
Abstract
We performed ultrastructural immunogold localization of osteopontin in the mucosa of human stomach. This adhesive glycoprotein was present in mucous and chief cells of the epithelial layer and in macrophages in the lamina propria. Parietal and endocrine cells of the epithelial layer and mast cells and plasma cells in the lamina propria did not contain osteopontin, serving as internal negative controls. Subcellular localizations of osteopontin included secretory granules and synthetic organelles in mucous and chief cells and phagolysosomes in macrophages. Extracellular concentrations of osteopontin were present in the glycocalyx and in an electron-lucent band between epithelial surface cells and the gastric lumen. Paracellular edema between the epithelium of the same cells was devoid of osteopontin. Immunogold localization of pepsinogen II was done to identify cells with mixed granule populations and contents of multicompartmental secretory granules. These studies revealed mucous cell granules and chief cell granules, each containing compartmentalized storage products, which included osteopontin and mucigen in mucous cells and osteopontin and pepsinogen II in chief cells. Cytochemical controls for the immunogold localizations were negative. The subcellular distribution of osteopontin in human gastric mucosa suggests possible roles for this glycoprotein in barrier function, host defense, and/or secretion.
O
To gain a better understanding of the biology of OPN in epithelia, we have embarked on a systematic examination of subcellular localizations of OPN in the gastrointestinal tract, using postembedding immunogold preparations of human tissues (Qu-Hong et al. 1994). Initially, we examined gallbladder epithelium, a major site of OPN production in the gastrointestinal tract (Brown et al. 1992). These studies revealed that human gallbladder columnar epithelial cells contained OPN in Golgi structures, membrane-bound secretory granules, and in the glycocalyx (Qu-Hong et al. 1994). Secretion of OPN-rich granules by exocytosis from gallbladder epithelial cells was also evident (Qu-Hong et al. 1994). These findings were consistent with the expected distribution of a glycoprotein synthesized for export (i.e., glycosylated) in the Golgi apparatus (Neutra and Leblond 1966a,b; Caro and Palade 1964) and packaged into membrane-bound structures for exocytosis. For the study reported here, we used a similar approach to localize OPN in human gastric epithelium.
Materials and Methods
Case Material
Material for the ultrastructural immunogold analysis of gastric mucosal epithelium was obtained either from surgical biopsy specimens or from gastric specimens removed at the time of surgery from four patients. The electron microscopic samples were obtained from either chronically inflamed or histologically normal areas of gastric resection margins or biopsies, remote from areas of primary disease. The diagnoses for each patient follow: reflux esophagitis, adenocarcinoma arising in Barrett's esophagus, adenocarcinoma of the stomach, and malignant gastric stromal tumor. Small blocks of tissue were excised and fixed by immersion for 2 hr at room temperature (RT) in a mixture of 2.5% glutaraldehyde and 2% paraformaldehyde buffered at pH 7.4 with 0.1 M sodium cacodylate containing 0.025% calcium chloride, and were further processed for electron microscopy as described (Qu-Hong et al. 1994).
Tissue Processing
After primary fixation, tissues were washed in 0.1 M sodium cacodylate buffer (twice for 5 min) and stored in the same buffer at 4C for 1 or 2 days. Tissues were then postfixed in 1.33% OsO4 in 0.2 M Sym-collidine buffer, pH 7.4, for 2 hr at RT, washed with 0.05 M sodium maleate buffer, pH 5.2 (three times for 2 min), and stained with 2% uranyl acetate in 0.05 M sodium maleate buffer, pH 6, for 2 hr at RT. Finally, tissues were washed again with 0.05 M sodium maleate buffer, pH 5.2 (three times for 2 min), and dehydrated through graded ethanols. The samples were infiltrated in a propylene oxide-Epon 812 sequence and embedded and polymerized in Epon 812 at 60C for 16 hr. Gold to silver thin sections were cut on an LKB IV ultratome (LKB; Bromma, Sweden). The sections were picked up on uncoated gold grids (Pelco; Tustin, CA) and air-dried for 30 min.
Immunogold Labeling for Osteopontin
Grids were stained by submerging them (section side up) in 30-μl drops of reagents on Teflon-surfaced glass slides (Roboz Surgical Instruments; Washington, DC) in the following sequence at RT: (a) 0.1% Triton X-100 in 0.1 M Tris-buffered saline (TBS), pH 7.6, 10 min; (b) TBS containing 0.1% bovine serum albumin (TBS-BSA), three washes, 10 min each; (c) TBS-BSA containing 5% normal goat serum, 30 min; (d) primary antibody (affinity-purified rabbit anti-human OPN antibody, as described previously (Brown et al. 1992), 1:20 dilution in TBS-BSA containing 1% normal goat serum and 0.1% Tween-20, 60 min; (e) TBS-BSA, three washes, 10 min each; (f) secondary antibody (10-nm gold-labeled goat anti-rabbit IgG) (Energy Beam Sciences; Agawam, MA), 1:40 dilution in the same diluent as for primary antibody, 60 min; (g) TBS-BSA, two washes, 5 min each; and (h) two washes with double-distilled water, 5 min each. The grids were air-dried overnight.
Three specificity controls were employed, substituting for primary antibody as follows: (a) primary antibody to OPN that had been absorbed with OPN purified from human milk, as described (Senger et al. 1989); (b) irrelevant rabbit IgG at equivalent concentration; and (c) omission of the primary antibody.
