
Abstract
KLH (Keyhole limpet hemocyanin) is highly immunogenic, and crossreactive epitopes occur widely in nature. In schistosomiasis, infected hosts generate antibodies reactive with KLH. This is of diagnostic importance but we lack detailed information on the immunogen-carrying molecules and their distribution in the worm. We used anti-KLH antibodies to localize cross-reacting epitopes in the various developmental stages of the parasite in experimental Schistosoma mansoni infection. The staining results show KLH crossreactivity in the life stages of the parasite. By immunoblotting we show that KLH-crossreactive antigenic epitopes in the parasite eggs are carbohydrates, also recognized by antibodies against soluble schistosome egg antigens. The localizations in the larval stages and in adult worms suggest that crossreacting antigenic epitopes are secretory products.
Keywords
K
Materials and Methods
S. mansoni Infection in Mice
The schistosome life cycle was maintained as described previously (Linder et al. 1991). Eight-week-old NMRI mice were infected percutaneously with about 100 S. mansoni cercariae (Puerto Rican strain). At 8–10 weeks after infection the mice were sacrificed, intravascular worms recovered by perfusion with RPMI medium, and liver and intestine removed for immunohistological staining experiments. Isolated schistosome eggs were hatched in water and emerging miracidia used for infection of Biomphalaria glabrata snails.
Antigen Preparations
Frozen sections of adult schistosomes and egg granuloma-and worm-containing tissues mounted on microscope slides were fixed in acetone at room temperature (RT) for 10 min. Paraffin sections of formalin-fixed material were also used as antigen.
Preparations of cercariae, to be used as target antigen, were made in two ways. Living cercariae were added in aliquots of 10 μl on microscope slides, coated with human skin lipids (Stirewalt 1978), dried at RT, and fixed in acetone for 10 min. The other preparation of cercariae was fixed in 4.5% paraformaldehyde and washed three times in PBS before use in immunofluorescence (IF) staining.
For some experiments, suspensions of isolated eggs were allowed to “hatch” in water. Released miracidia and “hatch fluid” were used as antigen for IF staining. Schistosomula were prepared as described (James and Taylor 1976) and fixed in 4.5% paraformaldehyde. Soluble egg antigen (SEA), prepared essentially as described (Carter and Colley 1978), and KLH (H-7017; Sigma, St. Louis, MO) were used in Western blotting.
Antisera and Lectins
Commercially available rabbit anti-KLH antibodies (H-0892; Sigma) were used at a 1:40 dilution for IF staining. A mouse monoclonal antibody (MAb) against paramyosin (Pearce et al. 1988), kindly provided by Dr Allan Sher (NIAID, NIH; Bethesda, MD) was used as a marker to show that the parasite surface was intact. Rabbit anti-SEA sera were raised by immunization with three monthly doses of 0.1 and 1 mg of SEA. Fluorescein isothiocyanate (FITC)-labeled Arachis hypogaea lectin (PNA) (Sigma L-7381), specific for terminal β-
Immunofluorescence Microscopy
Staining was performed as described previously (Thors and Linder 1998). Frozen sections or formalin-fixed, paraffin-embedded tissue sections were incubated with rabbit anti-sera and anti-immunoglobulin (Ig)–FITC conjugate (sheep anti-rabbit Ig) (National Bacteriological Laboratory; Stockholm, Sweden). Sections of adult worms and liver tissue with eggs and worms were treated with 20 mM sodium periodate in 0.1 sodium acetate buffer, pH 4.5, for 1.5 hr in the dark at RT. The reaction was stopped by adding an equal amount of 50 mM sodium borohydride for 30 min.
IF microscopy was performed using a Leica DMRB fluorescence microscope equipped with filters for FITC fluorescence essentially as described previously (Linder et al. 1991). For photography, an AxioCam digital microscope camera (Carl Zeiss; Oberkochen, Germany) was used and Openlab software (Improvision; Coventry, UK) for Apple Macintosh computer G4 using McOS9.1 system and Photoshop 6 (Adobe; San Jose, CA) for image processing.
