
Abstract
We describe the application of the biomolecular interaction (BIA) technique to detection of the interaction between protein (e.g., c-Jun) and DNA (e.g., two AP-1 motifs from bcl-2 promoter), compared with immunohistochemistry (IHC) of c-Jun. The specific binding assay for the interaction of c-Jun and activating protein-1 (AP-1) motifs was performed using a Biacore 2000 system. Intense immunoreactivity of c-Jun in glandular cells of the human uterine endometrium was observed in the proliferative phase, while c-Jun in stromal cells was expressed throughout the menstrual cycle. In contrast to the IHC of c-Jun, the specific binding of c-Jun to two separate AP-1 motifs in the bcl-2 promoter region was detected only in nuclear extracts of glandular cells, but not in stromal cells, during the proliferative phase. These results indicate that, while transmitting various signals, c-Jun enhances the transcription level of bcl-2, which in turn keeps glandular cells alive and proliferating in normal human endometrium during the proliferative phase. Moreover, the method involving real-time biomolecular interactions such as DNA–protein binding is novel for the study of transcription factors when combined with IHC.
T
Among many candidate binding sites in the bcl-2 5′-regulatory region (–1600 bp to −1 bp), two separate specific DNA sites should be recognized by the AP-1 family. The mammalian AP-1 family comprises several bZIP (basic region leucine zipper) proteins that can form either homo- or heterodimers among themselves (Angel and Karin 1991). The AP-1 proteins had been described as a family of transcription factors that is 12–0-tetradecanoyl-phorbol-13-acetate (TPA)-responsive. The dimerization of AP-1 factors is necessary for their binding to a palindromic DNA sequence, TGAC/GTCA, known as the TPA-responsive element (TRE) or AP-1 consensus site. Different AP-1 dimers exhibit similar DNA binding specificities but differ in their transactivation efficiencies, depending on their specific DNA sequence and the promoter context (Chinenov and Kerppola 2001). The first component of AP-1 protein is c-Jun (Maki et al. 1987), which plays a critical role among various functions of AP-1 transcription factors because it can interact with many other members of the AP-1 family to exert its function (Chinenov and Kerppola 2001). Because c-Jun is located at the end of the pathway that transmits signals for cell proliferation, differentiation, transformation, and apoptosis (van Dam and Castellazzi 2001), it is important to elucidate whether c-Jun is a transcription factor of the bcl-2 gene in normal human endometrium.
In the past 10 years, Biacore analysis has become the standard technique for studying real-time biomolecular interactions such as DNA–protein interactions (Fivash et al. 1998). The Biacore 2000 system consists of a microfluid cartridge that forms four separate channels, called flow cells, on the surface of a sensor chip. One reactant, referred to as the ligand, is co-valently coupled to the sensor chip surface; the other reactant, referred to as the analyte, flows past this surface in solution. By making use of surface plasmon resonance techniques, the Biacore system can measure local changes in refractive index near the surface of a sensor chip when analyte and ligand bind. A fluid sample can be injected either separately over each flow cell or serially over all flow cells or over combinations of flow cells. By introducing the sample in a flow, the concentration of the sample over the surface is maintained at a constant level (Fisher et al. 1994). The covalent nature of the bound ligand allows regeneration of the surface for many repetitive measurements of analyte binding. One of the unique advantages of this technique is that it allows rapid analysis of several samples with a very low concentration of analyte in a small volume. It appears that the Biacore technique is feasible for the study of nuclear protein–DNA interactions in crude nuclear extracts.
Our IHC of c-Jun showed that its cyclic expression pattern, peaking in expression level during the proliferative phase, was similar to that of bcl-2 in human endometrial glandular cells. However, IHC cannot determine with certainty the specific binding of c-Jun to AP-1 motifs from the bcl-2 promoter or exclude the possibility of the specific binding of c-Jun to promoters of other genes in endometrial glandular cells. This study using the Biacore system was carried out to elucidate the direct binding between c-Jun and AP-1 motifs of the bcl-2 promoter in human endometrium throughout the menstrual cycle.
Materials and Methods
Uterine Endometrial Samples
Endometrial tissue samples were obtained from patients undergoing hysterectomy for benign disease (proliferative phase, n = 5; secretory phase, n = 7). The patients (30–48 years old, average 43 years) showed normal menstrual cycles and had received no hormonal therapy for at least three menstrual cycles before surgery. Small tissue blocks were obtained from the fundus within the uterine cavity containing the endometrium and were used for IHC. Nuclear proteins were extracted from the remaining endometrium for Biacore analysis. The menstrual phase was determined on the basis of both menstrual history and the results of endometrial histology. Informed consent was obtained from each patient before the surgery, using protocols approved by the Ethical Committee of Osaka Medical College.