Immunogold Labeling for Pepsinogen II
Grids were stained in a similar fashion in the following sequence at RT: (a) freshly made saturated sodium metaperiodate (Fisher Scientific; Medford, MA) in double-distilled water, 30 min; (b) double-distilled water, two washes, 5 min each; (c) TBS, two washes, 10 min each; (d) TBS-BSA containing 5% normal donkey serum, 30 min; (e) primary antibody (sheep anti-human pepsinogen II IgG) (Biodesign International; Kennebunk, ME), 1:20 dilution in TBS-BSA containing 1% normal donkey serum and 0.1% Tween 20, 60 min; (f) TBS-BSA, three washes, 10 min each; (g) secondary antibody (20-nm gold-labeled donkey anti-sheep IgG) (Goldmark Biologicals; Phillipsburg, NJ), 1:20 dilution in the same diluent as for primary antibody, 60 min; (h) TBS-BSA, two washes, 5 min each; and (i) two washes with double-distilled water, 5 min each. The grids were air-dried overnight.
Three specificity controls were employed, substituting for primary antibody as follows: (a) normal sheep serum or irrelevant sheep IgG at equivalent concentration; (b) omission of the primary antibody; and (c) absorption of primary antibody with solid-phase or soluble pepsinogen II (both from Sigma; St Louis, MO).
Before observation the grids were stained with lead citrate for 4 min and viewed in a CM-10 electron microscope (North American Philips; Mahwah, NJ).
Quantitation of gold particles that labeled mucigen granules and appropriate specificity controls were done on randomly obtained photographs of labeled cells. The number of gold particles/(μm2 of granule area was analyzed for significance with Kruskal-Wallis nonparametric ANOVA, Dunn's Multiple Comparison, and Mann-Whitney tests.
Results
The mucosa of chronically inflamed or microscopically normal human stomach biopsies, or surgical specimens, was examined to define cellular, extracellular, and subcellular sites of OPN using a postembedding immunogold procedure. Cell lineages that were examined in detail included mucous epithelial cells, chief cells, parietal cells, and endocrine cells in the epithelial layer, and macrophages, plasma cells, and mast cells in the subepithelial area of the mucosa. Of these cells, we found OPN in mucous epithelial cells, chief cells, and macrophages. Other cells examined did not contain OPN.
