
Abstract
Matriptase is a type II transmembrane serine protease that has been implicated in the progression of epithelium-derived tumors. The role of this protease in the biology of normal epithelial cells remains to be elucidated. Matriptase mRNA has been detected by Northern analysis in tissues rich in epithelial cells, and the protein is expressed in vivo in normal and cancerous breast, ovarian, and colon tissues. However, a systematic analysis of the distribution of matriptase protein and mRNA in normal human tissues rich in epithelium has not been reported. In this study we characterized the expression of the protease in a wide variety of normal human tissues using a tissue microarray and whole tissue specimens. Significant immunoreactivity and mRNA expression were detected in the epithelial components of most epithelium-containing tissues. Matriptase expression was found in all types of epithelium, including columnar, pseudostratified columnar, cuboidal, and squamous. Distinct spatial distributions of reactivity were observed in the microanatomy of certain tissues, however. This suggests that although matriptase is broadly expressed among many types of epithelial cells, its activity within a tissue may be regulated in part at the protein and mRNA levels during the differentiation of selected epithelia.
Keywords
M
Matriptase has been shown in vitro to activate the latent forms of HGF, uPA, and PAR-2, and to cleave extracellular matrix (ECM) components such as laminin and fibronectin (Lee et al. 2000; Takeuchi et al. 2000; Satomi et al. 2001). Both HGF and uPA have been proposed to regulate ECM degradation, cell proliferation, cell survival, and cell motility (Andreasen et al. 2000; Comoglio and Boccaccio 2001), all processes that are important aspects of tumor progression, including cancer invasion and metastasis. Matriptase has been detected in a variety of epithelium-derived cancer cells both in vitro and in vivo (Oberst et al. 2001; Tanimoto et al. 2001). Therefore, matriptase may ultimately regulate tumor progression by acting as an upstream activator of HGF and uPA on the surfaces of tumor cells, or by directly remodeling the ECM.
The relationship between matriptase expression in cancer cells vs their normal counterparts has not been extensively studied. However, the expression of matriptase has been preliminarily characterized in several cancer sites. In ovarian cancer, Tanimoto et al. (2001) found that TADG-15 (matriptase) was overexpressed in low malignant potential (LMP) tumors and ovarian carcinomas relative to normal ovarian surface epithelium. In a separate study by our group, matriptase expression in ovarian tumors of a surface epithelial origin was found less frequently in laterstage disease (stage III vs stages I and II). However, the expression of matriptase occurred more frequently in the absence of its inhibitor, HAI-1, in later-stage disease, suggesting that matriptase may be expressed in the absence of its endogenous inhibitor during tumor progression in a subset of tumors (Oberst et al. 2002). In node-negative breast cancer, matriptase expression is significantly associated with larger tumor size and poor disease-free survival (Kang et al. 2003). Expression also correlated with that of HGF and the HGF receptor c-met, suggesting a functional relationship between matriptase and HGF/c-met signaling. Together, these studies suggest that matriptase expression and/or activity may be altered during the conversion of normal epithelial cells to invasive carcinomas.
In normal human tissues, the expression of matriptase has been detected by Northern blotting in tissues rich in epithelium, including the prostate, stomach, small intestine, colon, lung, and kidney, as well as in placenta and peripheral blood leukocytes (Takeuchi et al. 2000). In the human breast, immunohistochemistry has revealed that expression of matriptase is higher in ductal epithelium compared to surrounding terminal duct-alveolar units (TDLUs) (Oberst et al. 2001). In the rat small intestine, a gradient of matriptase expression exists along the crypt-villous axis, with lower levels present in the crypts of Lieberkuhn and increasing levels found toward the villous tip (Satomi et al. 2001). Whether matriptase is broadly expressed by all histological types of surface lining epithelium (e.g., columnar, cuboidal, and squamous) or by a subset of epithelial types is not known, nor is it known whether matriptase expression is limited to certain organ systems.
The function of matriptase in vivo is currently unknown. The mouse orthologue of matriptase, termed epithin, was cloned from mouse thymic epithelial cells, and may play a role in epithelial cell-directed thymic T-cell maturation (Kim et al. 1999). Matriptase−/- (knockout) mice die shortly after birth as a result of a deficient epidermal barrier function in the skin of newborns (List et al. 2002), suggesting that matriptase may play a role in cell-to-cell adhesion, such as tight junction formation. In addition, these mice demonstrate abnormal hair follicle development, and disturbed thymic homeostasis, as indicated by increased lymphocyte apoptosis in the thymuses of newborn mice.
In this study we have examined the expression of matriptase protein and mRNA in a broad range of normal human tissues using a tissue microarray and selected additional whole tissue sections. We wished to determine whether matriptase is expressed broadly among epithelial tissues or is more limited to one histological type of epithelium vs another (e.g., columnar vs stratified squamous). We also wanted to determine whether the expression level was spatially graded in the microanatomy of epithelium-containing tissues, as has been described for matriptase in the human breast and for rat matriptase along the crypt-villous axis in the small intestine. Finally, we sought to determine whether matriptase expression in normal human tissues correlates with the expression pattern of HAI-1, the matriptase inhibitor, which would further support a functional link between matriptase and HAI-1 in vivo. A description of the spatial distribution of matriptase expression in normal tissues could provide a framework for understanding the function of matriptase in normal cell biology and tissue homeostasis. Such information could furthermore be useful in understanding how matriptase expression may be deregulated in human pathology, such as carcinogenesis.