Immunohistochemistry
Part of the fresh tissue samples was rapidly frozen in liquid nitrogen and embedded in Tissue Mount (Chiba Medical; Saitama, Japan). The frozen blocks were cut serially into 6–7-μm sections with a cryotome (Microm; Heidelberg, Germany) at −20C. Serial cryosections placed on poly-
Nuclear Proteins from Endometrial Cells
Separation of the uterine endometrial glandular and stromal cells has been previously described (Otsuki et al. 1994). In brief, part of the tissue samples were dispersed by incubation of minced tissues in Mg2+- and Ca2+-free Dulbecco's modified (DPBS), pH 7.4, containing collagenase (2 mg/ml, 15 U/mg), DNase (0.2 mg/ml, 1500 U/mg), and 10% fetal calf serum for approximately 20 min at 37C with constant agitation. Disaggregated endometrial stromal cells were separated from glands by sequential filtration through nylon mesh filters with pore sizes of 300 μm and 40 μm. Endometrial glands remained largely intact and were retained by the sieve, while stromal cells passed through into the filtrate. The endometrial glands were allowed to sediment twice at 37C in DPBS for 30 min each before trypsin treatment. The endometrial glands were dispersed by passing them through a 26G needle.
The nuclear extracts were prepared from the dispersed stromal and glandular cells according to a previously reported protocol (Dong et al. 1999). All manipulations were carried out at 4C. Briefly, the dispersed cells were harvested and suspended in five-pellet volumes of a hypotonic buffer containing 10 mM HEPES (pH 7.9, 4C), 1.5 mM MgCl2, 0.2 mM EDTA, 10 mM KCl, 0.1 mM PMSF, 0.5 mM DTT, and 25% (v/v) glycerol. Then the cells were incubated in three-pellet volumes of the same buffer on ice for 15 min. The nuclei were collected by centrifugation at 3300 × g for 15 min. Pelleted nuclei were resuspended in 0.5-pellet volume of a low-salt buffer containing 10 mM HEPES (pH 7.9, 4C), 1.5 mM MgCl2, 0.2 mM EDTA, 10 mM KCl, 0.1 mM PMSF, 0.5 mM DTT, and 25% glycerol (v/v). A 0.5-pellet volume of a high-salt buffer, prepared by replacing 10 mM KCl in the low-salt buffer with 1.2 M KCl, was then added dropwise while mixing. The nuclei were incubated at 4C for 30 min with constant agitation. The nuclear lysate was cleared by centrifugation at 100,000 × g for 40 min and dialyzed against a buffer containing 10 mM HEPES (pH 7.9, 4C), 100 mM KCl, 0.1 mM PMSF, 0.5 mM DTT, and 20% glycerol (v/v). Aliquots of this lysate were frozen in liquid nitrogen and stored at −80C. The protein concentrations were determined using a DC Protein Assay Kit (Bio-Rad Laboratories; Hercules, CA). In general, about 50–120 μg of nuclear protein could be obtained.
Biotinylated Promoter Fragments
Two double-stranded oligonucleotides were used in the present experiments. The upper strand sequence information was obtained from GenBank NIH (Bethesda, MD) and arbitrarily named as follows: AP-1x 5′-CGACTCCTGATTCATTGGGA-3′ (–645 to −625) and AP-1y 5′-TCGGAGTTTAATCAGAAGAG-3′ (–993 to −973). Biotinylated and purified bcl-2 promoter fragments were obtained from Invitrogen (Carlsbad, CA). Only the 5′ ends in the upper strands were biotinylated. To make double-stranded oligonucleotides, the biotinylated upper strand and the non-biotinylated down strand oligonucleotides were added at a molecular ratio of 1:3 to a standard PCR buffer containing 25 mM MgCl2. Then the oligonucleotides were annealed by heating at 98C for 10 min, cooling to 50C at a rate of about 2C/min, and then chilling on ice. The double strands generated in this way could be subcloned and sequenced (data not shown). Control DNA was prepared from biotinylated bacteriophage Hinf II fragments from Roche Molecular Biochemicals (Tokyo, Japan). The DNA was sonicated for 20 min, and passed through a column to obtain DNA fragments of less than 50 bp.