Mucous Cells
Subcellular and Extracellular Sites of Osteopontin. Mucigen granules, Golgi structures, small apical vesicles, the glycocalyx of microvilli, and an electronlucent extracellular layer between the apical cell surface and luminal contents of the stomach contained gold particles, indicating the presence of OPN (Figures 1-4). Background label of cytosol, nucleus, and mitochondria was minimal in OPN-positive mucous cells.
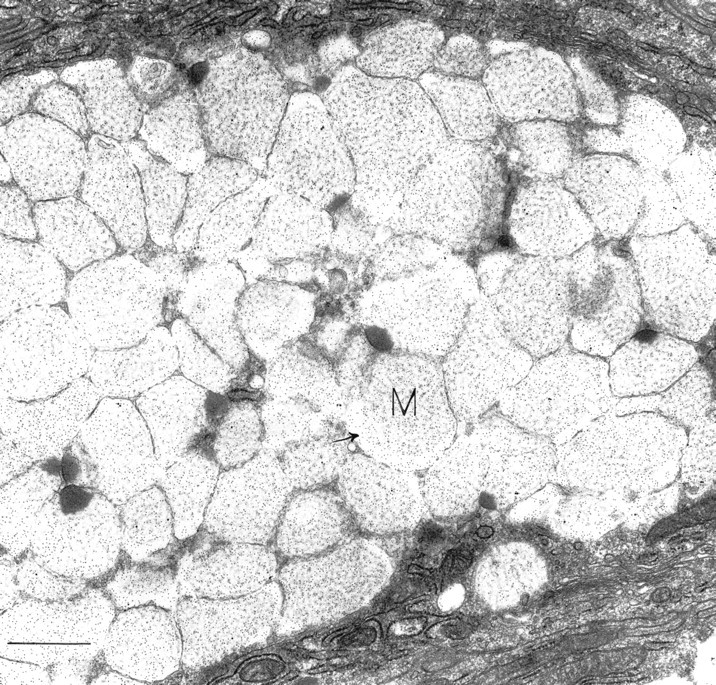
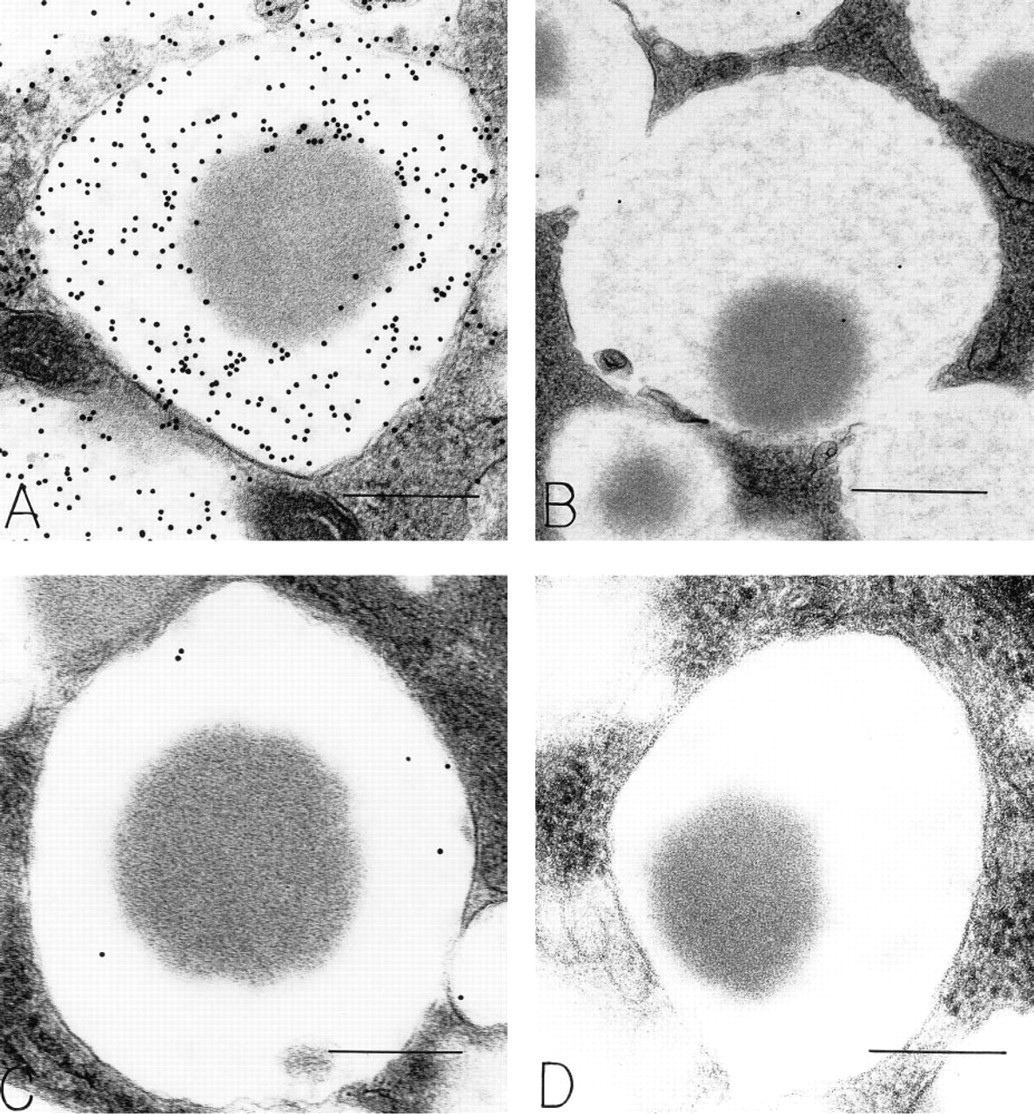
This and succeeding photographs are of gastric mucosa remote from areas of primary disease or microscopic areas of chronic inflammation. Osteopontin-positive surface mucous cell filled with mucigen granules (M). Note the OPN-positive reticular meshwork filling most granules. Several granules have peripheral electron-dense foci within them; these do not label for OPN. Many granules display peripheral electron-lucent areas (arrow) that are strongly labeled for OPN. Bar = 1.25 μm.
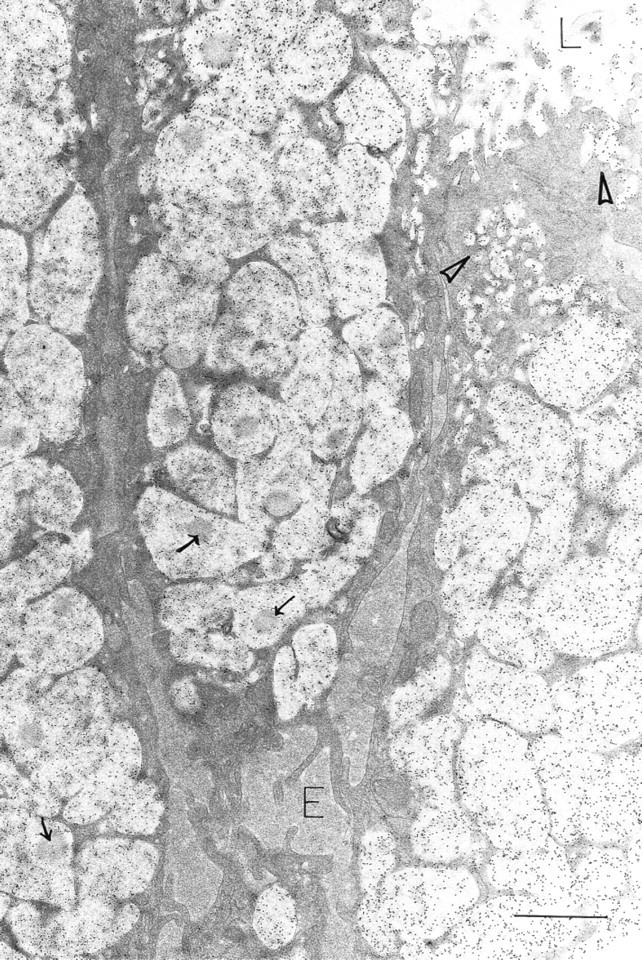
Surface mucous cells are filled with OPN-labeled mucigen granules. Rounded electron-dense areas within many mucigen granules do not label for OPN (arrows). Extensive protein-rich extracellular edema (E) outlines individual cells; OPN label is absent from this fluid. OPN is present in many small apical vesicles (arrowheads) and is attached to the apical cell surface and microvillous processes. L, gastric lumen. Bar =1.0 μm.
The Golgi structures that contained OPN were located on the mucigen granule-forming side oriented towards the apical membrane of mucous cells (Figure 4). Both immature small and mature large mucigen granules contained OPN. OPN was also present in granules undergoing secretion at the apical cell surface. Small apical cytoplasmic vesicles contained OPN (Figures 2 and 3A). Gold particles indicating OPN were often located near microvillous surfaces, in the glycocalyceal area (Figure 3A, inset). In some areas adjacent to the luminal surface of gastric epithelium, a broad electronlucent band with little to no detectable substructure bound large amounts of gold, indicating an OPN-rich layer overlying the gastric epithelium (Figure 3A).
