
Abstract
The endolymph in the endolymphatic sac (ES) is acidic (pH 6.6–7). Maintaining this acidic lumen is believed to be important for the normal function of the ES. The acid-base regulation mechanisms of the ES are unknown. Here we investigated the expression patterns of acid-base regulators, including vacuolar (v)H+-ATPase (proton pump), carbonic anhydrase (CA) II, and pendrin in the murine ES epithelium by immunohistochemistry (IHC) and compared their expression patterns by double immunostaining. We found that pendrin and vH+-ATPase were co-localized in the apical membrane of a specific type of ES epithelial cell. Pendrin- and vH+-ATPase-positive cells also expressed cytoplasmic CA II. Co-expression of pendrin, vH+-ATPase, and CA II in the same subgroup of ES cells suggests that this specific type of ES cell is responsible for the acid-base balance processes in the ES and pendrin, vH+-ATPase, and CA II are involved in these processes.
T
The functional significance of an acidic ES lumen has been postulated to be related to endolymph homeostasis, which is essential for the normal function of the inner ear (Couloigner et al. 2000). Mechanisms for generation and maintenance of this acidic lumen are poorly understood. Carbonic anhydrase (CA), an enzyme that catalyzes the hydration of CO2 to ultimately yield bicarbonate and protons, was initially proposed as being involved in the acidification, based on the observations that the ES epithelium contained high levels of cytosolic CA and that the CA inhibitor acetazolamide caused a decrease in luminal pH and bicarbonate concentration (Yamashita et al. 1992; Tsujikawa et al. 1993). More recently, mRNA for the vacuolar (v) H+-ATPase β1-subunit was detected in specific cells lining the mouse ES lumen (Karet et al. 1999; Dou et al. 2003). Immunocytochemical experiments also showed vH+-ATPase subunit E in the apical membrane of ES epithelial cells in guinea pig (Stankovic et al. 1997). The vH+-ATPases are a family of multisubunit ATP-dependent proton pumps responsible for intracellular as well as luminal or interstitial space acidification (Nelson and Harvey 1999; Alper et al. 2002). Identification of proton pumps in ES cells suggests their role in ES pH regulation. Further supporting evidence is provided by pharmacological studies in guinea pig demonstrating that luminal administration of a specific inhibitor of the proton pump, bafilomycin, resulted in significant increase of the ES luminal pH (Couloigner et al. 2000).
In addition to CA and vH+-ATPase, other acid–base regulatory proteins, including anion Cl–/HCO3 – exchangers (AEs) and pendrin, also may be involved in the regulation of acid-base balance in the ES. AEs are a family of variably expressed membrane proteins that exchange Cl– for HCO3 – across the plasma membrane. They are the products of at least three homologous genes of the SLC4 family encoding Na+-independent Cl–/HCO3 – exchangers that are designated as AE1, AE2, and AE3. These polypeptides contribute to regulation of intracellular pH or cellular electrochemical equilibrium potentials for Cl–, HCO3 –, and H+ (Alper et al. 1997,2002). Conversely, pendrin is a membrane protein encoded by the Slc26A4 gene (Pds), which belongs to a superfamily of anion exchangers (SLC26) (Everett et al. 1997). Mutations in Pds cause Pendred syndrome, a genetic disorder characterized by sensorineural deafness and goiter (Everett et al. 1997,1999). Recent data have shown that pendrin mediates Cl–/HCO3 – exchange (Royaux et al. 2001; Soleimani et al. 2001) and is downregulated in response to metabolic acidosis (Wagner et al. 2002; Frische et al. 2003; Petrovic et al. 2003), suggesting that pendrin may function as an acid–base regulator. Immunoreactive signals to antibodies that can recognize both AE1 and AE2 have been identified at the basolateral membrane of ES epithelial cells in guinea pig (Stankovic et al. 1997). Pendrin mRNA and protein have also been detected in a subpopulation of murine ES cells (Everett et al. 1999; Royaux et al. 2003). These data suggest that AE and pendrin may participate in acidification processes in the ES.
Although different acid–base regulatory proteins have been localized in mammalian ES cells (Yamashita et al. 1992; Stankovic et al. 1997; Royaux et al. 2003), there is lack of information on the spatial relationship of their expression patterns, which is important for the understanding of the function of these acid-base regulators. The purpose of this study was to determine the cellular expression patterns of pendrin, vH+-ATP-ase, and CA II in the murine ES epithelium by IHC. An additional objective was to compare their expression patterns by double immunostaining. Proteins that participate in pH regulation of the ES endolymph have not been extensively studied. The information from this study would provide direct in situ morphological information on the distribution patterns of these molecules in the ES, which would improve our understanding of ES acid–base regulatory mechanisms.
Materials and Methods
All animals were handled and maintained according to an IACUC- and Cincinnati Children's Hospital Research Foundation-approved protocol.