SDS-PAGE and Western Blotting
SEA (48 μg per gel) and KLH (20 μg per gel) were separated in 10.6% SDS-PAGE with a 3% stacking gel under reducing or non-reducing conditions and transferred onto nitrocellulose membranes (Laemmli and Favre 1973). The membranes were incubated in PBS containing 5% non-fat milk for 30 min. Serum samples were tested at a dilution of 1:200. Horseradish peroxidase (HRP)-conjugated anti-rabbit Ig (NA9340; Amersham, Poole, UK) diluted 1:1000 were used. PBS with 0.1% bovine serum albumin and 0.1% Tween-20 was used as diluent. The strips were developed in 4-chloro-1-naphthol 0.5 mg/ml (Sigma; C-6788) in triethanol-amine-buffered saline, pH 7.5, containing 0.0125% H2O2. Molecular weight standards were from Bio-Rad (161–0324; Hercules, CA).
For deglycosylation of the antigens on nitrocellulose membranes, incubation with periodate was performed as described (Hamilton et al. 1999).
Results
By immunofluorescence, rabbit anti-KLH antibodies reacted with the different developmental stages of the parasite (Table 1). The reaction was similar in paraffin and frozen sections. Schistosome eggs in both the liver and the gut wall were positive and the staining appeared rather homogeneous. Interestingly, the peri-oval tissue showed various degrees of diffuse staining in the close vicinity of the autofluorescent eggshells (Figures 1A and 1B). Reactive material was frequently seen as granular deposits in the periphery of granulomas (Figure 1B). Eggs, hatched in vitro, showed anti-KLH reactivity mainly at the internal surface of the eggshell, and distinct staining of the miracidial surface could be seen (Figure 1C). The cercarial surface showed strong reactivity (Figure 2A) and the cercarial secretions also reacted with anti-KLH antibodies (Figures 2B and 2C). Cercariae transformed into schistosomules