Mucigen granules in mucous epithelial cells were readily identified by a reticular distribution of electron-dense strands in an electron-lucent background (Figure 1). In general, mucous cells were filled with reticular granules, and these granules contained OPN (Figure 1). In addition, mucous granules with central, rounded, electron-dense structures within the reticular meshwork were present. In these granules, OPN was confined to reticular portions of the granule substance, and the electron-dense areas were devoid of OPN (Figures 1, 2, 3A, 5A, 6A, and 7A). The electron-dense areas within mucigen granules of mucous cells did, however, label for pepsinogen II (Figure 7B), and most of these mucous cells were in the necks of gastric glands. Mucigen granules devoid of these electron-dense structures did not contain pepsinogen II (Figure 6B), and most of these mucous cells were surface-related. Some mucous cells contained mucigen granules displaying reticular and dense nucleoid domains, as well as peripheral elongated electron-lucent domains that were rich in OPN (Figures 1 and 3B). Rarely, large zymogen granules with entirely electron-dense contents were present within mucous neck cells. These granules were not labeled for OPN. The densities of gold label indicating the presence of OPN in mucigen granules (and appropriate specificity controls) are listed in Table 1. These counts show that mucigen granules have a significantly greater label of reticular areas (285.5/μm2) compared to electron-dense, rounded areas (32.6/μm2, p<0.001) or to specificity controls (0/μm2, p<0.001). There was no significant difference in OPN label of electron-dense granule areas (32.6/μm2) compared to controls (0/μm2, p = NS).
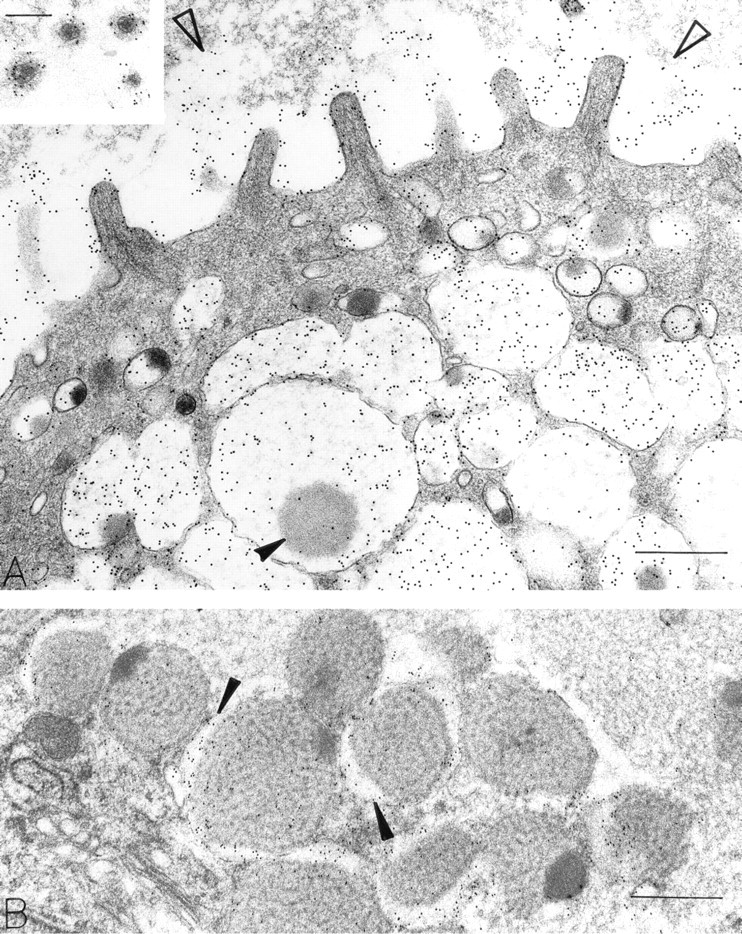
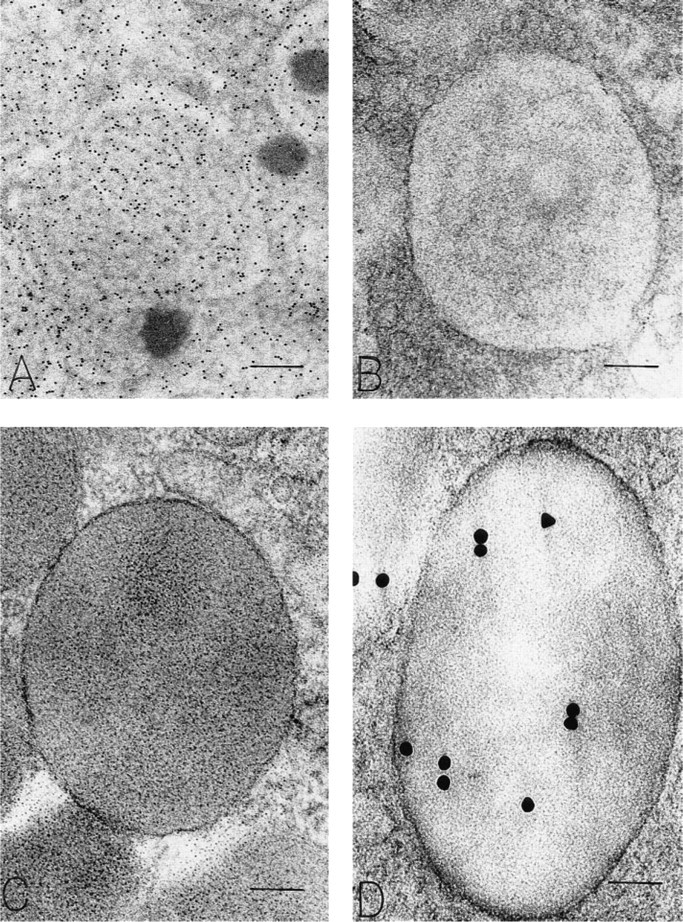
OPN-positive mucigen granules in surface mucous cells show variable density of the reticular mucigen. The lightly dense substructure in
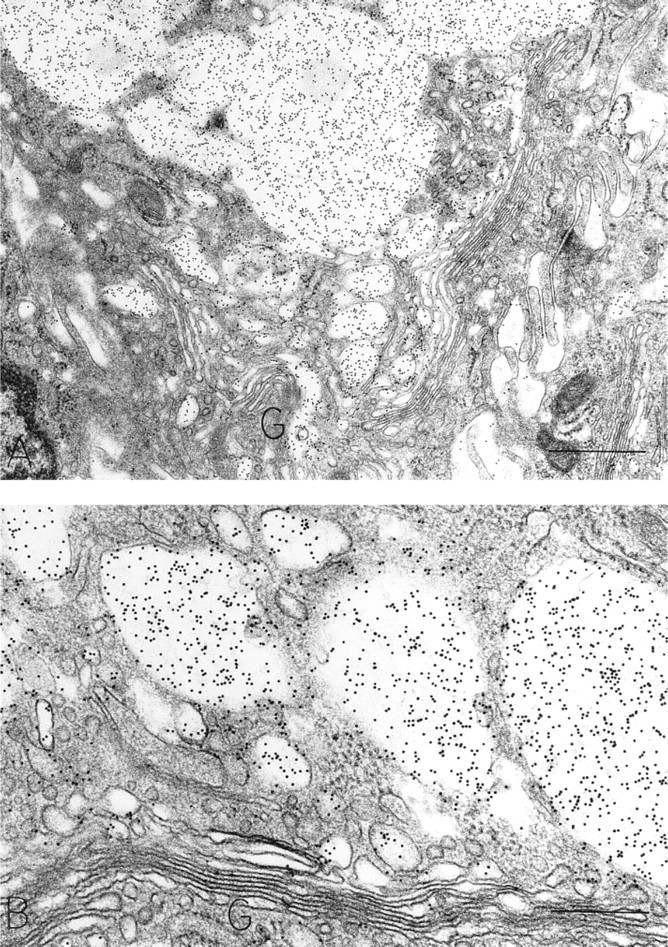