Materials and Methods
Tissue Specimens
Normal human tissues were obtained as a normal human tissue microarray from Imgenex (San Diego, CA) and as whole tissue sections from the Histopathology and Tissue Shared Resource at the Lombardi Cancer Center, Georgetown University. The Imgenex tissue microarray contains 59 cores of normal human tissues representing all of the major human organ systems. Each section is an approximately 1-mm-diameter disk of paraffin-embedded, formalin-fixed tissue. All specimens on the Imgenex slide and the slides obtained from the Histopathology and Tissue Shared Resource were processed within 4 hr of removal, fixed with 10% neutral formalin, paraffin-embedded according to standard protocols, cut to 4–5-μm thickness, and placed on glass slides for immunohistochemistry (IHC) or in situ hybridization (ISH).
Monoclonal Antibody
The specificity of the anti-matriptase monoclonal antibody (MAb) S5 used for IHC has been described elsewhere (Oberst et al. 2001,2002), and was further confirmed in this study by matriptase-specific ISH.
Immunohistochemistry
IHC was performed using the Vectastain ABC kit (Vector Laboratories; Burlingame, CA) with minor modifications to the manufacturer's protocol. Sections were stained using the matriptase-specific MAb clone S5 (IgG1) (Oberst et al. 2001,2002) at a concentration of 1 μg/ml. Mouse IgG1 at a concentration of 1 μg/ml was used with a duplicate of these same sections as a negative control. The colorimetric reactions for the negative control slides were developed for the same amount of time as experimental slides, and did not show any development of the color reagent. All experimental slides were scored for matriptase (S5) immunostaining by a histopathologist (BS). Strong staining was given a score of 3 (+ + +), moderate staining a score of 2 (+ +), and weak staining a score of 1 (+). Tissues lacking staining entirely were scored 0 (−). A score of 2 or 3 (+ + or + + +) was considered positive, and a score of 0 or 1 (- or +) was considered negative for discussion purposes. To confirm the IHC staining of the tissue microarray, matriptase-specific ISH of the microarray was performed. In addition, the majority of the tissue sections from the micro-array that are represented in this study were confirmed by IHC of whole tissue sections obtained from the Histopathology and Tissue Shared Resource. The tissues from the Histopathology and Tissue Shared Resource that were stained included skin, breast and subcutis fat, skeletal muscle, lung, salivary gland, liver, pancreas, esophagus, stomach, small intestine, colon, kidney, testis, endometrium, ovary, adrenal gland, and thyroid. A concordance in IHC staining patterns was always found. Images for the figures in this study were captured from the Imgenex tissue microarray using an Olympus AH2 Vanox Microscope System (Olympus; Melville, NY).
In Situ Hybridization
Probes for ISH were prepared by generating matriptasespecific digoxigenin-labeled sense and antisense RNA riboprobes using the Dig-RNA labeling kit (Boehringer-Mannheim; Mannheim, Germany) according to a modified manufacturer's protocol as previously described (Oberst et al. 2001). Sense and antisense probes were hybridized and washed under identical conditions, and the colorimetric reactions were stopped at the same time for sense and antisense hybridized sections. The matriptase probe was designed specifically against the 5' end of the protease cDNA, which contains no sequence homology with known proteins.
Results
Tissue and Cell Distribution of Matriptase
In all of the organs examined, matriptase was predominately expressed in epithelial cells of the surface-lining epithelium. Immunoreactivity was absent from most non-epithelial cell types, including stromal fibroblasts, although expression is observed in endothelial cells and isolated tissue leukocytes (data not shown). The pattern of matriptase expression is discussed below in the context of organ systems. Representative immunohistochemical and in situ staining is shown in Figures 1–6. Results of immunohistochemical staining are summarized in Table 1.
Respiratory System. Strong matriptase expression was found in the respiratory mucosa in the nasal and bronchial lining (Figure 1A and 1B). The glandular epithelium of serous and mucous glands in the nasal mucosa also stained positively (Figure 1A). In the lung, negative staining was observed in the alveolar epithelium (Figure 1C). Staining was also seen in bronchial seromucous glandular epithelium, but not in chondrocytes within cartilage (not shown) nor in stromal fibroblasts.