retained strong surface reactivity with anti-KLH antibodies (Figure 3A). Control staining with anti-PM antibodies indicated that the schistosomular surface was intact (Figure 3B). In adult worms in both frozen sections (Figure 1E) and paraffin sections (Figures 4A–4D), secretory ducts were seen in male and female worms. In addition, both the ovary and the testis were stained (Figures 4B and 4C). In female worms, connective tissue stroma stained strongly, e.g., as septa surrounding groups of vitelline cells (Figures 4A and 4B). In sections treated with periodate, staining was negative or significantly weaker (Figure 4E). Control sections stained with anti-SEA antibodies showed a similar staining pattern as anti-KLH antibodies (Figures 4F–4J). Thin fluorescent threadlike structures seemed to be extensions of subtegumental cells localized close to strongly stained ducts and ended as knobs located at the center of tubercles in males (Figures 4D and 4H).
Immunofluorescent reactivity of rabbit anti-KLH antibodies with different life stages of S. mansoni

Frozen and paraffin sections.
Whole mounts were used as antigen.
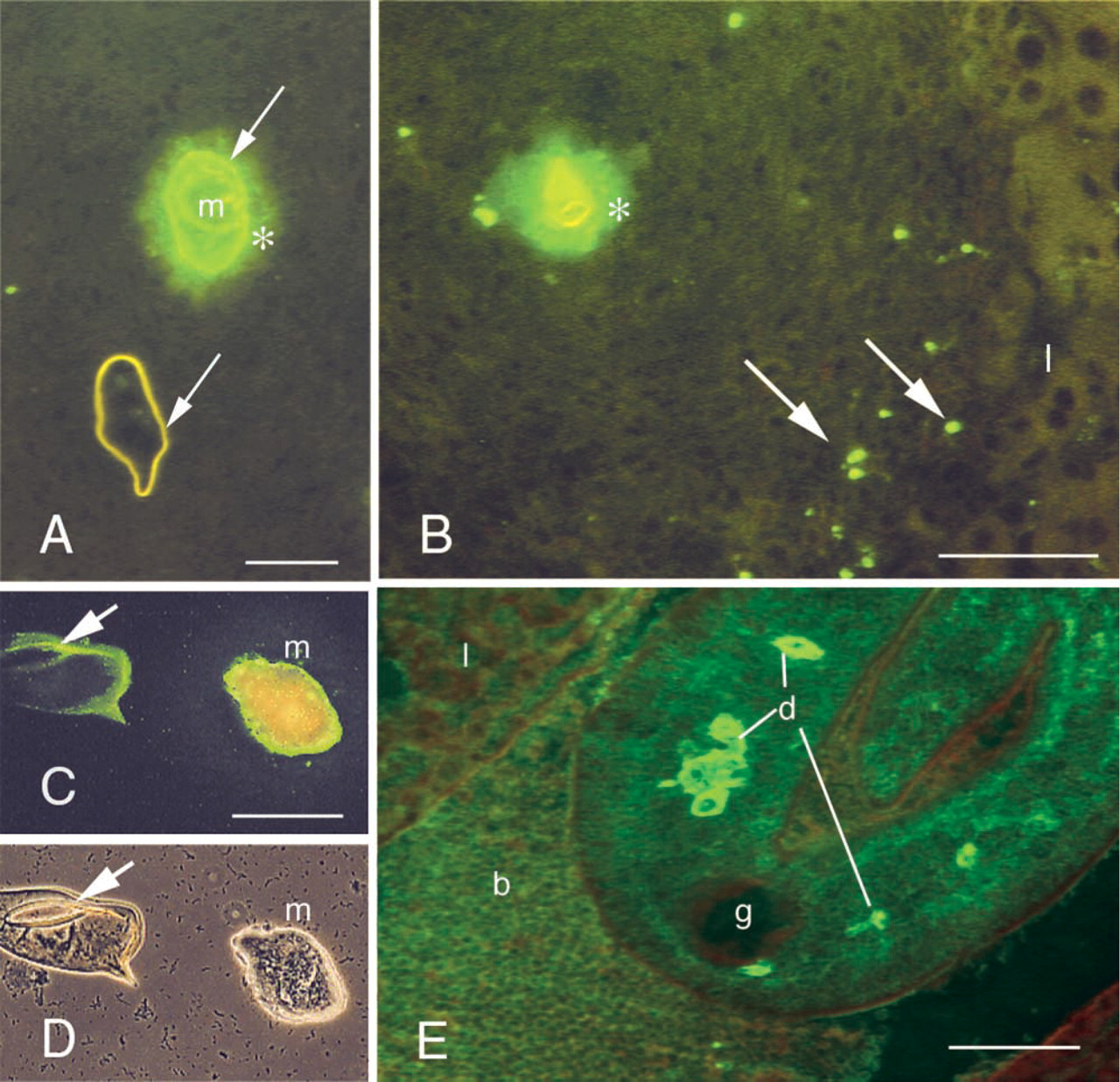
Anti-KLH reactivity with S. mansoni egg granuloma in indirect IF (
In Western blotting, rabbit anti-KLH and anti-SEA antibodies appear to recognize in part the same KLH components (Figure 5). However, all reactivity of anti-SEA antibodies with KLH was abolished after perio-date treatment, whereas only a slight reduction in the staining intensity of KLH bands in the 50–60-kD region was seen after staining with anti-KLH antibodies. When tested on periodate-treated SEA, all anti-KLH reactivity disappeared. A major ∼100-kD band in non-reduced SEA samples disappeared on reduction, and in the ∼50-kD region a band recognized by anti-SEA antibodies was periodate-sensitive.