OPN-positive Golgi structures are present in these surface mucous cells. In each cell, a majority of the OPN-labeled membrane-bound structures is forming from the apically oriented side of the Golgi structure. Bars:
OPN staining (and specificity controls) in bicompartmental mucigen granules in surface mucous cells. (
Chief Cells
Subcellular Sites of Osteopontin. Chief cells, located primarily in the depths of gastric glands, contained distinctive large, membrane-bound, electron-dense secretory zymogen granules (Figures 6C and 6D). These homogeneously electron-dense granules contained pepsinogen II, as detected with postembedding immunogold stains (Figure 6D). Some zymogen granules in chief cells characteristically were bizonal, with approximately one half of the granule electron-dense and the remaining half electron-lucent. The proportions between these two compartments in zymogen granules were variable among chief cells, but granules that were completely electron-lucent or completely filled with a reticular mucigen meshwork were not present in chief cells. The electron-lucent compartment of zymogen granules in chief cells contained OPN, but the electron-dense, pepsinogen II-containing compartment was devoid of OPN (Figures 7C and 8). Therefore, chief cells contained pepsinogen II-positive granules (Figure 6D) and granules with mixtures of pepsinogen II and OPN in readily identified different subcompartments of their bizonal granules (Figure 8). Golgi structures in chief cells contained some label for OPN (Figure 7C). Background label in cytosol, nucleus, and mitochondria was minimal.
OPN-stained (
Macrophages
Macrophages, present beneath the epithelial layer of the gastric mucosa in chronically inflamed samples, often contained large numbers of phagolysosomes with heterogeneous contents. These structures contained OPN, as indicated by gold label over electron-lucent components of the heterogeneous contents (Figure 9). Electron-dense components did not label with gold. Other subcellular structures in these phagocytes also did not label with the immunogold stain to detect OPN. Background labeling of cytosol, nucleus, and mitochondria was minimal.
