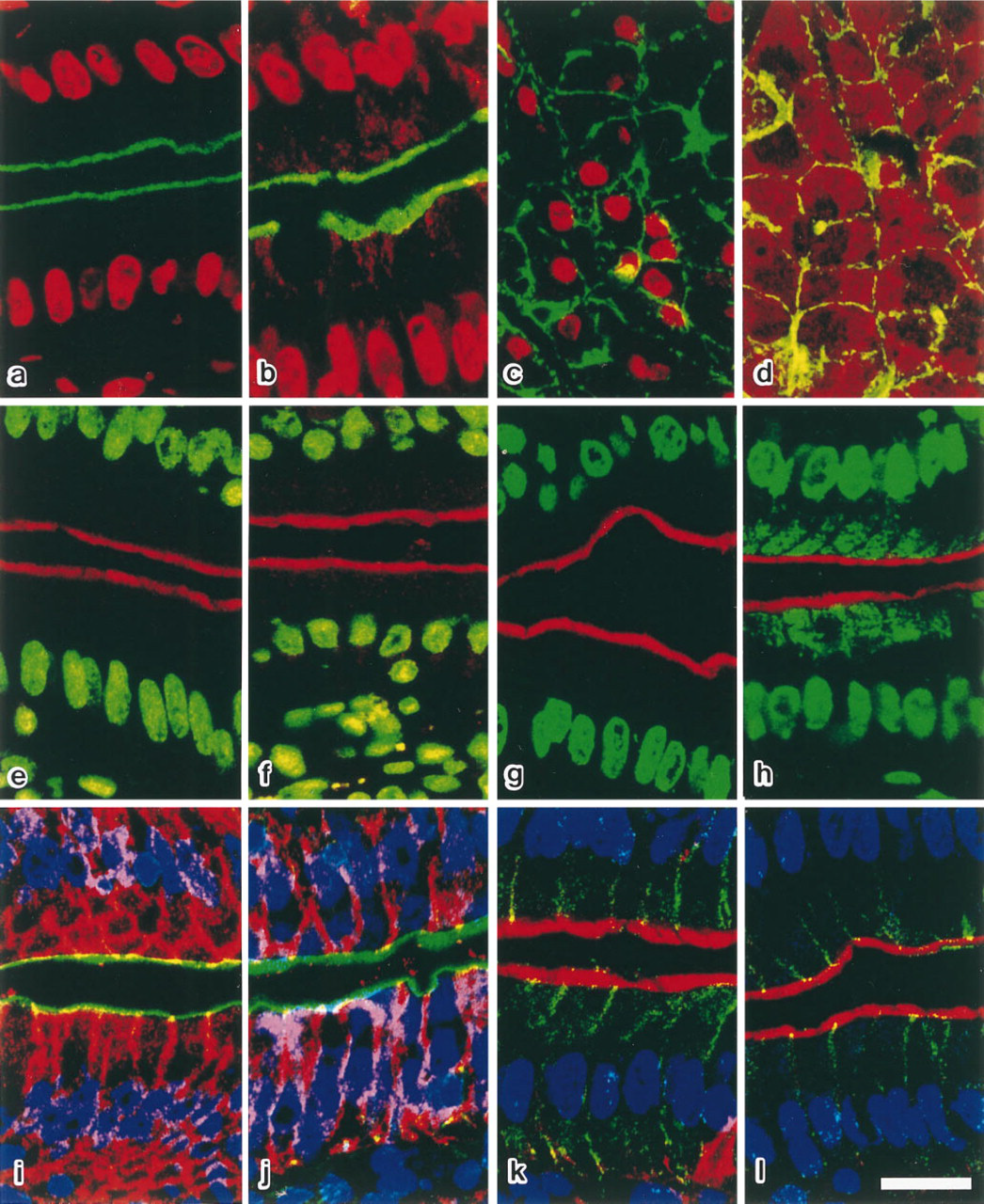
Laser confocal images of rat jejunum (
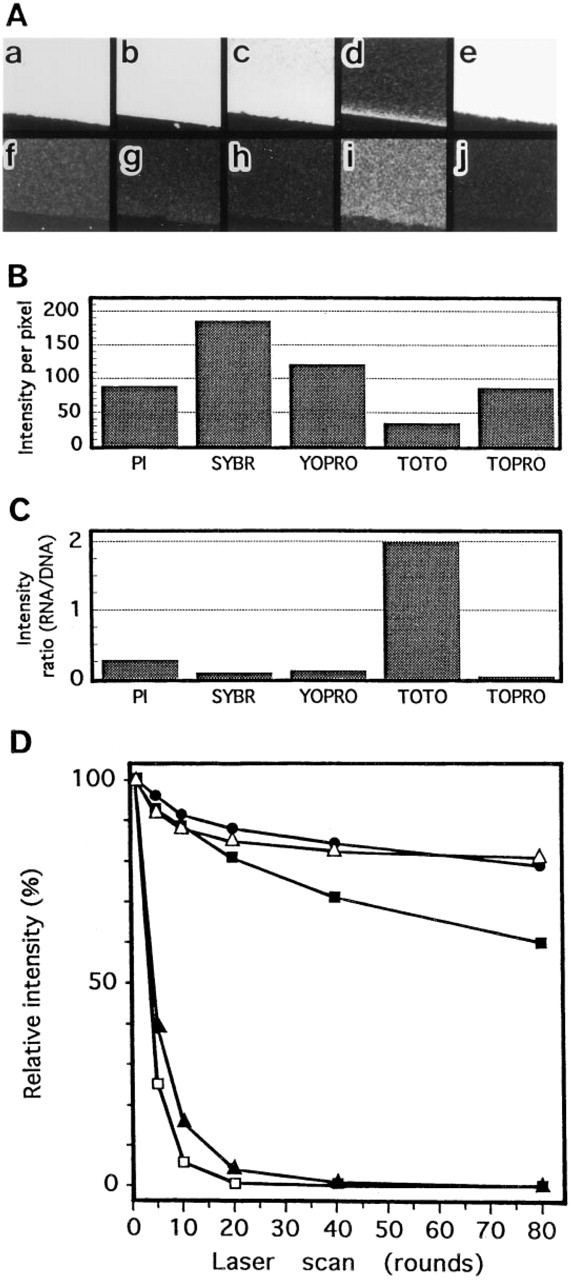
Specificity for DNA staining and bleaching characteristics of nucleic acid-binding fluorescent dyes. (
Immunofluorescence Staining and DNA Staining
Frozen sections (5 (μm thick) were cut with a cryostat and affixed to poly-
Laser Confocal Microscopy
Fluorescence-stained sections were examined under an epifluorescence microscope (BX-50; Olympus, Tokyo, Japan) equipped with a laser confocal system (MRC-1024; Bio-Rad Laboratories, Hercules, CA), comprising a 15-mW krypton/argon (Kr/Ar) laser (488-, 568-, and 647-nm excitations are possible) and three photomultiplier tubes with 522DF35, 605DF32, and 680DF32 emission filters. Image processing was carried out with LaserSharp computer software (Bio-Rad Laboratories).
In Gel Assay
Polyacrylamide gel sheets (8% acrylamide in TE buffer, 0.2 mm thick) including 300 μg/ml of salmon sperm DNA or yeast tRNA were cut into small pieces (about 5 mm square) and