Immunohistochemical reactivity of matriptase in normal human tissues
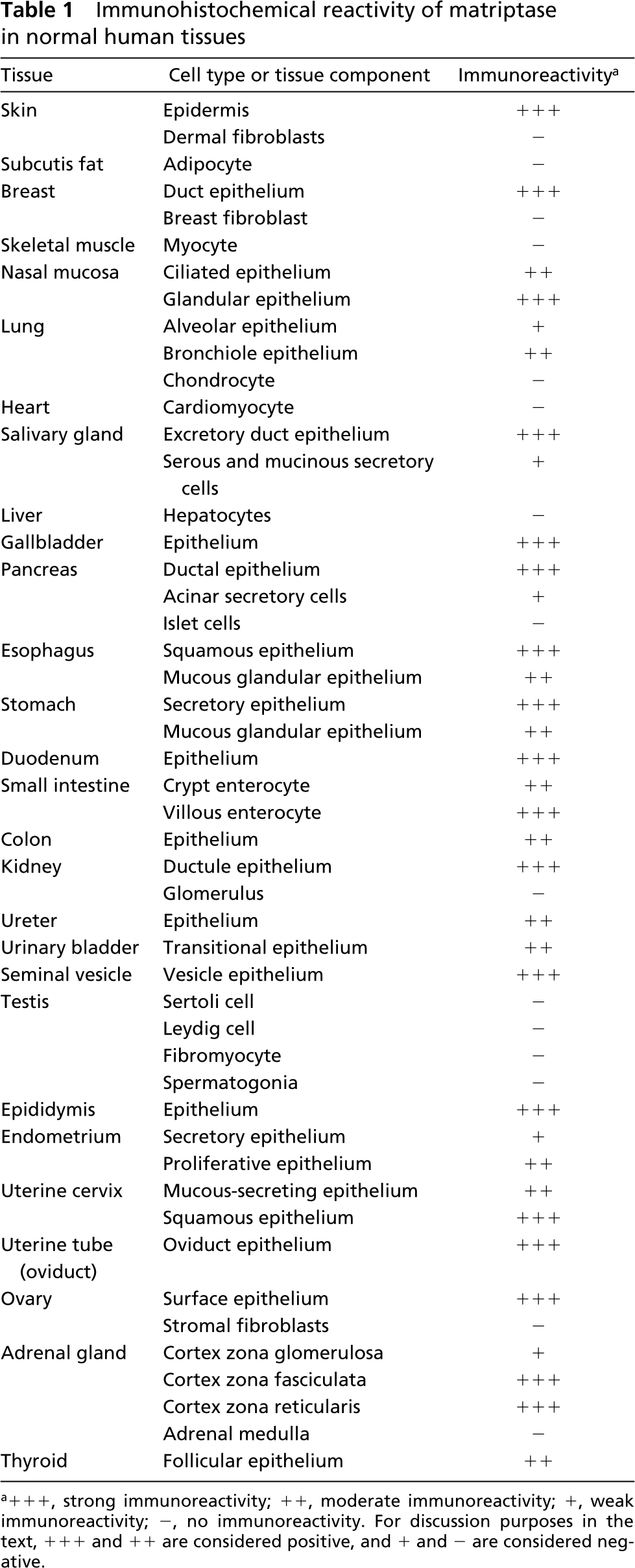
+ + +, strong immunoreactivity; + +, moderate immunoreactivity; +, weak immunoreactivity; −, no immunoreactivity. For discussion purposes in the text, + + + and + + are considered positive, and + and − are considered negative.
Digestive and Hepatobilliary System. The digestive tract epithelium showed strong matriptase immunoreactivity. In the salivary gland, strong staining was observed in excretory duct epithelium, but staining was negative in serous and mucous secretory cells (Figure 1D). Both squamous epithelium and underlying submucous glands were positive in the esophagus (Figure 2A). The basal and intermediate layers of the squamous esophageal strata showed stronger reactivity than luminal layers. ISH confirmed the production of matriptase mRNA primarily in the intermediate esophageal stratum (Figure 2B and 2C). In the stomach, the mucous neck glands and deeper gastric glands stained strongly, and moderate staining was observed in mucinous glandular epithelia (Figure 2D); ISH revealed strong staining in the deeper gastric glands (Figure 2E and 2F). In the duodenum and small intestine, a gradient of matriptase immunoreactivity was observed along the crypt-villous axis (Figures 1E-1G). Relatively moderate reactivity was seen in crypt cells, and progressively stronger reactivity appeared towards the villous tip. This observation is similar to that reported for the rat homologue of matriptase in the small intestine (Satomi et al. 2001). As in other tissues, scattered macrophages in the submucosa were strongly positive (not shown). Hepatocytes were negative (not shown). Strongly positive staining was found, however, in the gallbladder mucosa (Figure 1H). Likewise, strong matriptase staining was observed in the pancreas in ductal epithelia, with weak staining seen in acinar secretory cells (Figure 2G). ISH, however, revealed strong reactivity in acinar secretory cells (Figure 2H and 2I), indicating that matriptase may be produced by secretory units but concentrated on the surface of pancreatic ductal epithelia. Cells of the islets of Langerhans were negative (Figures 2G-2I).