Antibodies
An affinity-purified rabbit polyclonal antiserum raised against a synthetic pendrin peptide corresponding to amino acids 734–752 (CKSREGQD-SLLETVARIRDC) was used to detect pendrin. The sequence of the synthetic peptide used for antibody generation was identical for rat, mouse, and human pendrin and was previously reported (Petrovic et al. 2003). The generation and characterization of rabbit polyclonal antibody against the α4-subunit of vH+-ATPase (α4) also has been previously described (Herak-Kramberger et al. 2000). An affinity-purified goat antibody against a peptide mapping near the N-terminus of CA II of human origin was purchased from Santa Cruz Biotechnology (Santa Cruz, CA). Biotinylated goat anti-rabbit IgG, biotinylated rabbit anti-goat IgG (Vector Laboratories; Burlingame, CA), fluorescent isothiocyanate (FITC)-conjugated donkey anti-goat IgG, fluorescent FITC-conjugated Fab fragment of goat anti-rabbit IgG (H + L), and lissamine-rhodamine-conjugated donkey anti-rabbit IgG (Jackson ImmunoResearch; West Grove, PA) were used as secondary antibodies.
Tissue Preparation
Five postnatal-day (P) 5 and five 5-week-old (adult) mice were sacrificed by CO2 inhalation and the inner ears were removed and fixed in 4% paraformaldehyde dissolved in 0.1 M PBS overnight. Ears from P5 mice were decalcified in 0.12 M EDTA at room temperature (RT) for 3 days, while ears from adult mice were decalcified in 0.12 M EDTA at RT for 12 day. Then the ears were cryoprotected in 30% sucrose. After embedding in OCT (Tissue Tek; Miles, Elkhart, IN), 10-μm-thick sections were cryostat-cut, placed on Superfrost Plus slides (Fisher Scientific; Pittsburgh, PA) and stored at −20C until use.
Immunohistochemistry
Localization of acid-base regulators in the ES was performed using an immunoperoxidase procedure. Sections were washed in PBS, then incubated with 0.5% H2O2 in methanol (Fisher Scientific) for 15 min, followed by 5% normal rabbit serum (Sigma Chemical; St Louis, MO) for anti-CA II or 5% normal goat serum (Sigma) for anti-α4 and anti-pendrin primary antibodies for 30 min. After removal of the excess serum, the sections were incubated with the primary antibodies for 3 hr at 4C. Antibody dilutions for CA II, α4, and pendrin were 1:1000, 1:3000, and 1:300, respectively. Negative controls were performed by incubating slides with normal serum in place of primary antibodies. After three washes in PBS, sections were incubated with secondary antibodies (biotinylated goat anti-rabbit IgG or rabbit anti-goat IgG; Vector) diluted 1:200 in PBS for 30 min at RT. Avidinbiotin complex was then applied (Vector). Color was developed with 3,3-diaminobenzidine tetrahydrochloride (DAB; Sigma).
Immunofluorescence Double Labeling
Double staining of pendrin and α4 with CA II (antisera raised in different species) was performed sequentially. For the first immunoreaction, sections were washed in PBS, then incubated with 0.5% H2O2 in methanol (Fisher Scientific) for 15 min followed by 5% normal goat serum for 30 min. After removal of the excess serum, the sections were incubated with the primary antibodies of either rabbit anti-α4 (1:2000) or rabbit anti-pendrin (1:100) overnight at 4C. Then the sections were washed three times in PBS and incubated with a secondary rhodamine-conjugated donkey anti-rabbit IgG (red color) (1:200) for 1 hr at RT. Sections were washed in PBS five times (10 min each time), then processed for CA II immunolabeling.
The slides were similarly incubated in 5% normal rabbit serum for 30 min, then incubated with anti-CA II antibody (1:500) overnight at 4C. After washing in PBS three times, sections were incubated with an FITC-conjugated donkey anti-goat IgG (green color) (1:200) for 1 hr. Thereafter, sections were rinsed in PBS three times (10 min each time) and mounted using Vectashield mounting medium (Vector).
Double staining of pendrin and α4 (antisera raised in the same species) was performed according to the method described by Negoescu et al. (1994). The cryostat sections were incubated first with rabbit polyclonal anti-α4 antibody (diluted 1:2000) overnight at 4C, followed by an FITC-conjugated Fab fragment of goat anti-rabbit IgG (H+L) (diluted 1:200) at RT for 1 hr After rinsing, sections were incubated with unlabeled Fab fragments of goat anti-rabbit IgG (H+L) (diluted 1:100) for 3 hr (Jackson) to block all possible remaining binding sites of the first primary antibody. Afterwards, sections were incubated with a second rabbit polyclonal antiserum against pendrin (diluted 1:200). This primary antibody was detected with rhodamine-conjugated donkey anti-rabbit IgG (1:200). After a final wash, sections were mounted in Vectashield. Controls for crossreaction of the first and second immunolabeling were performed by omitting the primary antibody in the first immunolabeling step.