stained with either PI (2.5 μg/ml), SYBR Green I (1:500,000 dilution), YO-PRO-1 iodide (2 μM), TOTO-3 iodide (2 μM), or TO-PRO-3 iodide (1 μM) for 15 min. Gels were mounted and observed with a laser confocal microscope under illumination at 488 nm for PI, SYBR Green I, and YO-PRO-1 and at 647 nm for TOTO-3 and TO-PRO-3. The fluorescence intensity was measured on a 256-grade scale by the LaserSharp image processing software in each pixel (a pixel corresponds to 0.211 μm square) and average of 512 × 512 pixels was determined. A bleaching test was carried out by recording the fluorescence intensity on the designated round of laser scanning. Each round of scanning takes 1 sec, and each point in the specimen is irradiated for 3.8 μsec per scan.
Properties of the nucleic acid binding fluorescent dyes a
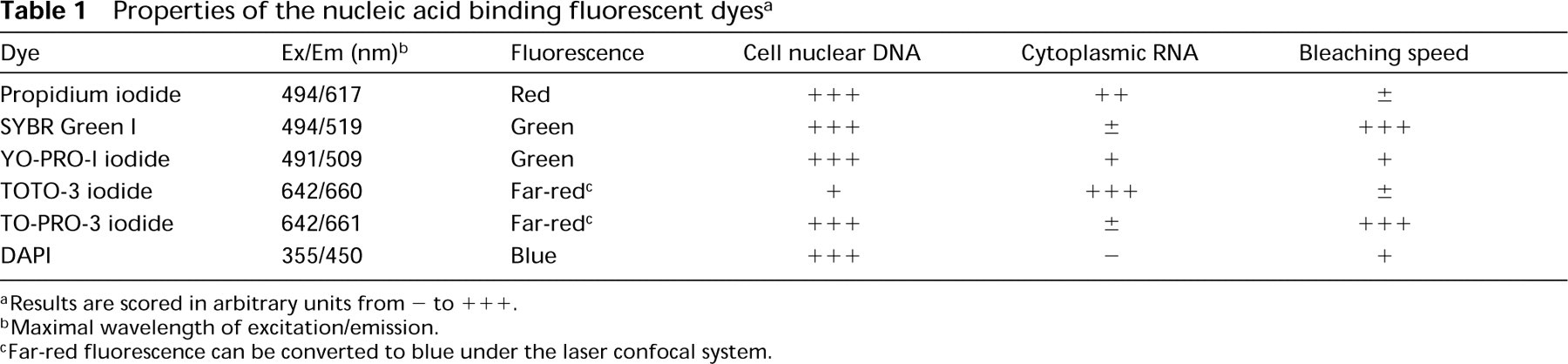
aResults are scored in arbitrary units from - to +++.
bMaximal wavelength of excitation/emission.
cFar-red fluorescence can be converted to blue under the laser confocal system.