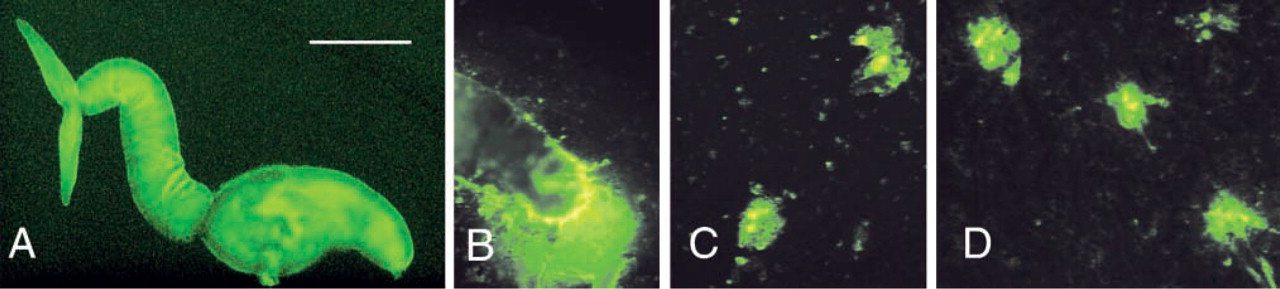
Anti-KLH reactivity of the surface of a paraformaldehyde-fixed cercaria (
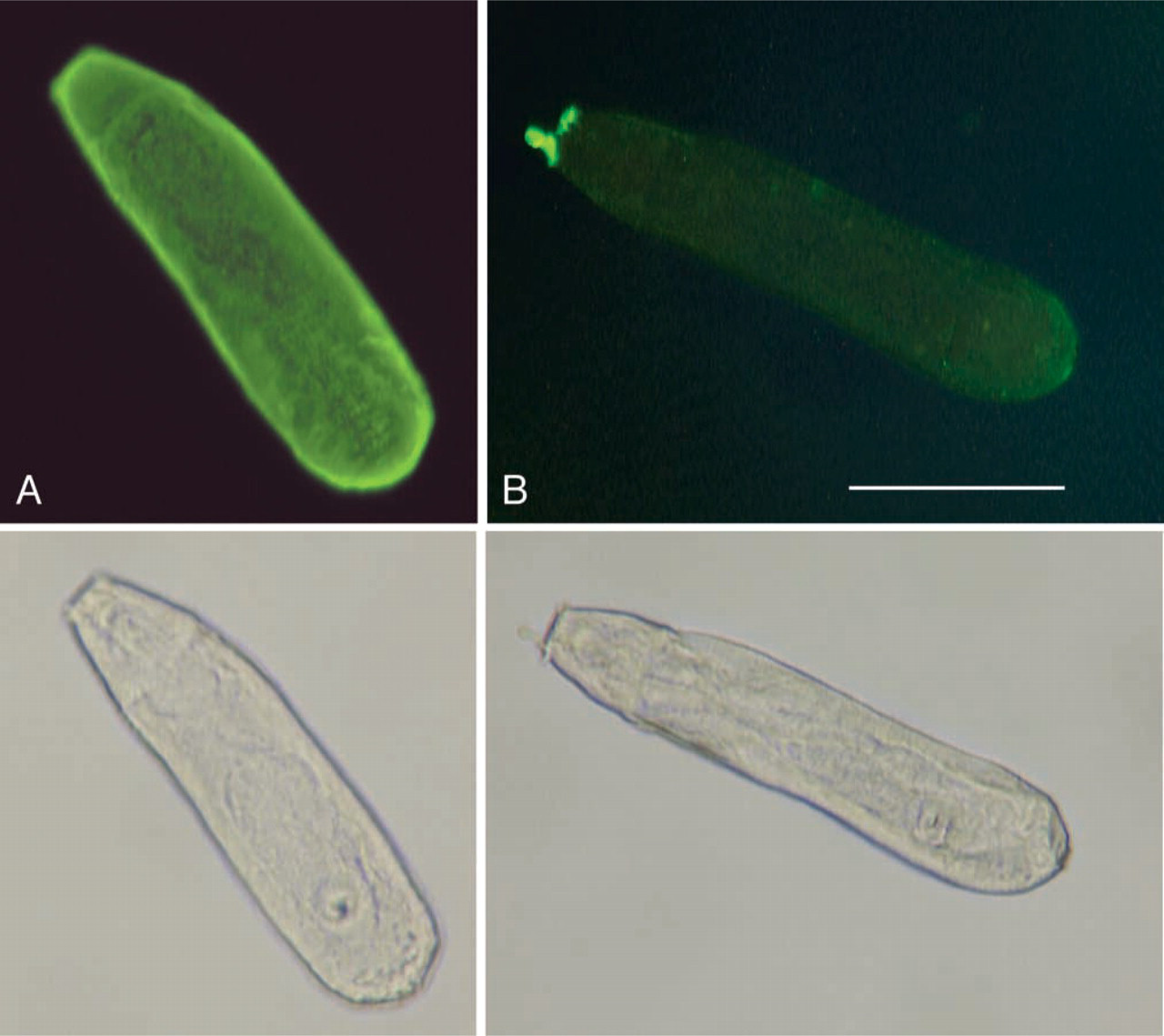
Anti-KLH reactivity with the intact (paraformaldehyde-fixed) schistosomular surface (
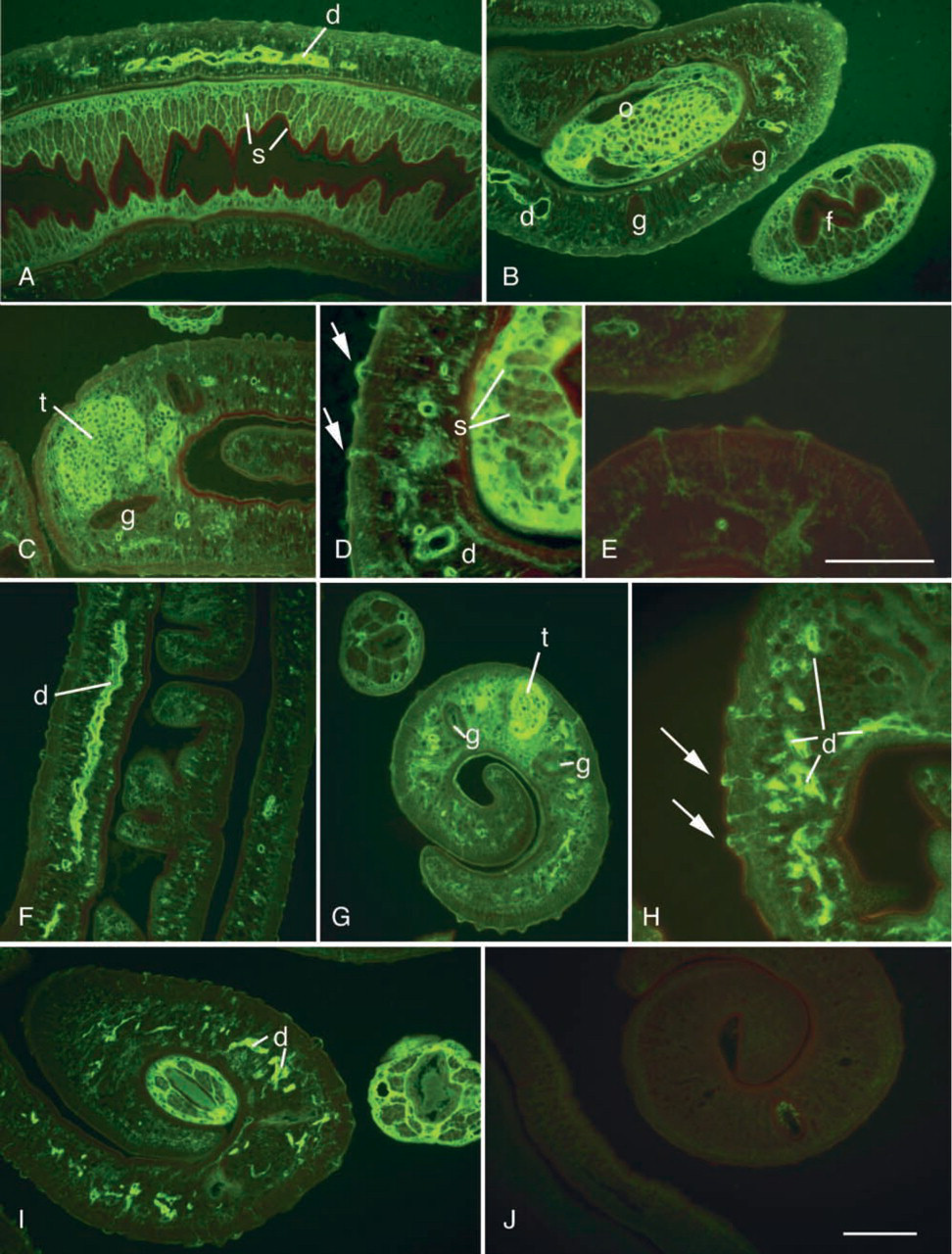
Anti-KLH (
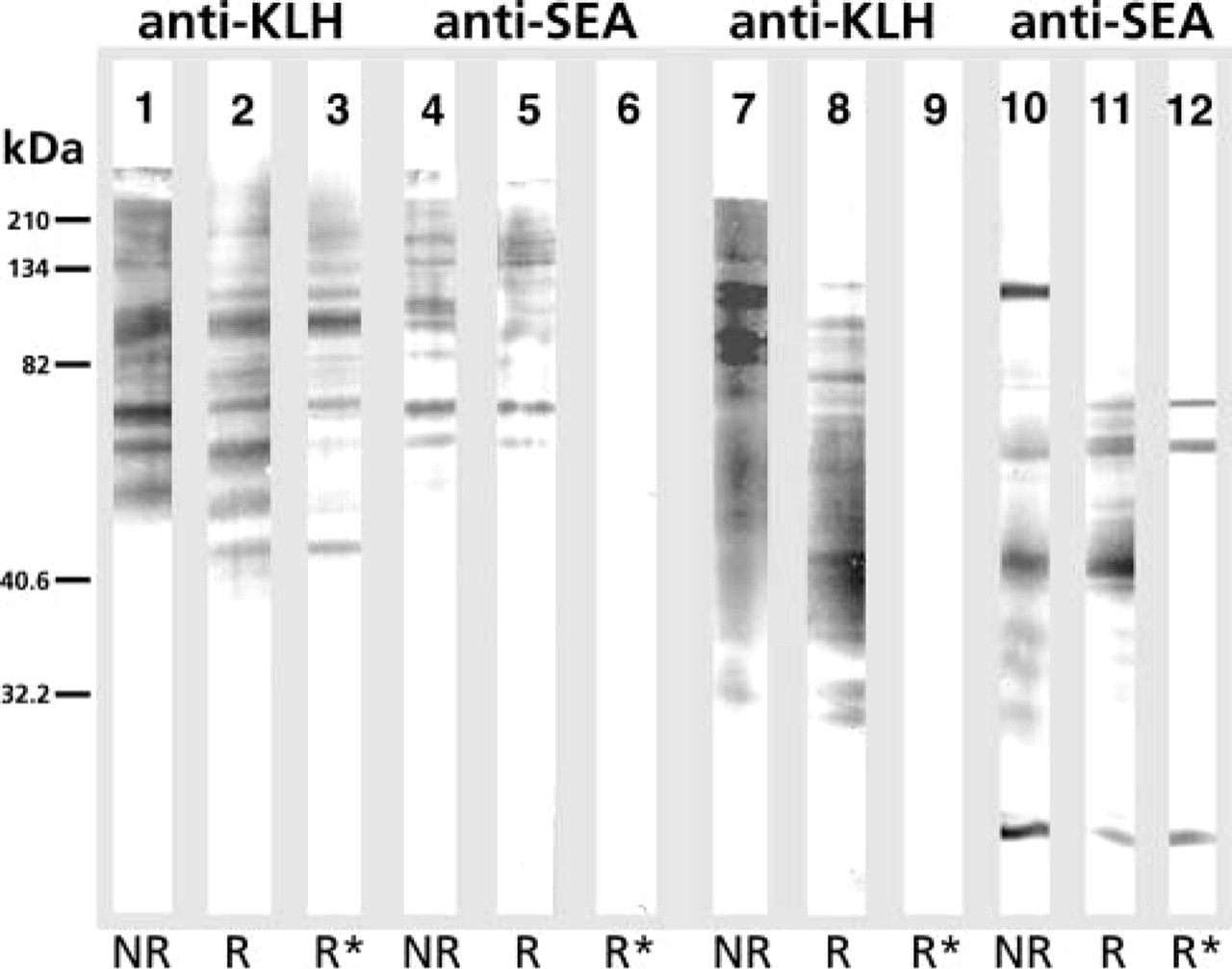
Reactivity of rabbit anti-KLH (1–3, 7–9) and rabbit anti-SEA (4–6, 10–12) antibodies with KLH (1–6) and SEA (7–12) in Western blotting after separation in PAGE 10.6% gel under non-reducing (NR) (1, 4, 7, 10), reducing (R) (2, 5, 8, 11) conditions, and after periodate treatment under reducing conditions (R∗) (3, 6, 9, 12). Reactivity of anti-SEA (6) but not anti-KLH antibodies (3) against KLH was completely abolished after periodate treatment, whereas the reactivity of anti-KLH antibodies (9) was completely abolished when periodate-treated SEA was used as antigen. Molecular weight markers: myosin (210,000), β-galactosidase (134,000), bovine serum albumin (82,000), carbonic anhydrase (40,600), soybean trypsin inhibitor (32,000) are shown at left.