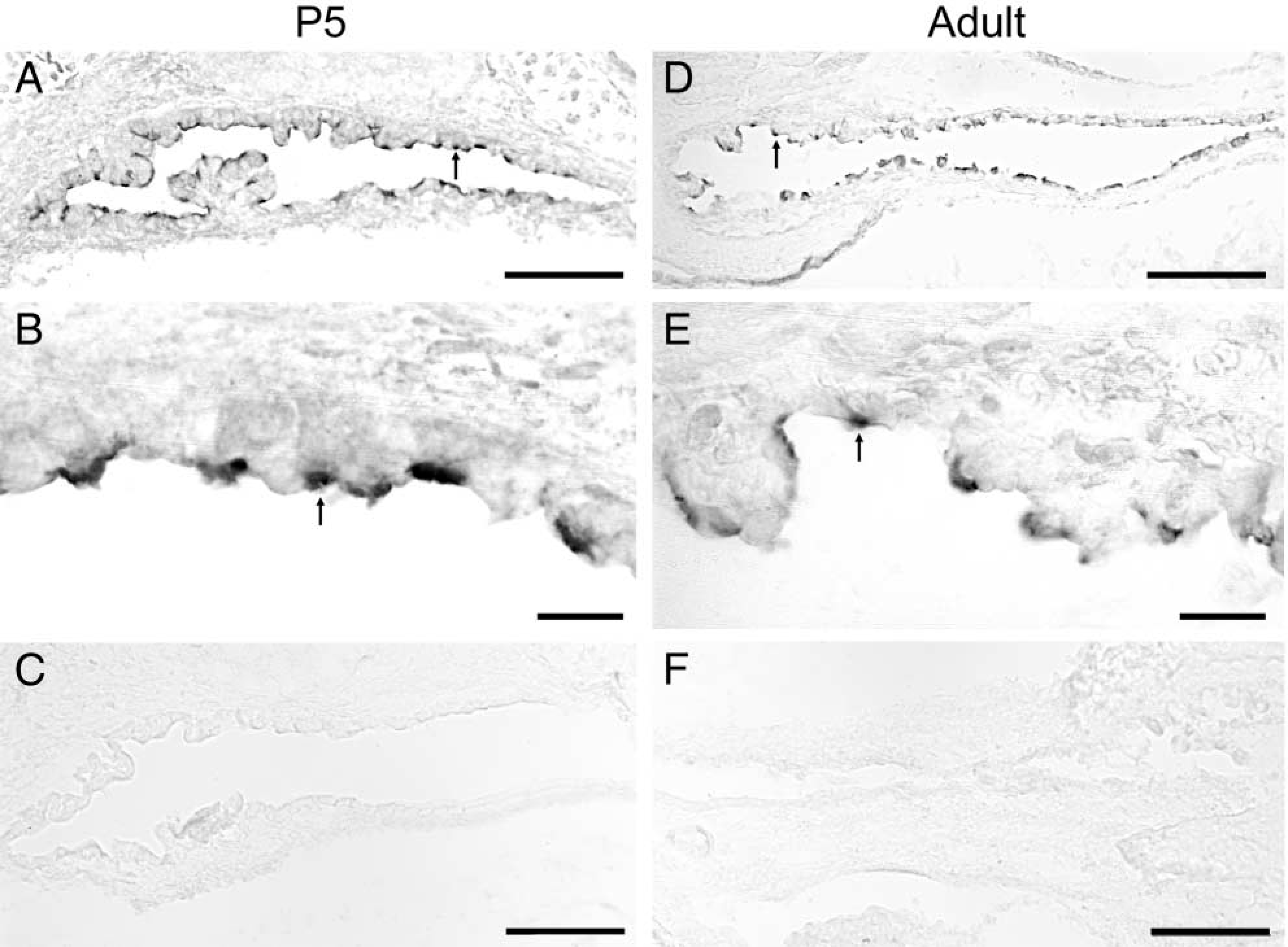
Expression of pendrin protein in the murine endolymphatic sac. Strong immunoreactive signals were observed in a subpopulation of the ES cells of both developmental stage (P5) (arrow in
All immunofluorescence labeling slides were examined with a Nikon epifluorescence microscope with fluorescein filters.
Results
Expression of Pendrin in the Murine ES
Figure 1 illustrates the IHC labeling pattern observed with the antibody against a synthetic pendrin peptide on cryosections of the mouse inner ear. Strong immunoreactive signals were observed at the apical regions of a subpopulation of ES cells of both P5 (arrows in Figures 1A and 1B) and adult mice (arrows in Figures 1D and 1E). The pattern of immunostaining, with pendrin-positive cells interspersed among unlabeled cells, suggests the heterogeneous nature of the ES epithelium. No labeling was seen in sections without the primary antibody (Figures 1C and 1F).