Surface Plasmon Resonance (SPR)
The SPR measurements were conducted using the Biacore 2000 system, BIA evaluation software 3.0, Sensor Chip SA, and HBS buffer (pH 7.4) containing 10 mM HEPES, 150 mM NaCl, 3 mM EDTA, and 0.005% surfactant P20 (from Biacore AB, Uppsala, Sweden). The measured SPR value is expressed in arbitrary resonance units (RU), where a change of 10–4 degrees is equivalent to a change of 1 RU. RU is a function of the change in surface concentration of molecules. Approximately 1000 RU is equivalent to a change in surface concentrations of ∼1 ng/mm2 protein and ∼0.8 ng/mm2 DNA. The system has an effective dynamic range from 3 to 4 RU to 30,000 RU. All experiments were performed at 25C according to the manufacturer's instructions and the methods reported by Galio et al. (1997, 1999). To immobilize the biotinylated DNA fragments to the sensor chip, 0.1 μM of the annealed oligonucleotides in HBS buffer containing 150 mM NaCl was injected at a rate of 5 μl/min over each flow cell.
HBS buffer with 25 ng/μl poly [d (I-C)] was used to prepare the nuclear proteins for injection. Human recombinant c-Jun (Promega Biosciences; Madison WI) was injected at a concentration of 10 or 20 footprinting units/ml, corresponding to 6.4 or 12.8 ng/μl of nuclear protein. For samples from human endometrium, 1.0–7.0 μl of the crude nuclear extracts was diluted to 10 ng/μl or 20.0 ng/μl. The samples were incubated at 24C for 15 min. The anti-c-Jun antibody was from Santa Cruz Biotechnology. Normal rabbit IgG (Chemicon International; Temecula, CA) was used as a negative control antibody. Each association/dissociation cycle was performed at a constant flow rate of 30 μl/min. For protein-DNA binding reactions, each sample was injected over the flow cells for 5 min, followed by a constant flow of HBS buffer for 5 min to observe dissociation of bound proteins. Then 20 ng/μl normal rabbit IgG or anti-c-Jun antibody in HBS buffer was injected in the same manner. At the end of each cycle, bound proteins were removed by two-pulse injections of 1.5 M NaCl for 30 sec each to regenerate the chip. Sensorgrams were recorded automatically and adjusted to zero baseline level by subtracting the response recorded immediately before the injection of each sample. Comparison between sensorgrams was carried out by subtracting the responses in control flow cell, which is the lowest sensorgram in each figure on the right panel of Figures 2–4.
Results
Immunohistochemistry
Immunoreactivity of c-Jun could be detected in sections from all 12 endometrial samples collected during the proliferative through the secretory phase of menstrual cycle (Figure 1). When multiple sections of the same uterine endometrium were examined, an identical staining pattern was observed in different sections. Intense immunostaining of c-Jun could be detected in sections from the proliferative phase (Figure 1A). Expression of c-Jun was localized on the nuclei of both glandular and stromal cells, although the signal appeared stronger in the glandular cells. However, the intensity of expression was markedly decreased in endometrial sections from seven samples from the secretory phase (Figure 1B), especially in the glandular cells. Although many of the glandular nuclei did not exhibit positive immunoreactivity, only a portion of the nucleus was stained in many others. In all sections examined, no definite immunoreactivity could be observed outside of nuclei. The expression pattern observed in our study is consistent with what has been previously reported (Salmi et al. 1998), showing intense staining during the proliferative phase and a weakened signal in nuclei of the glandular cells before menstruation.
Surface Plasmon Resonance (SPR)
The amounts of oligonucleotides immobilized were 300 RU (AP-1x300) or 600 RU (AP-1x600) for AP-1x, and 300 RU (AP-1y300) for AP-1y, to obtain the most reproducible results. The first flow cell was immobilized with similar amount of control DNA. Injection of the human recombinant c-Jun 6.4 ng/μl resulted in a rising peak for both AP-1x and AP-1y (Figures 2B and 2B'). Afterwards, a rapidly rising curve could be detected during injection of the rabbit anti-human c-Jun antibody instead of the normal rabbit IgG (Figures 2A and 2A'). Then the rising curve descended very slowly when the running buffer was flowing over, indicating a high association rate of the bound molecules. Increasing the concentration of the injected nuclear protein to 12.8 ng/μl showed similar results (Figures 2C and 2C). It is noteworthy that AP-1y more efficiently bound the purified recombinant c-Jun than did AP-1x, because a similar binding value between AP-1x300 and AP-1y300 could be detected when the amount of c-Jun was elevated from 6.4 ng/μl to 12.8 ng/μl (Figure 2C). The above results demonstrate that specific binding of the purified c-Jun to AP-1x and AP-1y can be effectively detected with the Biacore 2000 system.