Results and Discussion
When fluorescein or its derivatives was used as the fluorescent dye for immunostaining, PI was suited for DNA staining. In this case, DNA in the cell nucleus appears red under the green-yellow or blue excitation (568- or 488-nm laser excitation, respectively), and immunosignals are observed as green fluorescence under blue excitation (488-nm excitation) (Figure 1a; Table 1). Because PI binds to the nucleotide pair of guanine and cytosine, PI stains not only the DNAs but also the RNAs. When the RNAse treatment was omitted, the cytoplasm was stained as well (Figures 1b, 1d, 2A, and 2C). RNAse digestion at RT for 30 min usually eliminated this cytoplasmic staining, but some tissues that contain large amounts of cytoplasmic RNA, such as pancreas, needed to be treated with RNAse at 37°C for 30 min (Figures 1b-1d).
SYBR Green I intensely stained the DNA (Figures 1e and 2B). Cell nuclei stained with SYBR Green I appeared green under the blue (488 nm) excitation (Table 1). When rhodamine or its derivatives was used as the immunofluorescent dye in combination with SYBR Green I, immunosignals were detected in marked contrast against the nucleus as red fluorescence under the green-yellow (568 nm) excitation. SYBR Green I preferentially stained the nuclear DNA, as only a little cytoplasmic RNA staining was observed (Figures 1f, 2A, and 2C). RNAse treatment is therefore not needed for DNA staining with SYBR Green I. One of the disadvantages of SYBR Green I is that the fluorescence fades rapidly and the observation must be done as quickly as possible (Figure 2D). YO-PRO-1, an impermeable DNA binding dye, also stained the DNA with green fluorescence under the blue excitation (Figure 1g; Table 1) (Idziorek et al., 1995). Fluorescence fading of YO-PRO-1 was slower than that of SYBR Green I (Figure 2D), but the affinity of this dye for RNA was slightly higher than that of SYBR Green I. Therefore, RNAse treatment is recommended (Figures 1h, 2A, and 2C).
When dyes with red or green fluorescence, such as PI, SYBR Green I, or YO-PRO-1, were used for DNA staining, only one type of fluorescent dye could be used for immunostaining with visible fluorescence, since far-red fluorescence is not suitable for surveying the positive sites with a conventional fluorescence microscope before the confocal imaging. To allow triple staining, i.e., DNA, actin filaments, and an immunolabeled cell component, or DNA and two other immunolabeled cell components, we examined the two fluorescent dyes TOTO-3 and TO-PRO-3 for DNA staining. Both stained the DNA with far-red fluorescence under red (647 nm) excitation (Table 1) (Doornbos et al., 1994; Van Hooijdonk et al., 1994). This far-red fluorescence can be easily observed under a laser confocal microscope equipped with a Kr/Ar laser and a 680-nm emission filter or in the fluorescence microscope equipped with a Texas red filter. The signals emitted from TOTO-3 and TO-PRO-3 were detected as red fluorescence under the fluorescence microscope, but they could be converted to blue under the laser confocal system (Figures 1i-1l). Therefore, triple staining is feasible for laser confocal microscopy using these fluorescent dyes. TOTO-3 stained the DNA more weakly than the other fluorescent dyes, and intensely stained the cytoplasmic and nucleolar RNAs (Figures 1i, 1j, and 2A-2C). Thus, RNAse treatment is necessary for DNA staining with TOTO-3. However, the fluorescence stability of TOTO-3 was better than that of the other dyes (Figure 2D). TO-PRO-3 was highly specific for DNA, and more intensely stained the cell nuclei compared with TOTO-3 (Figures 1k, and 1l, and 2A-2C). The fluorescence, however, faded very rapidly (Figure 2D), and so the observation must be performed very quickly.
As summarized in Table 1, a variety of dyes with different characteristics can be used for nuclear counterstaining in immunofluorescence microscopy. Among them, the best-suited one can be chosen depending on the fluorochromes used for immunostaining, microscopes used (conventional or confocal), and the illumination light source.
Footnotes
Acknowledgements
Supported by the Dutch Science Organization, Area Medical Sciences (MW-NWO) project no. 900–543-109, and by a grant from Diabetes Fonds Nederland.
We wish to thank Dr H. Smeets of Nijmegen University Hospital for providing fibroblast cell cultures of Patient 2.