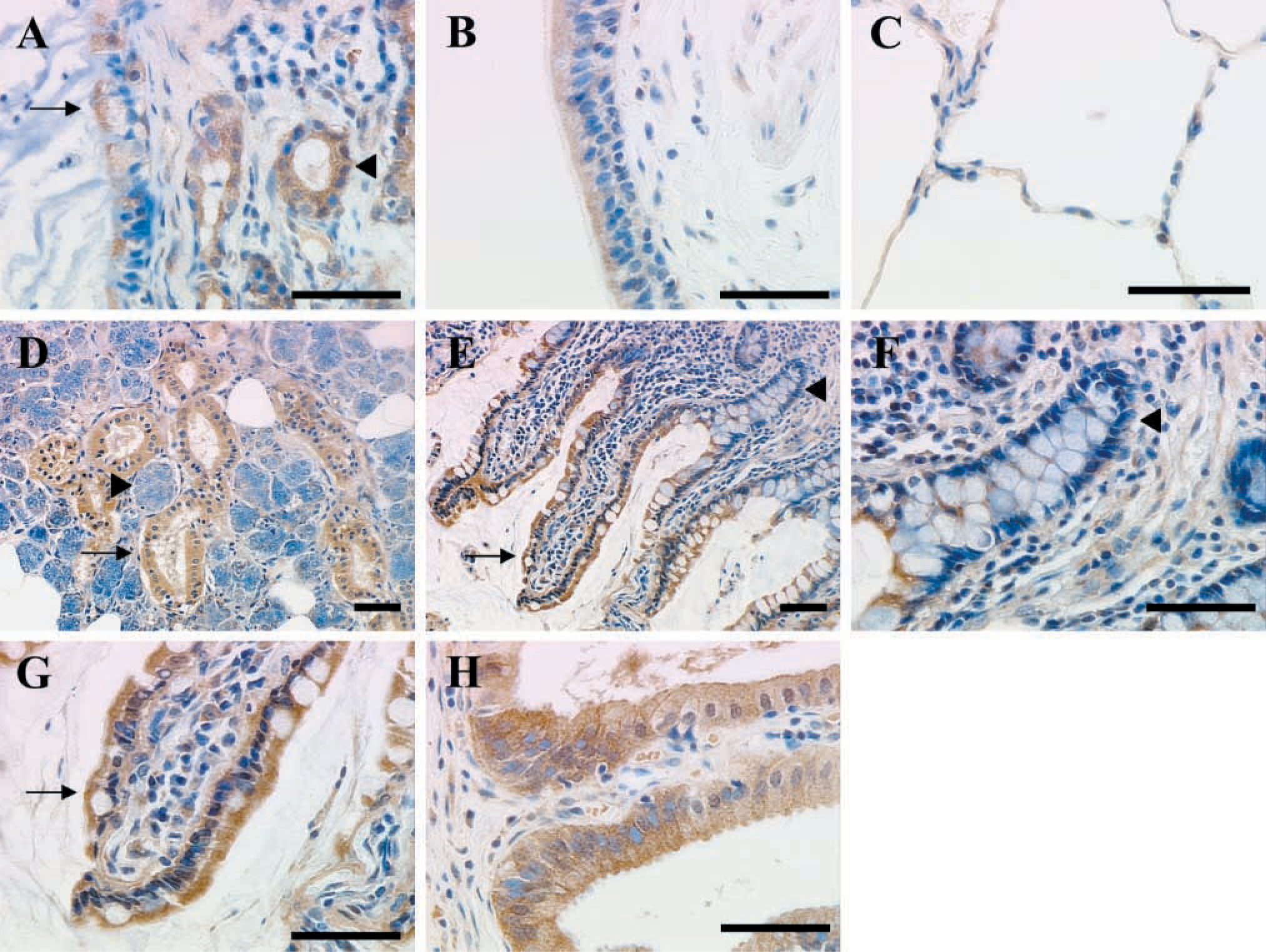
Examples of matriptase immunostaining in respiratory, digestive, and hepatobiliary tissues using MAb S5. (
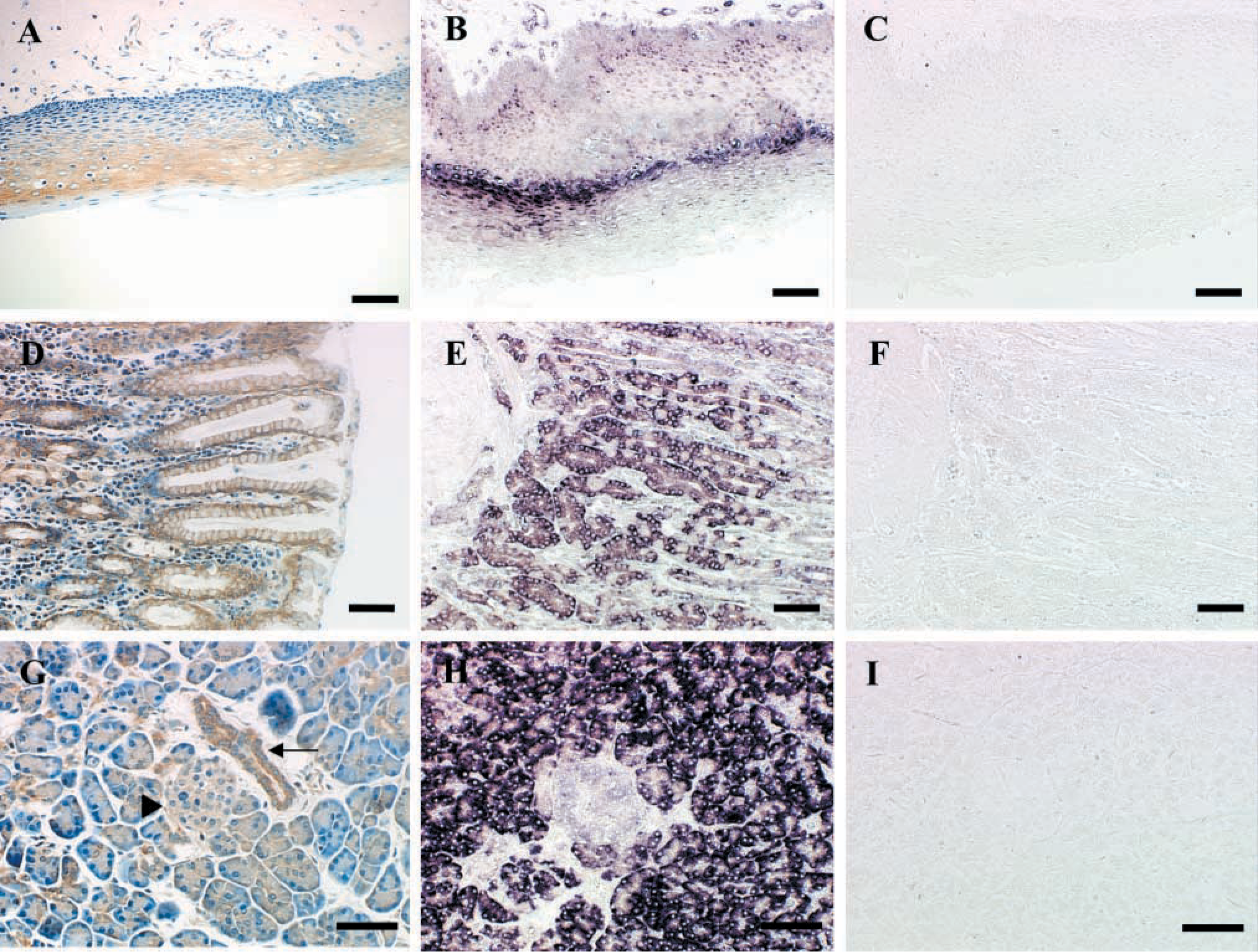
Examples of matriptase immunostaining in esophagus, stomach, and pancreas, with confirmation using matriptase-specific ISH. (
Genitourinary and Reproductive System. Kidney tubule epithelial cells showed strong reactivity, whereas cells of the glomerular apparatus were negative (Figure 3A and 3B). Transitional epithelium of the urinary bladder was positive (Figure 3C), as was the epithelium of the ureter (not shown). Strong reactivity was observed in seminal vesicle epithelium (Figure 4A) and was confirmed by ISH (Figure 4B and 4C). All cell types in the testis were negative, including Sertoli and Leydig cells, fibromyocytes, and spermatogonia (Figure 3D). The epithelial lining of the epididymis, however, was positive (Figure 4D), and was confirmed as such by ISH (Figure 4E and 4F). In the endometrium, a difference was observed in matriptase immunoreactivity in epithelium during the proliferative and secretory stages of the menstrual cycle (Figure 3E and 4F). The endometrial lining showed moderate positivity during the proliferative phase, but weak staining during the secretory phase. The stromal elements showed no reactivity. Moderate reactivity was seen in the mucous-secreting glandular epithelium of the cervix (Figure 3G). As previously reported (Oberst et al. 2002), the surface epithelial components of the ovary showed moderate reactivity, whereas the stromal components were negative (data not shown).