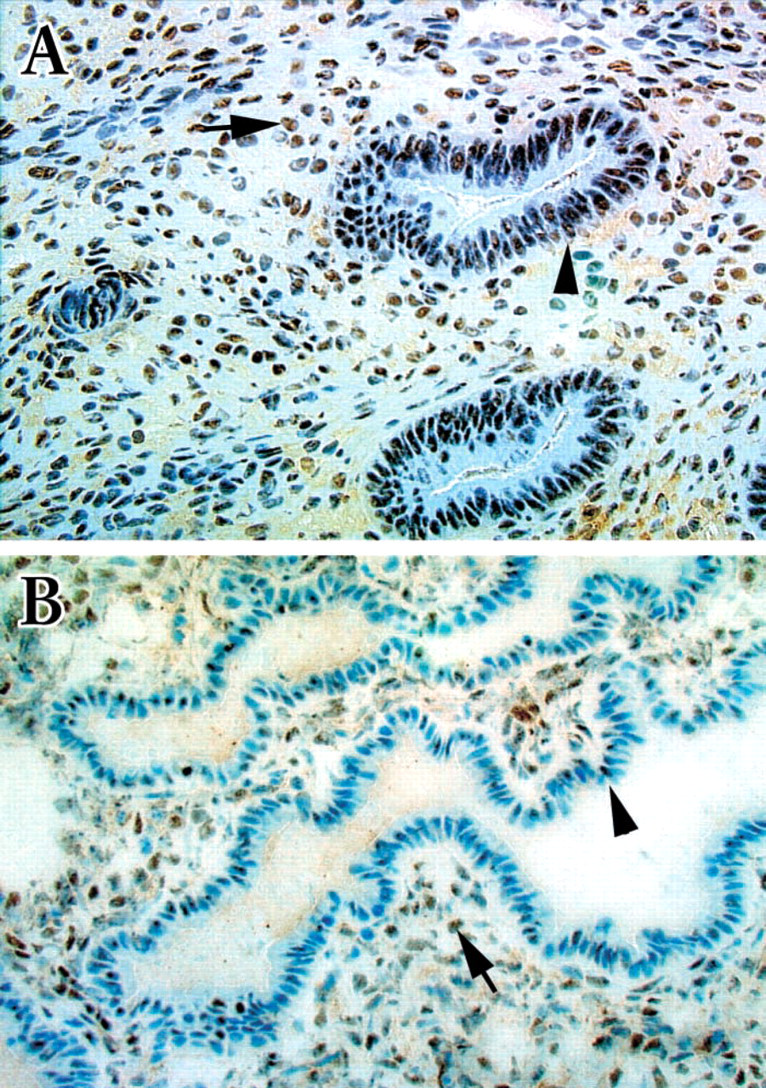
Immunohistochemical staining of normal human endometrium with anti-c-Jun antibody. Immunoreactivity can be detected in both the glandular (arrowheads) and stromal (arrows) nuclei of sections from either the proliferative (
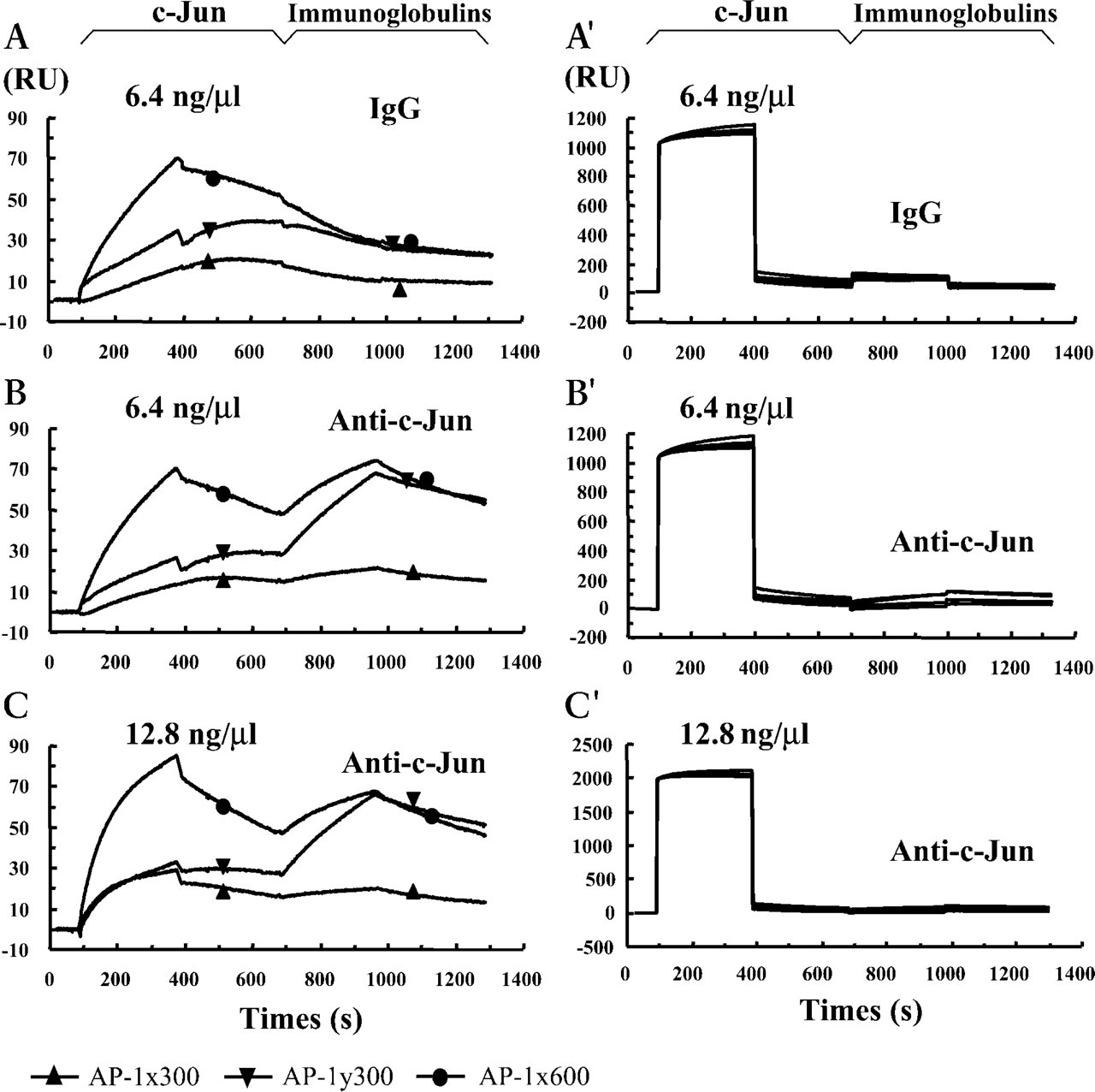
Control sensorgram depicted from injection of the purified recombinant human c-Jun followed by the anti-c-Jun antibody (Anti-c-Jun) or the normal rabbit IgG (IgG). Sensorgrams
The crude nuclear extracts were prepared from five samples obtained in the proliferative (menstrual days 7, 10, 11, 12, and 14) and seven samples in secretory (menstrual days 17, 19, 20, 20, 21, 22, and 24) phases, respectively. Each sample was measured more than three times and reproducible results were obtained each time. Typical results are shown in Figures 3 and 4. After the crude nuclear extracts 10 ng/μl from the proliferative glandular cells were passed over the flow cells, injection of the anti-c-Jun antibody 20 ng/μl resulted in sharply rising curves for both AP-1x and AP-1y (Figures 3A and 3A'). Increasing the concentration of injected nuclear proteins to 20 ng/μl yielded more intense association (Figures 3B and 3B'). Similar to the purified c-Jun (Figure 2B), a stronger association was detected in the flow cells coupled with AP-1y than in those coupled with AP-1x. Even in this case the response was rarely detectable after injection of the proliferative nuclear extract prepared from the stromal cells (Figures 3C, 3C, 3D, and 3D') of the same tissue sample. These results indicate that, in the proliferative stromal cells, although the nuclear c-Jun exists (Figure 1A) it is not available for binding to the AP-1 motifs in the bcl-2 promoter, because of the fact that the purified c-Jun could bind the AP-1 motifs very efficiently (Figure 2). When the secretory nuclear extracts were used in the assay, neither the glandular (Figures 4A, 4A', 4B, and 4B') nor the stromal samples (Figures 4C, 4C, 4D, and 4D') showed apparent binding activity after injection of the anti-c-Jun antibody, although the nuclear c-Jun could be detected (Figure 1B) by IHC. Our results showed that, during the menstrual cycle, only the glandular c-Jun in the proliferative nuclear extracts could bind to the two AP-1 motifs in the bcl-2 promoter, indicating spatial and temporal binding activities.