Discussion
KLH elicits a strong immune response in vertebrates. Interestingly, shared antigenic epitopes occur in schistosomes. These common epitopes of schistosomes are major immunogens in schistosomiasis, which is reflected by the use of KLH as a diagnostic antigen. In the present study we used anti-KLH antibodies to localize the KLH-crossreacting parasite components that may give rise to an immune response in schistosomiasis.
The major crossreacting immunogenic epitopes of KLH have been demonstrated in preparations of schistosome eggs (“soluble egg antigens,” SEA) (Hamilton et al. 1999). Both in Western blotting and in sections of the parasite, the target antigens recognized by anti-KLH antibodies were shown to be carbohydrates. This is consistent with the results of Hamilton showing that the crossreaction is due to carbohydrate epitopes (Hamilton et al. 1999; Kantelhardt et al. 2002). The immunoblotting results suggest that some periodate-sensitive SEA components recognized by anti-KLH antibodies are the same as those recognized by anti-SEA antibodies. Further studies are needed to determine the extent to which our immunoblotting and IF results depend on the recently described major Fuc(α1→3)GalNAc motif (Kantelhardt et al. 2002).
Considering the observed intra- and perioval distribution, KLH-crossreactive antigenic material may participate in granuloma formation by serving both as immunogen and as target for an immune response.
Our results show that KLH cross-reactive immunogens are localized at the surface of cercaria, cercarial secretions, and schistosomula, the invading larval stage.
Cercarial excretion products, originating from the penetration glands, “kissing marks” (Linder 1986, 1990), are deposited at the site of attachment of cercariae in the process of scanning for a site to penetrate the skin. Those water-insoluble deposits, released on contact between the larvae and skin lipids, were shown to contain KLH-crossreactive material. The lectin reactivity of “hatch fluid” produced by intraoval miracidia has been used in a test to demonstrate excreted S. hematobium eggs (Linder 1986), and lectin-reactive material has been demonstrated periovally in granulomas of infected mice (Linder et al. 1991). “Hatch fluid” components appear to leak out through pores of the eggshells (Cao et al. 1982) into tissues, where they initiate a host immune response as part of the granulomatous reaction. The distinct staining of gonads raises questions regarding the role of carbohydrates in cell–cell interactions in germ line development. This type of gonadal staining was also seen previously with MAbs raised against GP50 (Linder et al. 1991).
Based on the observed localization of KLH-crossre-active material, the antigen appears to be both surface-associated and excreted in the larvae, the cercariae/schistosomula, and miracidia. In contrast to the surface staining seen in larval forms, no surface staining was seen in the adult worms. This is consistent with the fundamental ultrastructural difference between schistosomula and adult schistosome surface membranes (Hockley and McLaren 1973). Moreover, in adult worms the crossreacting structures seem to be related to an excretory function. These internal ducts recognized in adult worms by anti-KLH antibodies also react with PNA (Linder and Huldt 1982). This localization appears to be similar to that of anti-LDN-DF MAbs described by van Remoortere et al. (2000). The fine threadlike structures, apparently linking a subtegumental cell with the surface tubercles of the adult male worm, have previously been shown to be recognized by anti-GP50 MAb 2B11 (Linder et al. 1991), and by schistosomiasis patient sera (Linder 1994), but these structures have not been identified and their putative secretory or neural localization remains to be demonstrated.
Footnotes
Acknowledgements
Supported in part by the Swedish International Development Cooperation Agency.
We thank Aija Lassfolk for expert technical support.