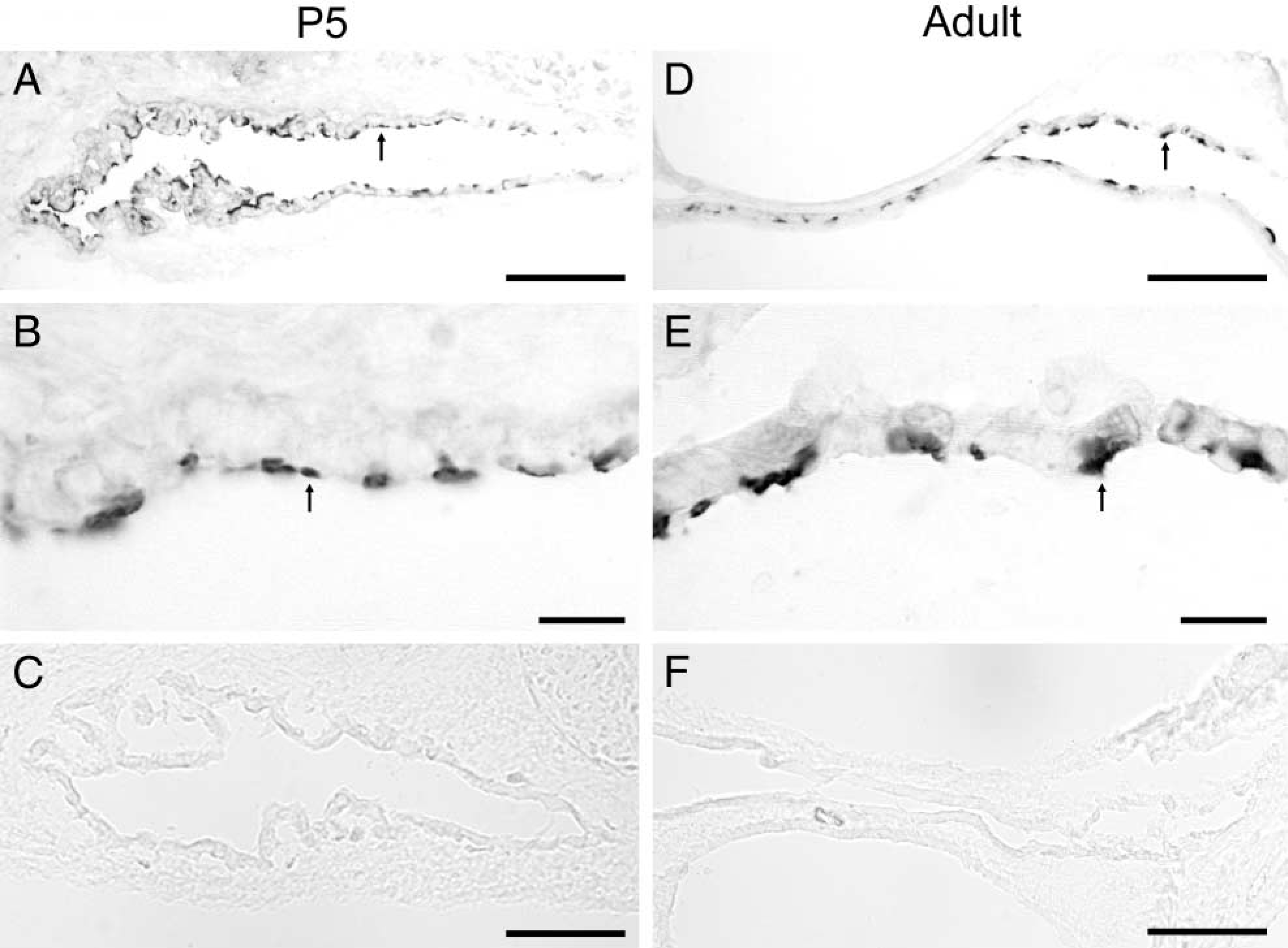
Expression of vH+-ATPase in the murine endolymphatic sac. Intense immunoreactive signals were also observed in a subgroup of the ES cells of both P5 (arrow in
Expression of vH+-ATPase in the Murine ES
Figure 2 shows the IHC labeling pattern of vH+-ATP-ase in murine ES cells using an antibody raised against the α4-subunit of vH+-ATPase (α4). Intense vH+-ATP-ase immunoreactivity was also observed at the apical cell sides of both P5 (arrows in Figures 2A and 2) and adult (arrows in Figures 2D and 2E) mouse ES. The expression pattern is also specific and heterogeneous throughout the ES epithelium. Sections incubated with normal serum in place of the primary antibody showed no immunoreactive signals (Figures 2C and 2F).
Expression of CA II in the Murine ES
Figure 3 shows the staining pattern of CA II. Strong cytosolic immunoreactive signals of CA II were observed in discrete cells of the ES of both P5 (arrows in Figures 3A and 3B) and adult mice (arrows in Figures 3D and 3E). Slides with omission of the primary antibody did not show any immunostaining (Figures 3C and 3F).