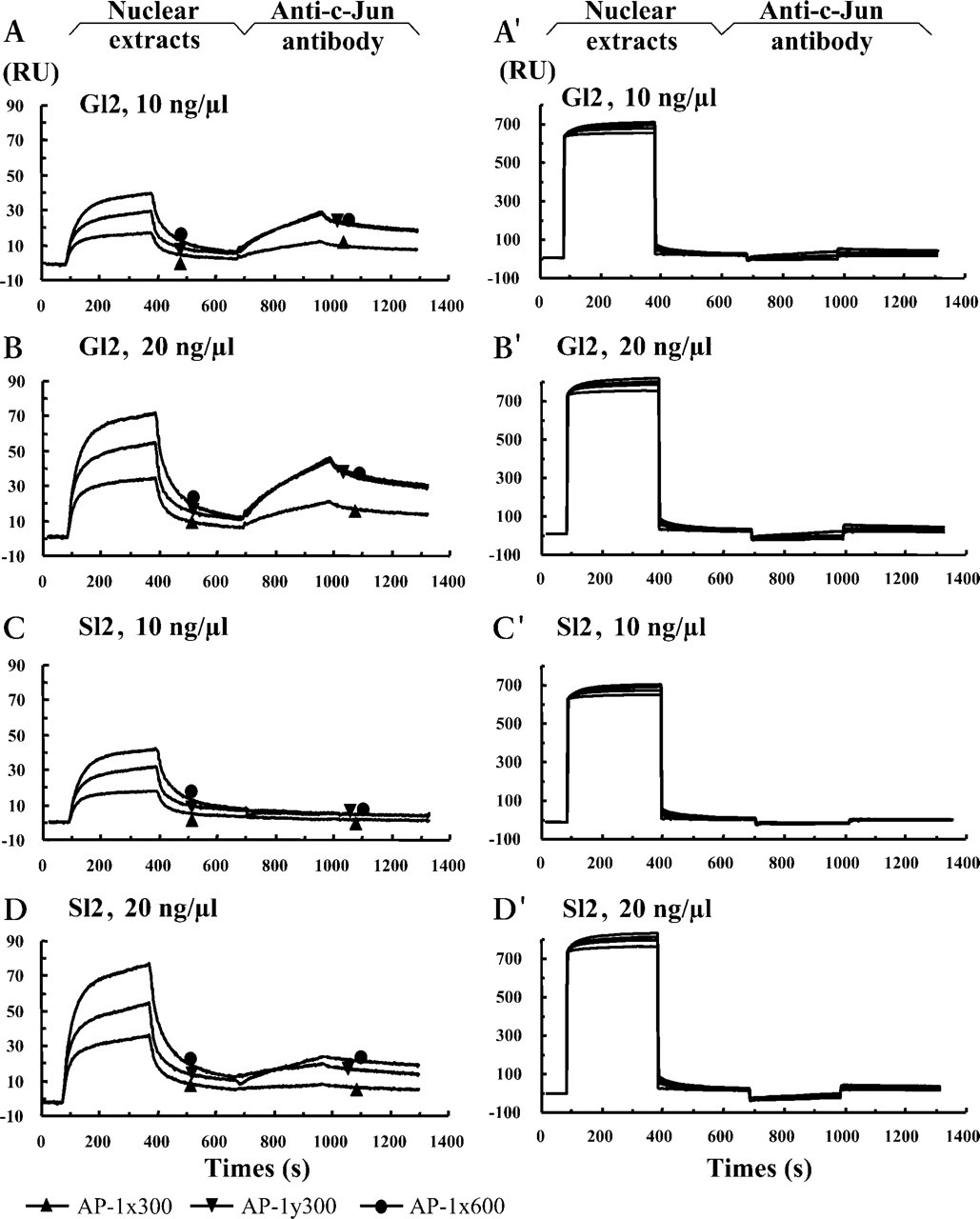
Binding activities of the glandular (G12) and stromal (S12) nuclear extracts from the proliferative phase, menstrual day 12. Sensorgrams
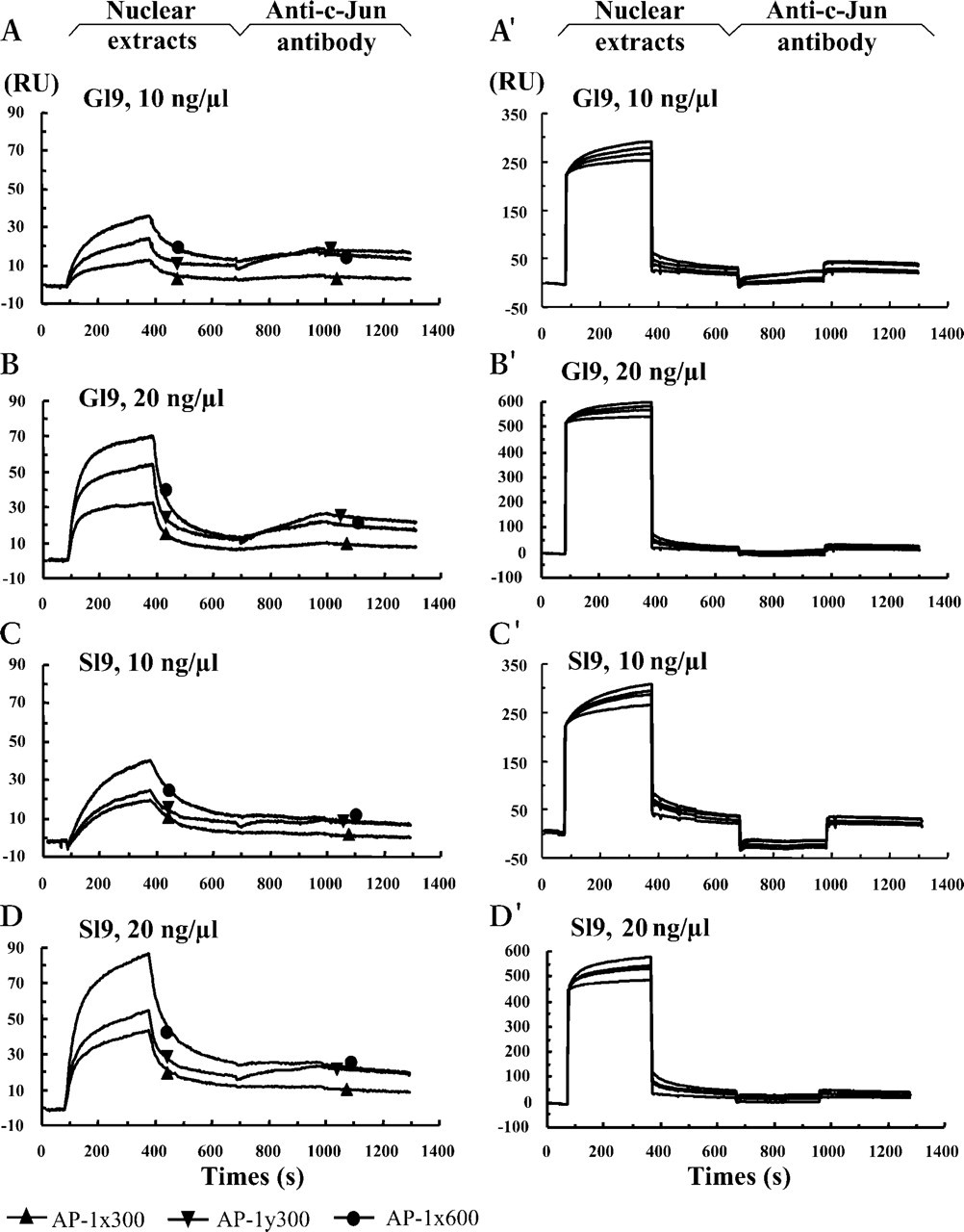
Binding activities of the glandular (G19) and stromal (S19) nuclear extracts from the proliferative phase, menstrual day 19. Sensorgrams
Discussion
Regulation of transcription is the dominant mechanism for the control of gene expression that involves various protein-DNA interactions. The electrophoretic mobility shift assay (EMSA) is one of the conventional techniques employed to obtain in vitro evidence of nuclear protein-DNA interactions. However, because only a small amount of glandular nuclear extract can be collected from each uterine endometrium, it is difficult to use EMSA for screening candidate transcription factors. Our experiments showed that the Biacore technique is feasible for solving this problem. The Biacore technique can be used to monitor relative affinities of proteins binding to immobilized DNA fragments (Fisher et al. 1994; Buckle et al. 1996). The Biacore 2000 used in this study permits detection in up to four flow cells, which means that one injection of nuclear extract could enable the determination of binding activities of several different oligonucleotides coupled to separate flow cells. Therefore, while the amount of nuclear extract required for each measurement could be reduced still further, efficient observation could now be achieved at this stage by the current technique. In terms of protein-DNA interactions, however, most researchers observed binding affinities and kinetics between biotinylated oligonucleotides immobilized on a SA sensor chip and purified molecules (Buckle et al. 1996). It should be noted that there is a good correlation between results from gel shift assay and Biacore using crude nuclear extracts from cultured cells (Galio et al. 1997,1999). In the present study we examined crude nuclear extracts from both glandular and stromal cells separated from the human endometrium. It is not known how many molecules in the crude nuclear extracts will bind to the coupled oligo-nucleotides. Therefore, we devised several methods for detecting specific binding of a known oligonucleotide to its target molecule(s) in nuclear extracts. In our experiments, the mixed DNA fragments from a bacteriophage were coupled to the first flow cell as a reference surface. Although still unconfirmed, these DNA fragments may not specifically bind to molecules in mammalian nuclear extract. It could be useful for distinguishing between true binding events, nonspecific binding of nuclear proteins, and signals that are due to differences in refractive index between samples. In addition, normal IgG from rabbit was used to distinguish between specific binding and nonspecific binding of the anti-c-Jun antibody.