Endocrine System. The adrenal gland displayed a distinct pattern of matriptase staining (Figures 5A-5C). All secretory epithelial cells of the zona fasciculata and zona reticularis of the adrenal cortex stained strongly. However, cells of the zona granulosa showed mostly weak or negatively stained cells, with moderate staining appearing in some cells. The adrenal medulla, a neural crest-derived tissue, lacked reactivity (Figure 5C). As previously described, cells of the islets of Langerhans of the endocrine pancreas were negative. Thyroid follicular epithelium, however, showed matriptase staining (not shown).
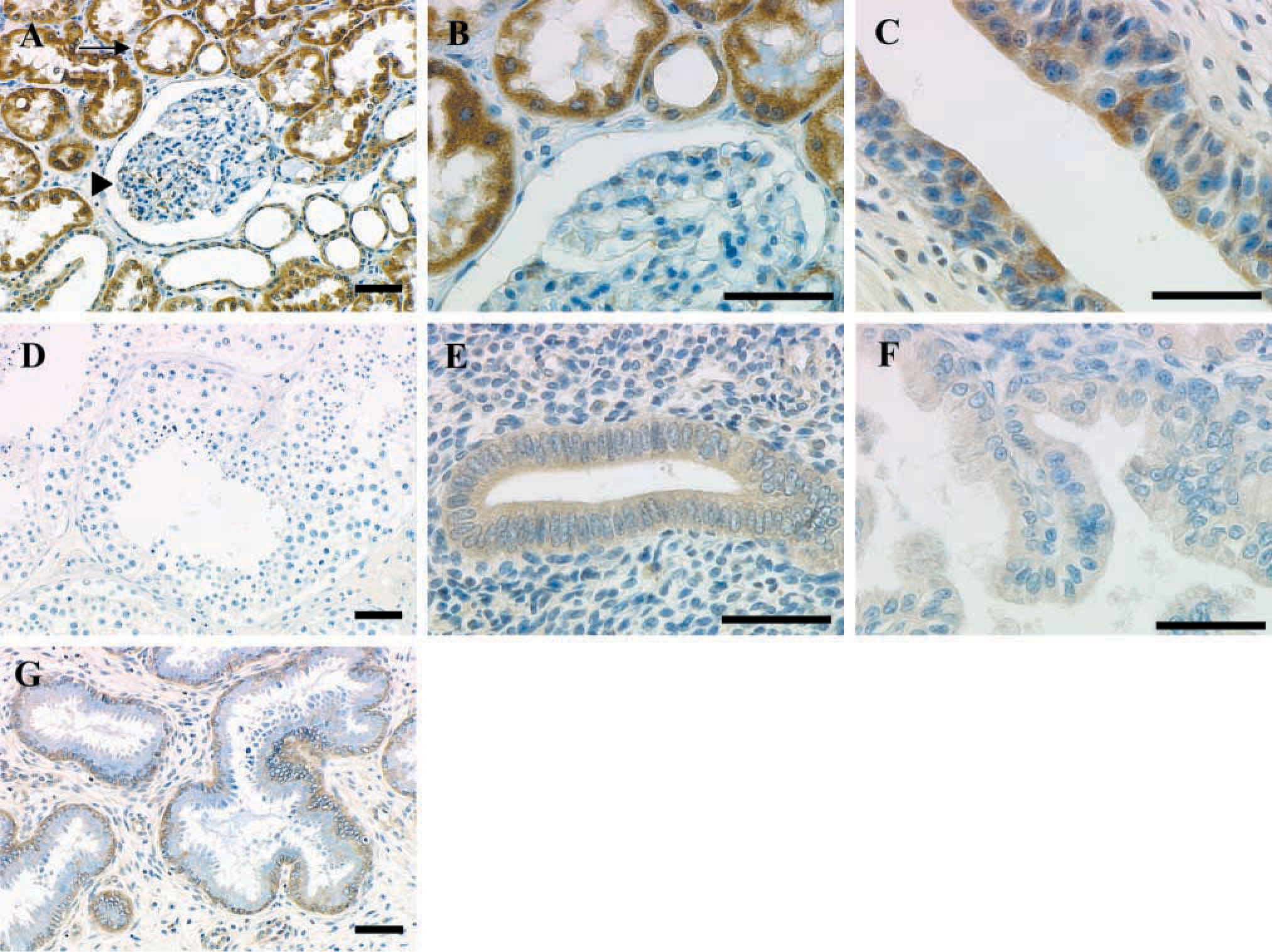
Examples of matriptase immunostaining in genitourinary and reproductive epithelia. (
Skin and Subcutaneous Tissues. Matriptase expression was detected in squamous epithelial cells in the epidermal layer of the skin (Figure 6A). No staining was seen in dermal fibroblasts. Matriptase immunostaining in the skin was confirmed by ISH (Figure 6B and 6C). As previously reported, breast duct epithelia demonstrated matriptase immunostaining, in contrast to underlying tissue stromal fibroblasts and adipocytes (Oberst et al. 2001).
Discussion
Using a normal human tissue microarray and selected normal human tissues, we examined the expression of matriptase in a broad range of epithelium-containing tissues. IHC and ISH analysis indicated that matriptase is broadly expressed in the surface epithelial lining of almost all tissues examined and was absent from most mesenchymal components. Immunoreactivity was detected in epithelia of many types: simple columnar, pseudo-stratified columnar, cuboidal, and stratified squamous. Therefore, matriptase expression is not limited to epithelia of any particular histological type, indicating that matriptase plays a role common to the function of all epithelia. However, the intensity of matriptase immunoreactivity did show distinct spatial distributions in the epithelial components of several tissues. For example, the intensity in the esophagus was strongest in the intermediate layer and less intense in the surface layer. In the intestine, a gradient of intensity was observed in the crypt-villous axis, where matriptase immunoreactivity increased towards the villous tip, consistent with the pattern previously seen for rat matriptase in the rat small intestine (Satomi et al. 2001). In the pancreas and salivary gland, matriptase protein immunoreactivity was stronger in ductal epithelia than in acinar secretory and serous/mucous glandular epithelia. This was similar to the stronger staining observed previously in breast ductal epithelium than in breast acinar epithelium (Oberst et al. 2001). A gradient was also seen, focally, in the adrenal gland. Here, immunostaining was strongest in the zona reticularis and zona fasiculata and weaker in the zona glomerulosa. Matriptase staining was stronger in proliferative endometrium than in secretory endometrium. In total, the distinct spatial gradients of matriptase immunostaining indicate that expression of the protease is differentially regulated, perhaps reflecting unique functions during the differentiation of epithelial cells within an organ system.