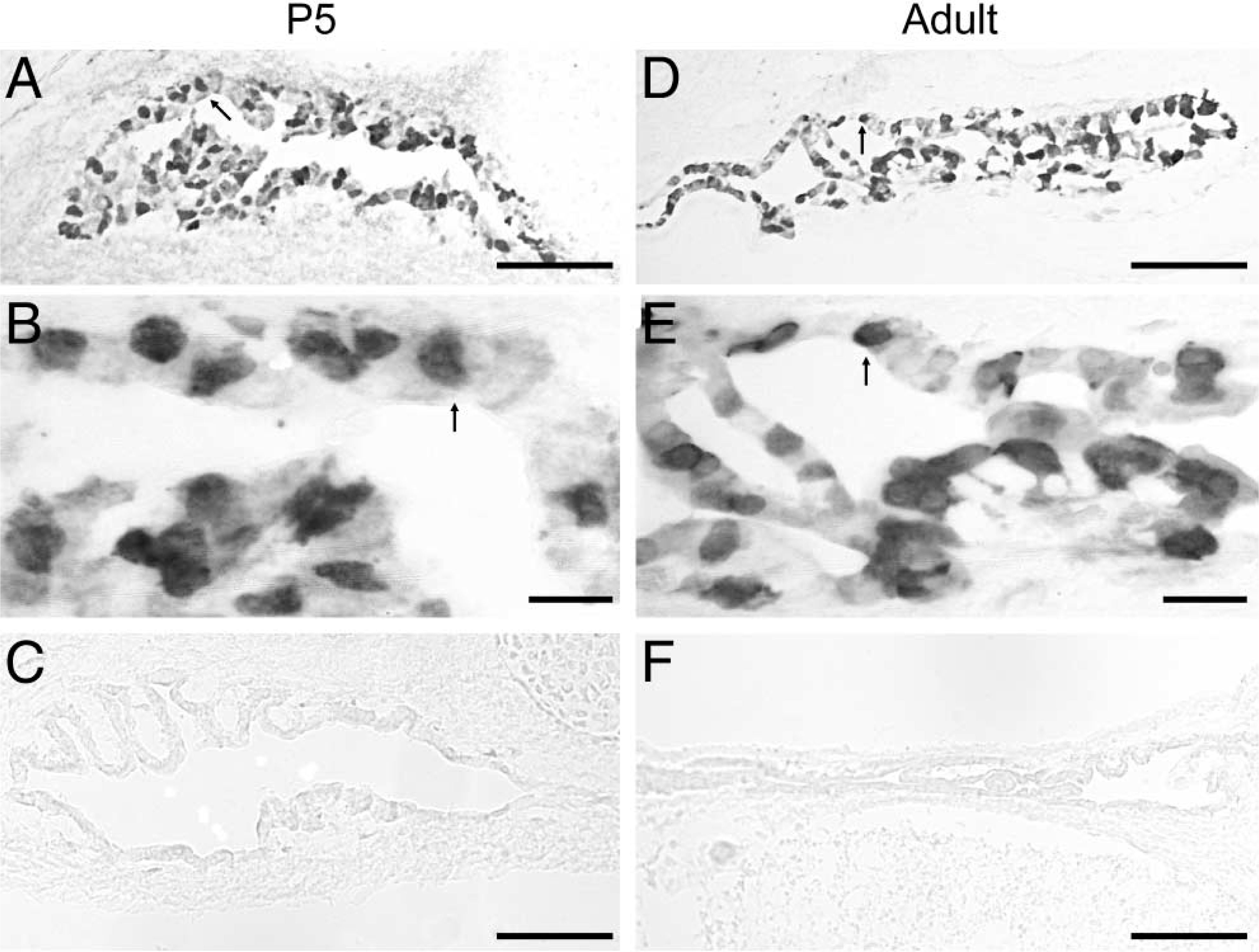
Expression of carbonic anhydrase II (CA II) in the murine endolymphatic sac. Strong cytosolic immunoreactive signals of CA II were observed in discrete cells of the ES of both P5 (arrow in
Co-localization of Pendrin and α4 with CA II
Similar expression patterns of pendrin, vH+-ATPase, and CA II in ES epithelial cells prompted us to investigate if they localize in the same population of ES cells by immunofluorescence double-labeling techniques. Immunoflurescence staining of pendrin (red) (Figure 4A), α4 (red) (Figure 4D), and CA II (green) (Figures 4B and 4E) gave the same general patterns of staining as immunoperoxidase-based labeling (Figures 1A, 2A, and 3A, respectively). Merged images revealed that cells with strong apical staining for pendrin (red) or α4 (red) were also positive for CA II (green) (Figures 4C and 4F, respectively). These data suggest that pendrin and vH+-ATPase α4-subunit co-expressed with CA II in the same group of ES cells.
Co-localization of Pendrin with vH+-ATPase α4-Subunit
Both pendrin- and α4-immunoreactive cells also positive for CA II suggest that pendrin co-localizes with α4 in the apical membrane of the ES cells. Further direct evidence was provided by double immunostaining of pendrin with α4. The distribution of pendrin (red) (Figure 5A) and α4 (green) (Figure 5B) is completely identical when a dual image was acquired (Figure 5C). Omission of primary antibody in the second immunolabeling step failed to show double labeling (data not shown), suggesting that the immunoreaction from the second immunolabeling was not the result of crossreaction.