To elucidate whether c-Jun could transcriptionally regulate expression of the bcl-2 gene in a normal human uterine endometrium, we first examined the expression pattern of c-Jun in both the proliferative and secretory phases of the menstrual cycle by IHC. Intense immunoreactivity of c-Jun could be detected in the endometrial glandular cells during the proliferative phase, whereas its immunoreactivity was markedly decreased in the secretory glandular cells. The expression pattern of c-Jun was consistent with that reported by Salmi et al. (1998), showing intense staining and increased amount of expression during the proliferative phase and a decreased amount of expression and immunoreactivity in nuclei of the glandular cells before menstruation. It is worth noting that this expression pattern is well associated with the cyclic expression of bcl-2 from the proliferative to the secretory phase as described previously (Otsuki et al. 1994).
The oncogene product c-Jun has been implicated in mediation of a wide variety of signals by transcribing them into expression of proteins encoded by specific genes (Shaulian and Karin 2001). Therefore, c-Jun may sometimes cause contrasting effects, such as cell death or survival, depending on the expression of different genes. It is well known that c-Jun determines the proper proliferation and differentiation of primary human keratinocytes, based on the fact that lack of c-Jun results in formation of an aberrant epithelium containing a decreased number of proliferating cells and few cell layers (Szabowski et al. 2000). In contrast to cell survival, the hepatoblasts in mouse embryos lacking functional copies of c-jun show enhanced apoptosis and impaired proliferation in vitro, while fibroblasts exhibits severely reduced growth rates in culture (Eferl et al. 1999). The identification of additional c-Jun regulator and effector genes that are important for cell death or survival will help in elucidating not only the mechanism of c-Jun regulation but also the functions of c-Jun on various cell types in different tissue contexts.
It is important to determine whether the association of c-Jun with AP-1 motifs from the bcl-2 promoter is regulated in a spatial and a temporal manner. The c-Jun was expressed in the nuclei of stromal cells during both proliferative and secretory phases and in those of a few glandular cells during the secretory phase, as well as in those of glandular cells during the proliferative phase. However, sharp peaks after injection of the anti-c-Jun antibody, indicating the specific binding of c-Jun to AP-1 motifs, were not observed in nuclear extracts from stromal cells during both phases and from glandular cells during the secretory phase. The difference between IHC and binding activities of AP-1 motifs to c-Jun in different nuclear extracts may be explained as follows. First, the function of c-Jun may be different between the proliferative and secretory phases and even between the glandular and stromal nuclei of the same menstrual phase. Of course, the difference in binding activities may be caused by different phosphorylation status of c-Jun protein in nuclear extracts (Rutberg et al 1999). Second, other factors in the nuclear extracts may influence specific binding of c-Jun to AP-1 motifs. It is very important to remember that c-Jun can interact with other members of AP-1 factors, such as JunD or ATF-1, to exert its effects (Angel and Karin 1991). Other transcription factors, especially steroid hormone receptors such as the ER (Dong et al. 1999), may be crucial for c-Jun to function properly in human endometrium.
Both the immunohistochemistry and the Biacore analysis clearly showed that c-Jun contained in the nuclear extracts of glandular cells during the proliferative phase specifically bound to two AP-1 sites in the bcl-2 promoter region. These results explain why bcl-2 is expressed cyclically in proliferative glandular cells. Further studies with transcriptional regulation and upstream signal transduction may provide new insights to understand the regulation of proliferation, differentiation, and apoptosis of cells in normal human endometrium.
Footnotes
Acknowledgements
Supported in part by a Grant-in-Aid for General Scientific Research from the Ministry of Education, Culture, Sports, and Technology of Japan (no. 10671576).