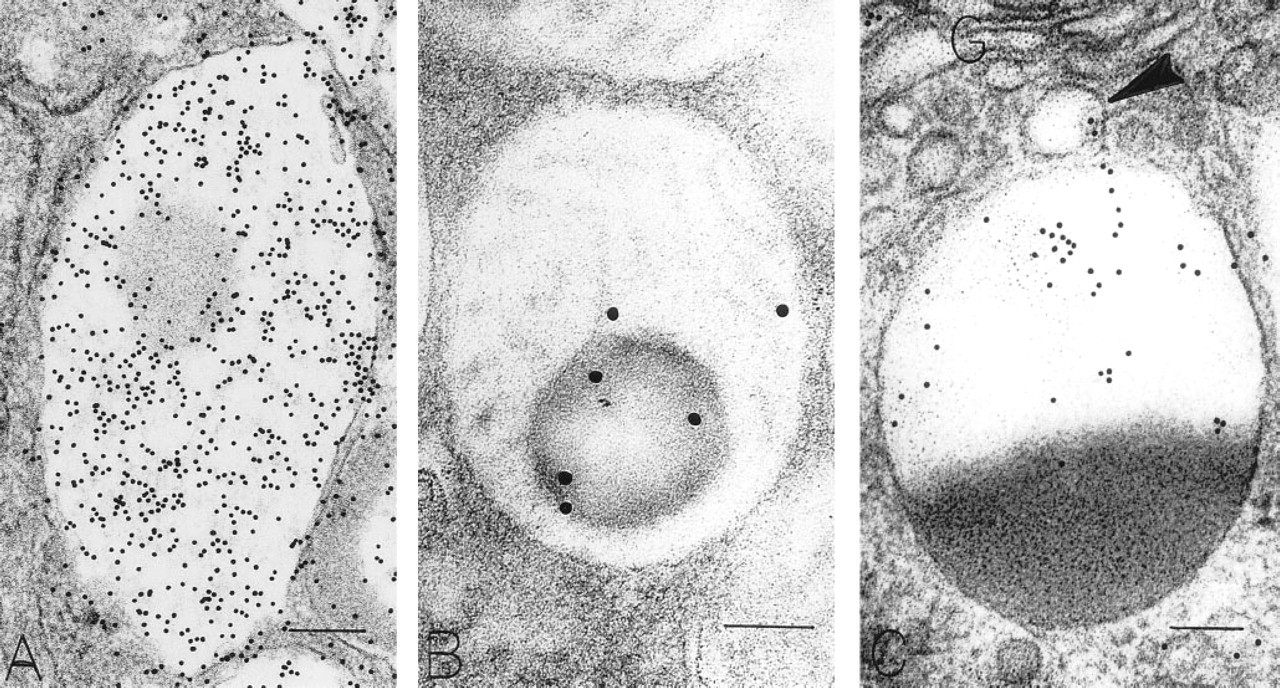
Mixed (bicompartmental) granules from a surface mucous cell (
Specificity Controls for Immunogold Stains
Substitution of irrelevant primary antibodies (Figure 5B) or omission of primary antibodies (Figure 5D) revealed no gold-labeled structures in gastric mucosal epithelial cells or macrophages. Absorption of the OPN-specific primary antibody with OPN (Figure 5C) or of the pepsinogen II-specific primary antibody with pepsinogen II (data not shown) resulted in marked diminution of specific gold-labeled structures.
Discussion
We report here the first subcellular localization of OPN, an adhesive glycoprotein (Weber et al. 1996; Senger et al. 1994; Chambers et al. 1993; Denhardt and Guo 1993; Brown et al. 1992; Kiefer et al. 1989; Wrana et al. 1989; Oldberg et al. 1986), in the mucosa of the human stomach. Subcellular sites of OPN were present in mucous cells and chief cells but were absent in parietal cells and endocrine cells of the epithelial layer of the mucosa. Macrophages, but not plasma cells or mast cells, in the lamina propria layer of the mucosa also contained OPN. In addition, a prominent OPN-positive extracellular layer extended over the luminal surface of many epithelial cells. OPN was absent, however, in paracellular, edema-filled extracellular spaces of samples that were chronically inflamed. Subcellular organelles that contained OPN included the glycocalyx of surface mucous cells, mucigen secretory granules of surface mucous cells, mixed OPN/mucigen/pepsinogen II secretory granules of glandular neck mucous cells, mixed OPN/pepsinogen II secretory granules of glandular chief cells, Golgi structures and forming granules of surface mucous cells, Golgi structures of chief cells, apical vesicles of surface mucous cells, and phagolysosomes of macrophages in the lamina propria.
Immunogold labeling for osteopontin in mucigen granules of human gastric mucous epithelial cells

∗ p<0.001 compared to electron-dense areas or controls.
∗∗ p=NS compared to controls.
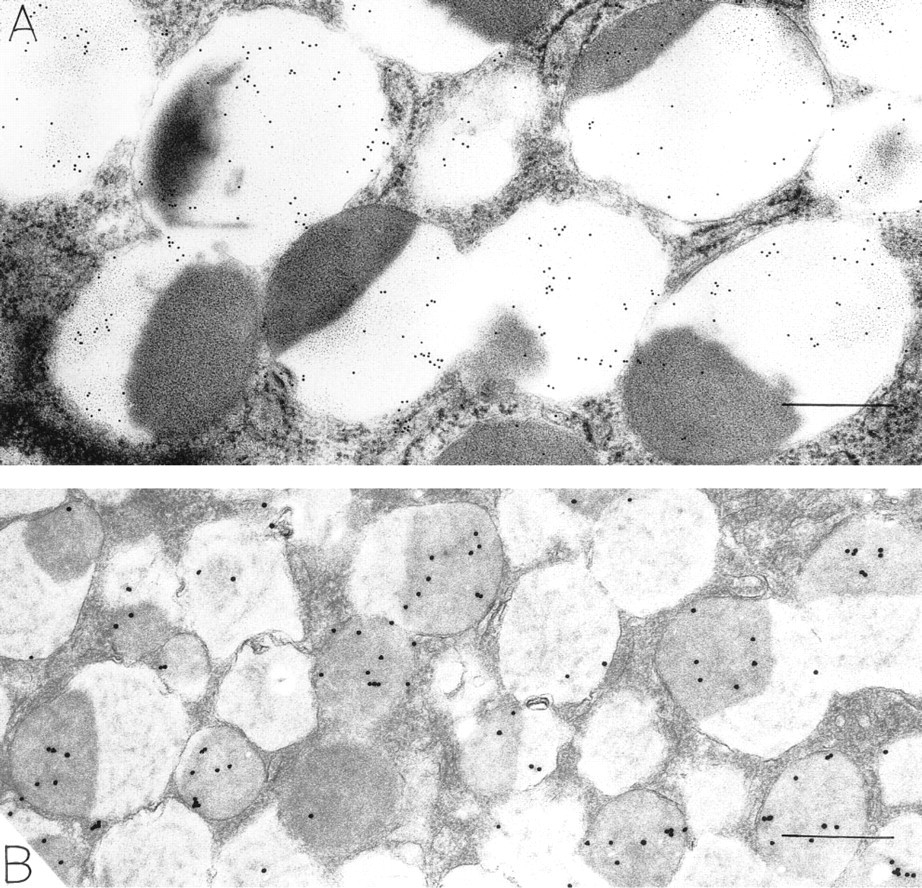
Chief cells in the glandular body are filled with mixed granules stained for OPN (
On the basis of the presence of the GRGDS sequence [the peptide sequence of OPN that promotes cell adhesion (Denhardt and Guo 1993; Kiefer et al 1989; Wrana et al. 1989; Oldberg et al. 1986)] and the prominent extracellular location of OPN in bone cell biology (McKee and Nanci 1993), some of the substructural localizations we define in the gastric epithelium are surprising. The cellular structures that are labeled in human stomach generally serve at least three major functions. These include barrier function, secretion, and phagocytosis, thereby implicating OPN in a variety of physiological and pathophysiological processes in gastric epithelium. Barrier function is a combination of the integrity of epithelial cells (which was not compromised in the samples examined here) and their overlying glycoprotein-rich attached glycocalyx and loosely associated mucous coat. Our immunogold staining for OPN clearly showed that the glycocalyx contained OPN, as we found to be true for gallbladder columnar epithelial cells (Qu-Hong et al. 1994). This layer, in the gastric