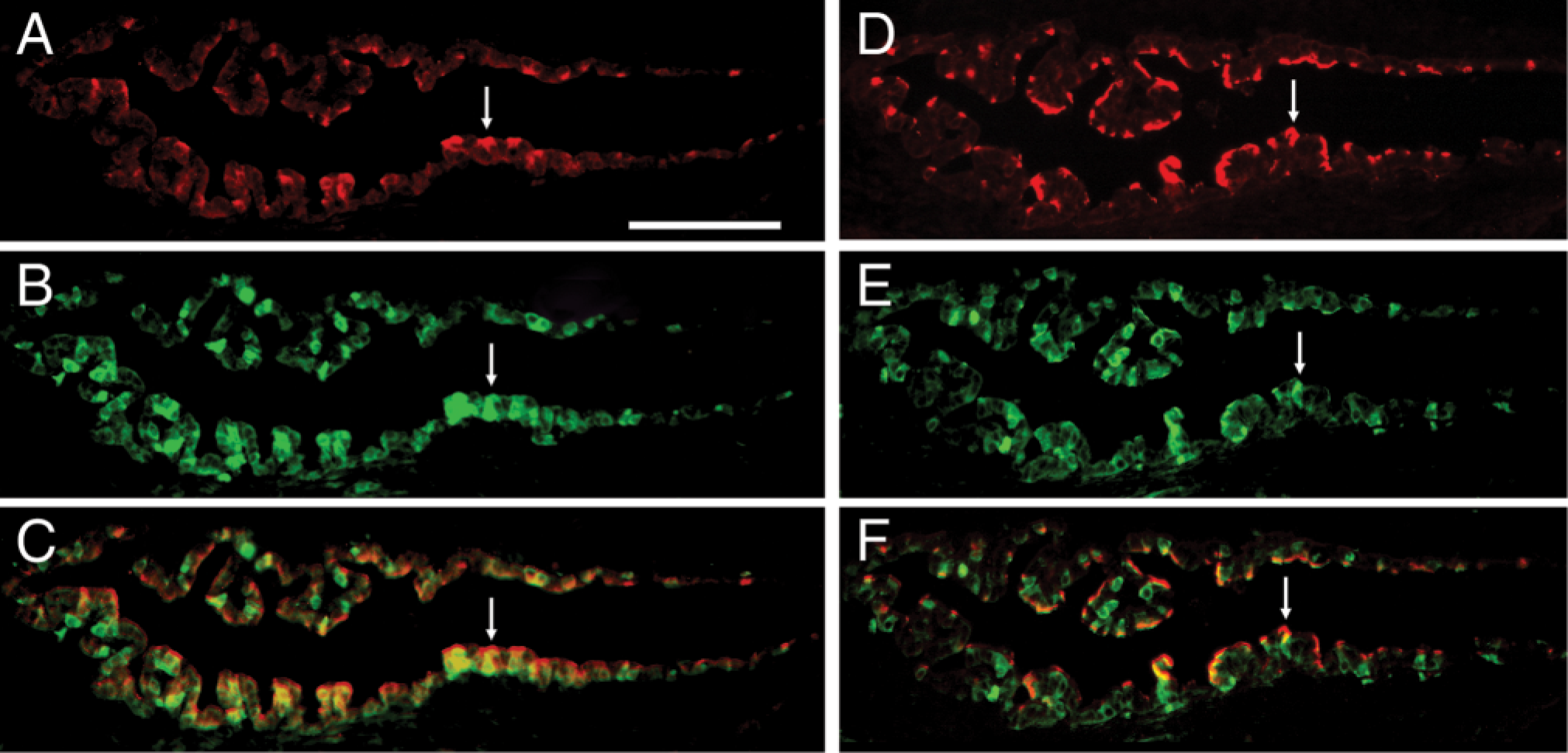
Immunofluorescence double labeling of pendrin and vH+-ATPase with CA II. Fluorescence images of the ES stained with anti-pendrin (
Discussion
The ES epithelium is characterized by plasma membrane infoldings, wide lateral intercellular spaces, vesicles, and microvilli, which are similar to those of other ion- and fluid-transporting cells (e.g., intercalated cells of renal nephrons) (Lundquist et al. 1984; Peters et al. 2002,2003). These intercalated cells are the main cell type involved in the acid–base transport in the kidney and are classified into subtypes, type A and type B cells. Type A intercalated cells express apical vH+-ATPase and basolateral AE1, resulting in the secretion of H+ into the lumen coupled with a trans-epithelial reabsorption of HCO3 –. Conversely, type B intercalated cells secrete HCO3 – by a coordinated action of apically located pendrin and basolaterally expressed proton pumps (Royaux et al. 2001; Petrovic et al. 2003). Whereas the mechanisms of H+ secretion and HCO3 – absorption to maintain acid-base balance in the kidney are relatively well established (Rodriguez-Soriano 2000; Wagner and Geibel 2002; Petrovic et al. 2003), studies on the acid-base regulation mechanisms in the ES are very limited.
In this study we used IHC to identify the cellular expression patterns of pendrin, vH+-ATPase, and CA II in ES epithelial cells. We show that pendrin is co-expressed with the α4-subunit of vH+-ATPase and CA II in the same subgroup of ES cells. The role of pendrin in the ES is unclear. However, on the basis of our data and the observation that pendrin functions as an acid–base regulator in the kidney (Wagner et al. 2002; Frische et al. 2003; Petrovic et al. 2003), we propose that pendrin may function as an acid-base transporter in the ES by mediating Cl–/HCO3 – exchange across the apical membrane of the ES cells. Loss of function of pendrin may cause pH changes in the ES endolymph, which would affect normal absorptive function of the ES (Lundquist et al. 1984) and result in enlargement of endolymphatic duct and sac, the pathology often seen in patients with Pendred syndrome as well as in knockout mice (Everett et al. 1997,1999,2001).
Incorporating our results and previous findings of AE on the basolateral site of ES cells (Stankovic et al. 1997), we have employed a working model of acid-base regulation in the ES epithelium (Figure 6). We hypothesize that polarized distribution of acid-base transporters as well as cellular distribution of an acid-base buffering system in the specific type of the ES cells are involved in the ES endolymph acid-base regulation. Protons are secreted into the ES lumen through apically located proton pumps coupled with transepithelial reabsorption of HCO3 – and secretion of Cl– through apically located pendrin. Parallel, basolaterally located anion Cl–/HCO3 – exchangers (AE) facilitate H+ and Cl– secretion by transporting HCO3 – out of the cells across the basolateral membrane. Cytoplasmic CA II participates in H+ secretion process by catalyzing the formation of HCO3 – and H+ in the presence of H2O and CO2.
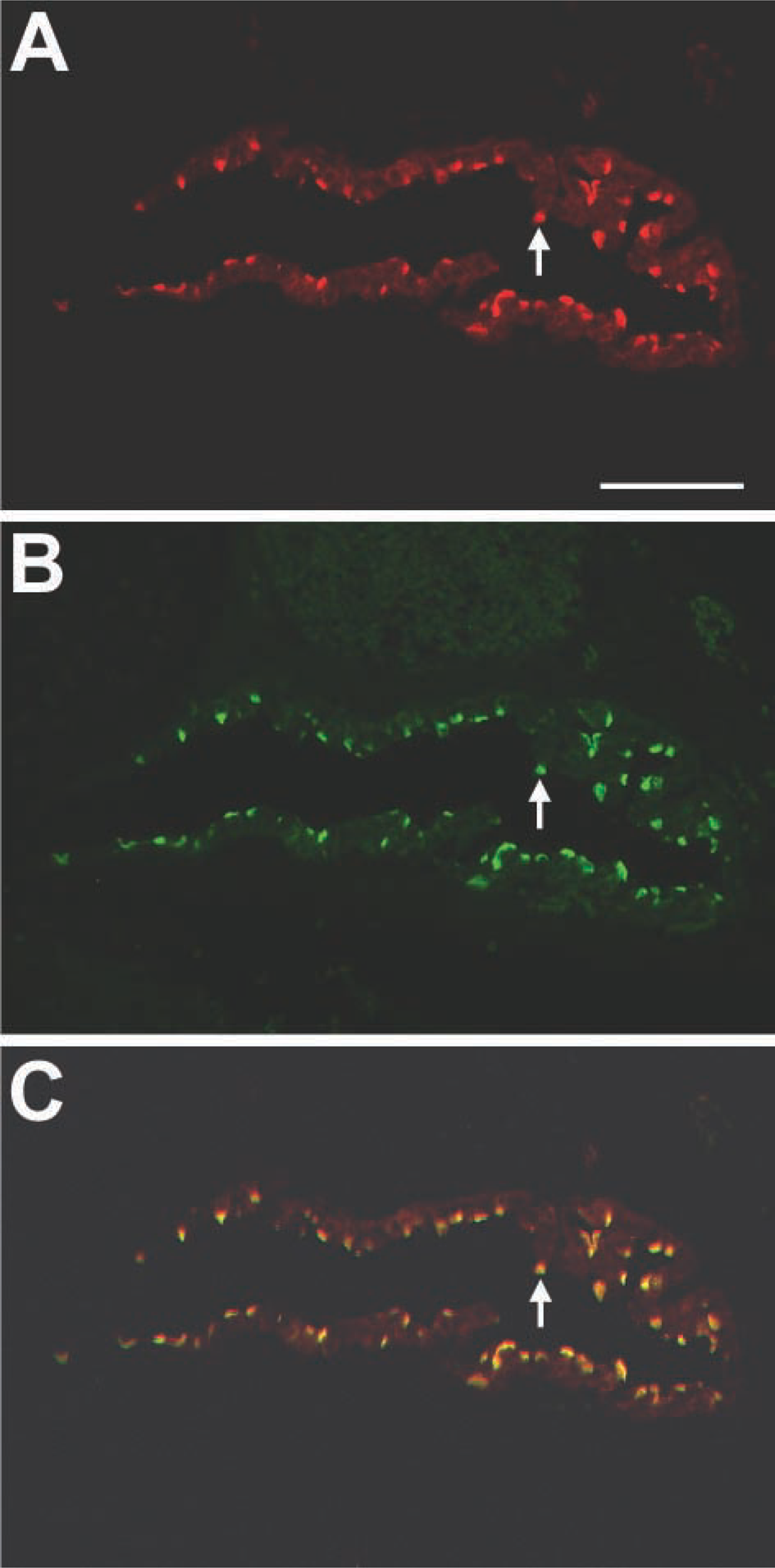
Co-localization of pendrin with vH+-ATPase α4-subunit. The distribution of pendrin (red) (
Co-expression of pendrin, vH+-ATPase, and CA II in the same subpopulation of ES cells also suggests that the specific cell type of the ES epithelium is responsible for the regulation of the ES acid-base balance. Earlier reports based on electron microscopic studies have suggested two different cell types encompassing the ES epithelium, i.e., light and dark cells, or mitochondrion-rich and ribosome-rich cells, respectively (Lundquist et al. 1984; Peters et al. 2002). On the basis of our light microscopic studies, we cannot establish precisely which of the two cell types is responsible for the acid-base regulation. Further ultra-structural studies in which pendrin, vH+-ATPase, and CA II expression can be ascribed to a specific subtype of the ES cells may help to answer the question.