epithelium, was vastly smaller than the extensive OPN-positive glycocalyx visualized in the gallbladder (Qu-Hong et al. 1994). Gastric luminal contents were usually devoid of OPN, but a thick electron-lucent band overlying the surface epithelium was richly endowed with OPN. Therefore, in addition to protective barrier functions ascribed to secreted mucus (Goldsworthy and Florey 1930; Florey 1962) in the stomach, the distinctive OPN-positive coat may also function as a protective barrier in this location. The source of the extracellular OPN layer is not known. It could arise from regulated secretion of OPN-containing mixed secretory granules or by constitutive secretion of OPN contained within prominent apical vesicles and subsequent attachment of these materials to the cell surface. Alternatively, OPN arising from oral gastrointestinal secretory organs (i.e., salivary glands) (Brown et al. 1992) could attach to the gastric epithelium and provide barrier protection in this location. In this case, OPN-containing apical vesicles could originate by endocytosis of OPN from this attached layer with subsequent traffic to Golgi structures. Alternatively, both constitutive secretion and endocytosis could be operative in the cellular cycling of OPN in gastric surface mucous cells. The static data we present here cannot distinguish among these possibilities. Similarly, phagocytosis by tissue macrophages is an important mechanism of host defense, and the presence of OPN in phagosomes of gastric mucosal macrophages suggests a role for phagocytosis of OPN in macrophage biology. A large number of reports describe the presence of the message for OPN as well as protein product in the macrophage lineage (Hirota et al. 1995; Brown et al. 1994; Murry et al. 1994; Pichler et al. 1994; Giachelli et al. 1993). Although we were unable to localize OPN to classical synthetic organelles in macrophages, it is possible that the necessary signal for immunolocalization is too low to detect with these tools. The OPN in macrophage phagolysosomes could originate internally by targeted traffic of synthetic transport vesicles to lysosomes or externally by phagocytosis, for example. Further experiments are necessary to choose between these alternatives.
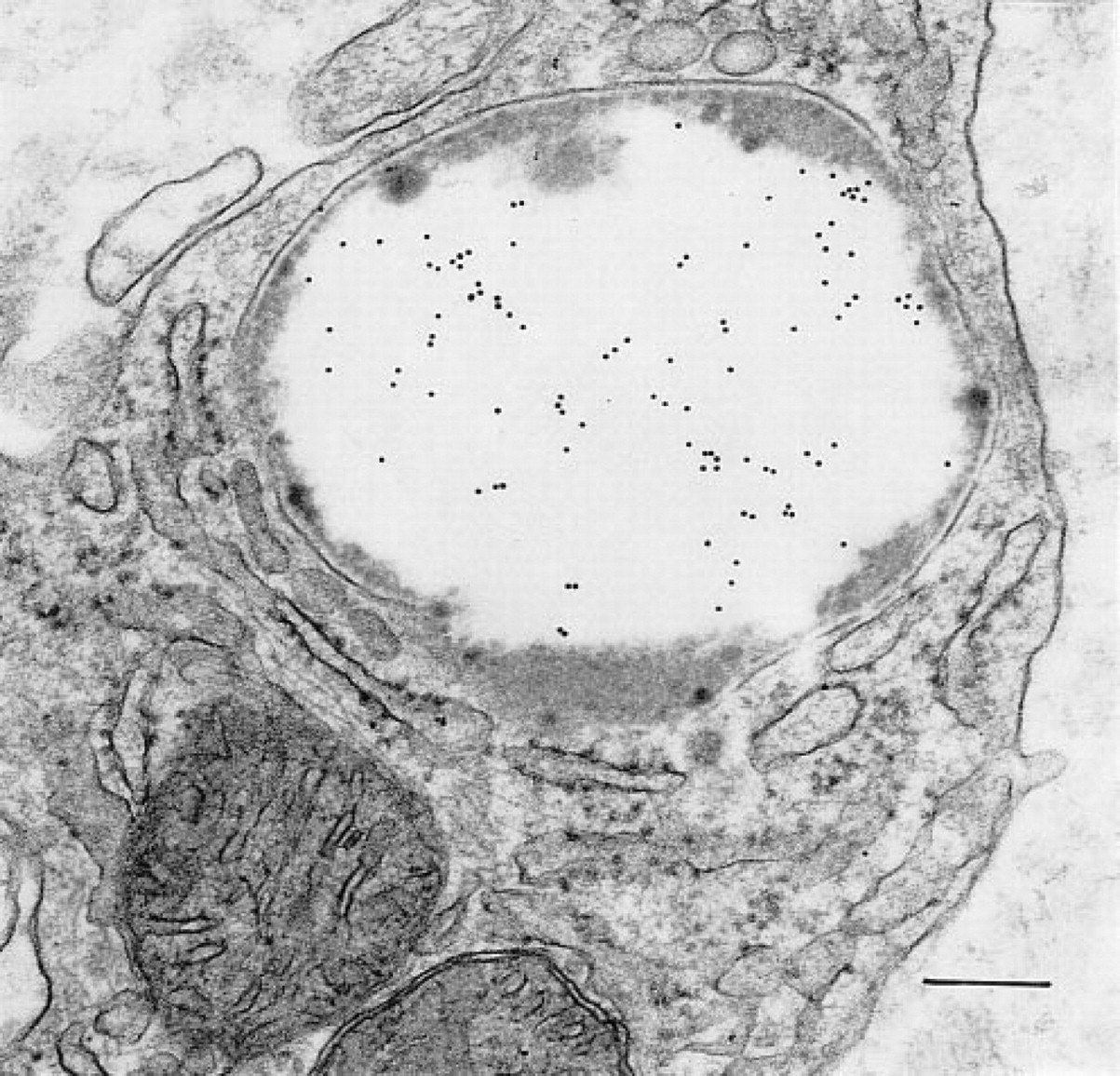
A portion of the cytoplasm of a subepithelial cell layer macrophage in the mucosal lamina propria contains mitochondria, rough endoplasmic reticulum, and a large phagolysosome. The central large area of electron-lucent material in the phagolysosome contains OPN. Gold label is not associated with the mitochondria or the rough endoplasmic reticulum. Bar = 0.2 μm.
Classical synthesis, granulogenesis, and regulated secretion (reviewed in Dvorak 1991) of OPN was, however, clearly evident in gastric epithelia in these studies. Thus, synthesis and packaging of new granules filled with OPN and secretion of the contents of the large mature granules by fusion with the apical membranes of epithelial cells were all evident. In total, the ultrastructural images providing documentation of the classical secretory pathway for OPN in gastric epithelia were similar to our findings in gallbladder epithelia (Qu-Hong et al. 1994).