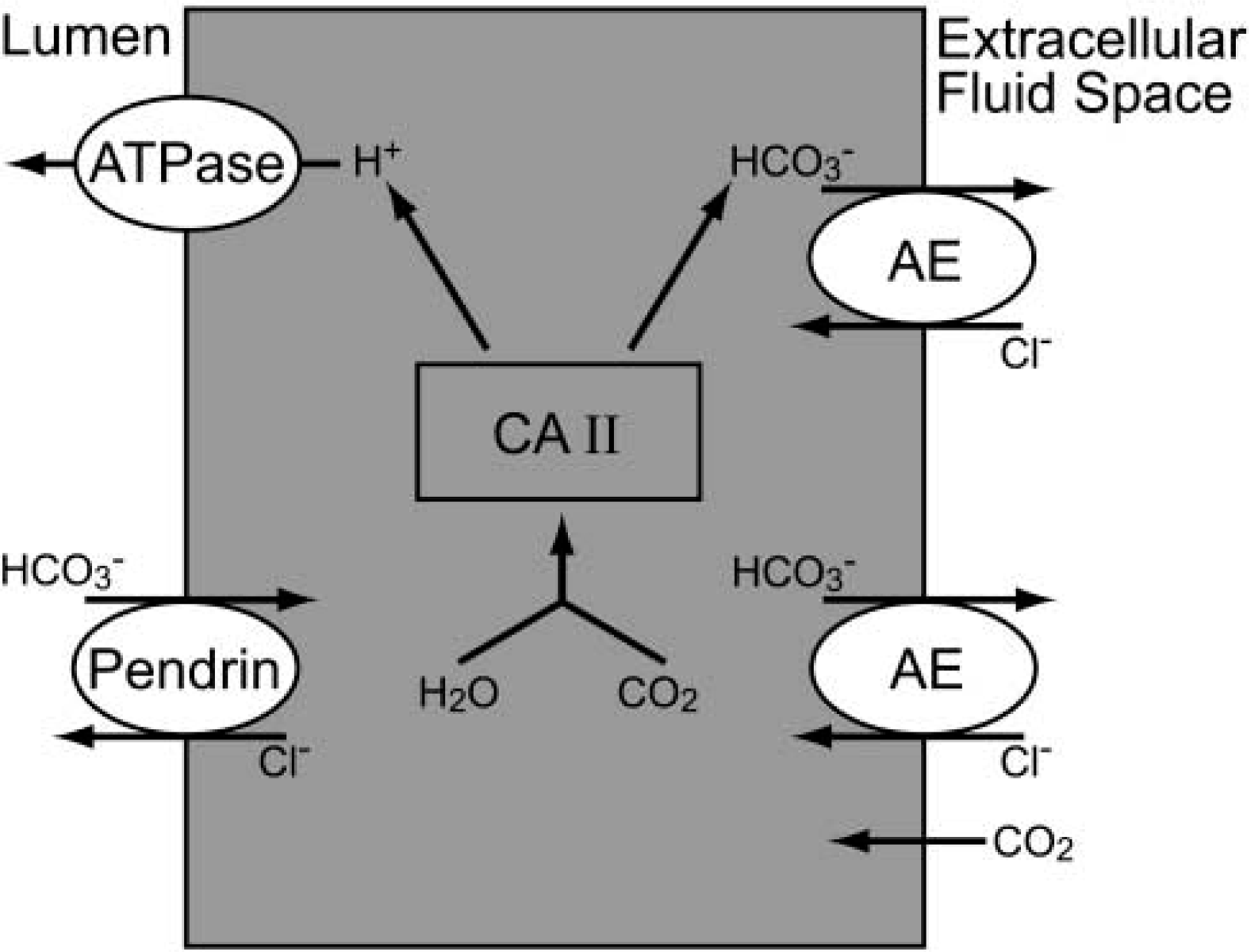
Hypothetical cellular model for acid-base regulation in the murine endolymphatic sac. Protons are secreted into the ES lumen through apical located proton pumps coupled with transepithelial reabsorption of HCO3 – and secretion of Cl– through apically located pendrin. Basolateral located anion Cl–/HCO3 – exchangers (AE) facilitate H+ secretion by transporting HCO3 – out of the cells across the basolateral membrane. Cytoplasmic CA II participates in the H+ secretion process by catalyzing the formation of HCO3 – and H+ from H2O and CO2.
In summary, the ES endolymph is acidic and the mechanisms involved in the acid-base balance processes have not been extensively studied. The cellular expression patterns of pendrin, vH+-ATPase, and CA II demonstrated in this study suggest that a specific subset of cells in the ES epithelium are responsible for ES acid-base homeostasis and that coordinated activities of pendrin, vH+-ATPase, and CA II are involved in the processes.
Footnotes
Acknowledgements
Supported by a grant from the National Institute on Deafness and Other Communication Disorders (DC00193) (D.C.) and by the Wellcome Trust (F.E.K. and A.N.S.), DK 54430, and DK 62829 (MS),
We thank Mr Joseph Alward for assistance in the preparation of this manuscript.