The immunolocalization of OPN in human gastric mucosa also provides support for the existence of mixed-function epithelial cells, and compartmentalization of individual secretory granules, in this epithelial cell layer. The epithelium of gastric mucosa contains four epithelial cell categories: mucous cells, parietal cells, chief cells, and endocrine cells (Owen 1986). Mucous cells reside on the epithelial surface, interfaced with the gastric lumen, and in the neck region of gastric glands in the fundal mucosa. (In the pyloric mucosa, mucous cells are also glandular in location.) Parietal cells reside in the neck and body of gastric glands; their primary function is the production of hydrochloric acid. Chief cells, the zymogen granule-containing cells of the gastric mucosa, are principally located in the body of glands, as are granulated endocrine cells. Two of these cell categories (i.e., parietal and endocrine cells) were devoid of OPN in our samples, thus providing important negative controls for OPN staining in the same section. In addition, endocrine cells are secretory cells filled with secretory storage granules that did not contain OPN, thus providing documentation that OPN is not a matrix protein generally present in secretory granules. Moreover, and in support of light microscopic evaluations (Brown et al. 1994), the secretory granules of mast cells did not stain for OPN. Some mucous cells contained large numbers of mucigen (OPN-positive) granules and small numbers of zymogen (OPN-negative, pepsinogen II-positive) granules, indicating mixed functions for single cells in the gastric epithelium. Both mucous and chief cells had membrane-bound secretory granules with clearly delineated subcompartments. These were evident on inspection and contained different proteins by immunolocalization. Some granules were bicompartmental and contained either OPN plus pepsinogen II or OPN and mucigen. Others were trizonal, with OPN-, mucigen-, and pepsinogen II-rich domains. The necessity for and the function of subcompartment formation in secretory granules is unknown but may be important for differential secretion of granule matrix proteins. Other secretory cells also have subdivided granule compartments. For example, subcompartmen-talization of secretory granules is a prominent feature of the main granule populations in mature human eosinophils (Dvorak et al. 1991) and basophils (Dvorak 1996; Dvorak et al. 1995).
In summary, an ultrastructural immunogold study of the human gastric mucosa shows that OPN is a prominent component of this epithelium. We found it in the glycocalyx, in an overlying electron-lucent layer, and in apical cytoplasmic vesicles of epithelial cells as well as in Golgi structures, immature, mature, and secreting mucigen granules of mucous cells, in multi-compartmental granules in mucous cells and chief cells, and in phagolysosomes of macrophages. These locations suggest possible roles for this glycoprotein in barrier function, secretion, and host defense in the human gastric epithelium.
Footnotes
Acknowledgements
We thank Peter K. Gardner for assistance in the preparation of the manuscript and Jodi A. Wood for assistance with the photographic prints.
Supported by USPHS grants AI-33372 and DK-34854.