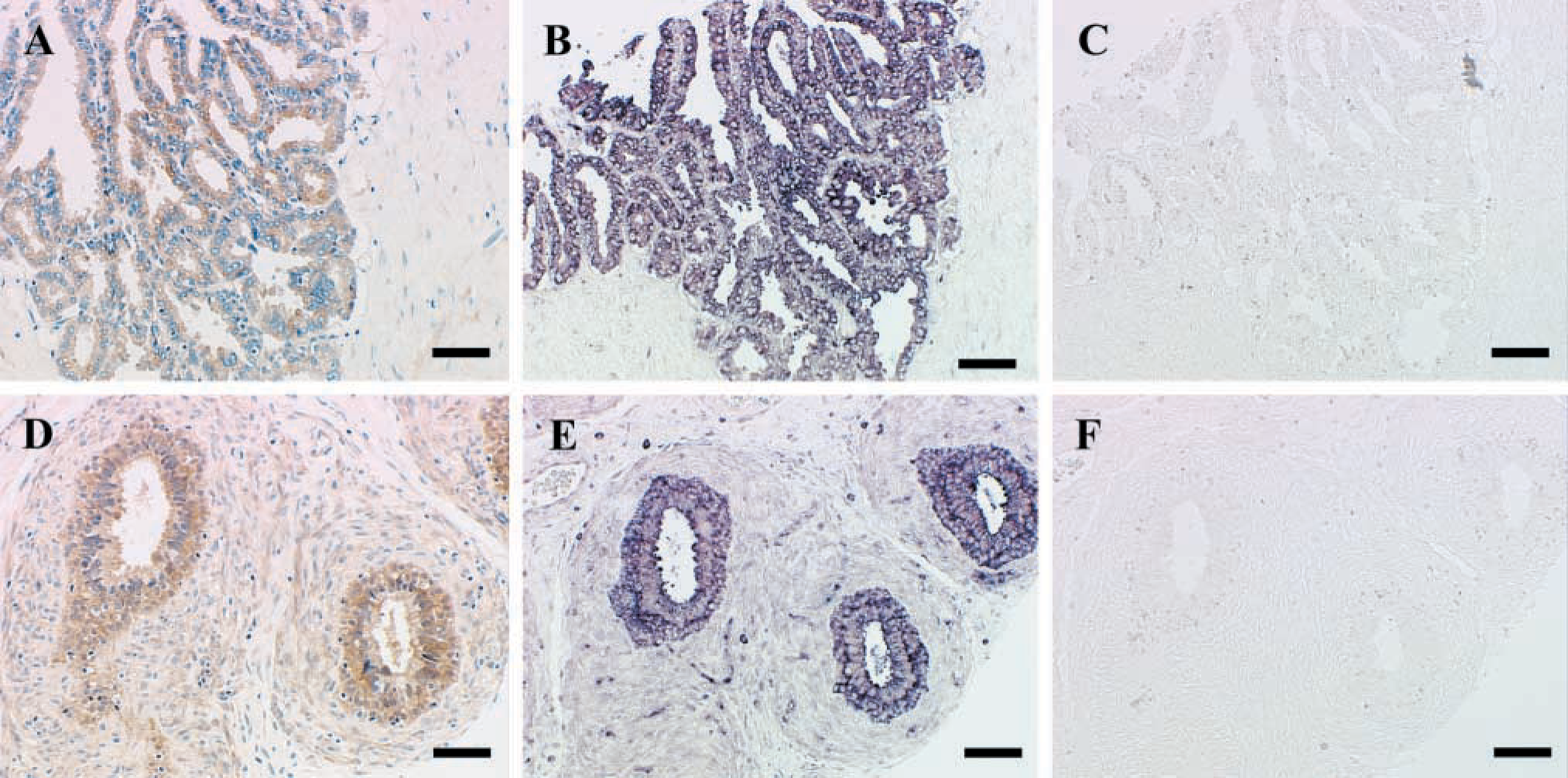
Examples of matriptase immunostaining in seminal vesicles and epididymis, with confirmation using matriptase-specific ISH. (
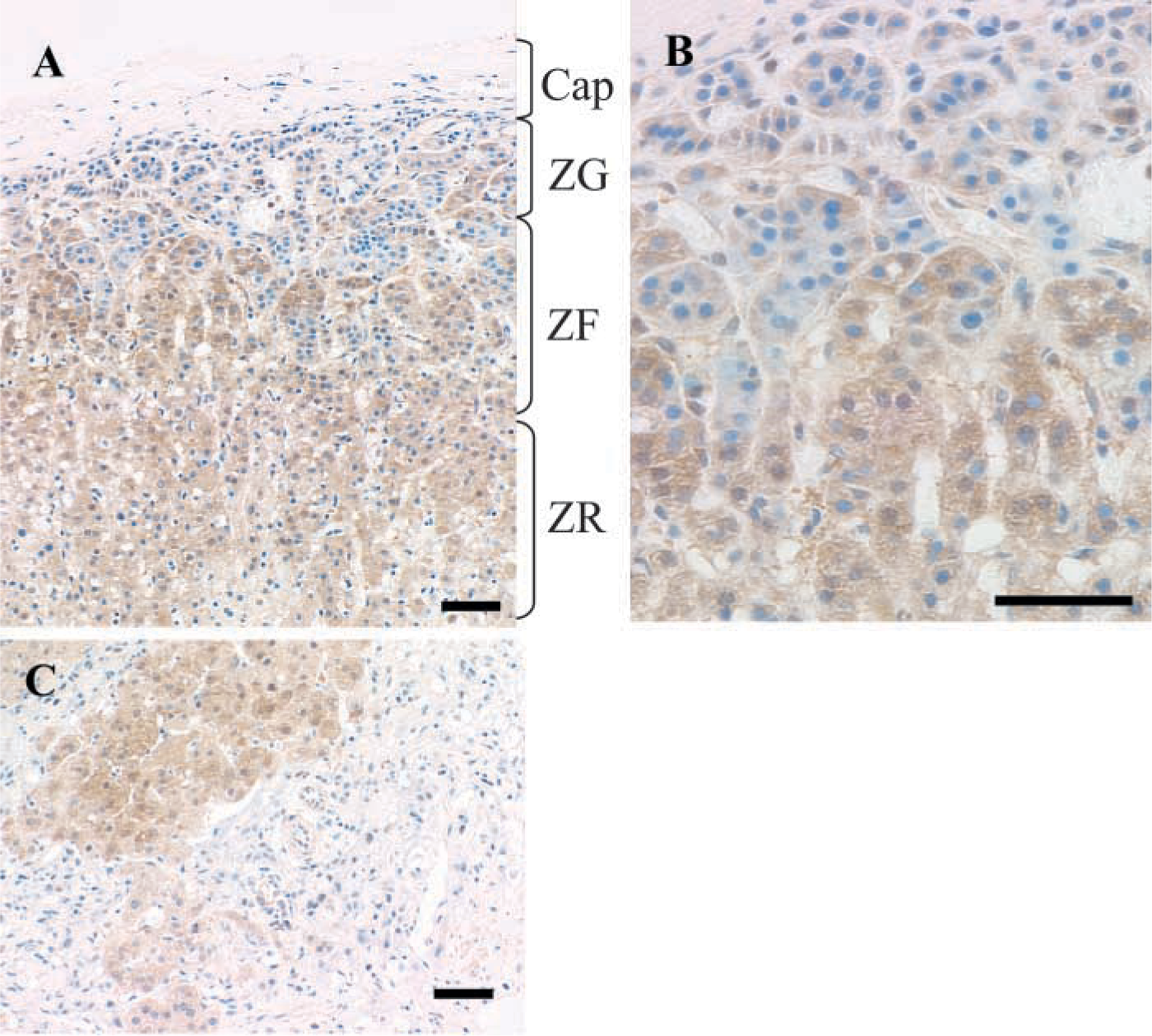
Example of matriptase immunostaining in the adrenal gland. (
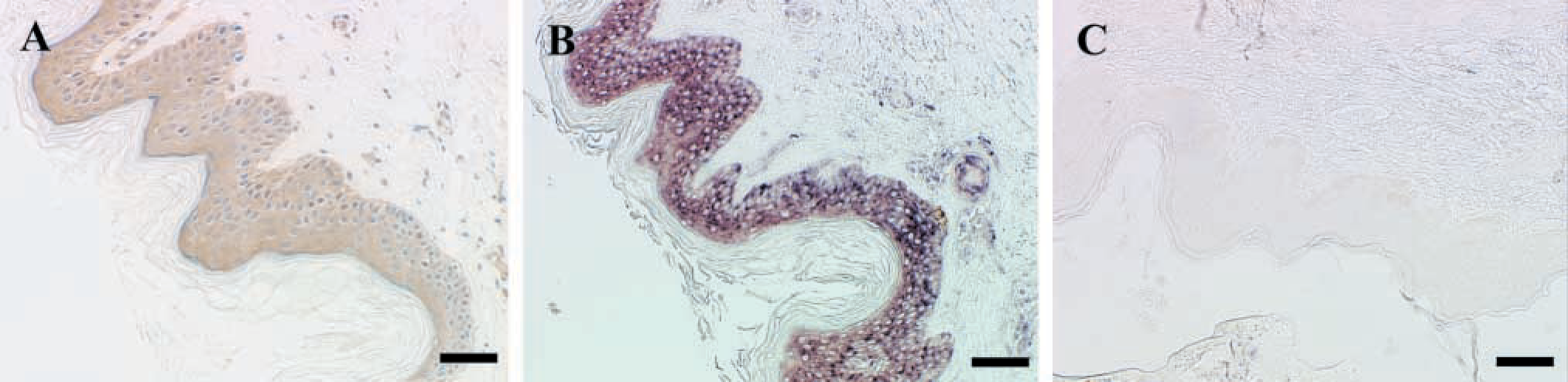
Example of matriptase immunostaining in the skin, confirmed with matriptase-specific ISH. (
The expression pattern of matriptase closely correlated with that previously described for the matriptase inhibitor HAI-1, which is broadly localized to surface lining epithelia (Kataoka et al. 1999). These results suggest that HAI-1 is spatially available to be the cognate inhibitor of matriptase in vivo, and that matriptase and HAI-1 form a functional pair in vivo as they do in vitro. It is possible that other inhibitors may also function to inhibit matripase activity in vivo. However, this study reinforces the importance of taking into consideration the expression of HAI-1 when considering matriptase expression and any presumed link to activity. For example, loss of HAI-1 expression in tumors could signify unopposed matriptase activity. Alternatively, an increased expression of HAI-1 in tumors could indicate a compensatory response to increased matriptase activity.
To date, several in vitro substrates for matriptase have been identified and suggest a role for the protease in matrix remodeling and the regulation of cell growth and survival, cell motility, and cell morphogenesis. However, without proven knowledge of the true in vivo substrates of the enzyme, it is difficult to predict the physiological role of matriptase in normal human tissues. Therefore, an important future goal must be the demonstration of the in vivo cleavage and activation of putative matriptase substrates. In addition, it will be essential to characterize how matriptase protease activity is regulated physiologically. This information, together with the above description of the relevant cell types that express matriptase in vivo, will help pave the way for understanding the role of matriptase in the biology of normal epithelia, and how deregulation of its activity may promote pathological states in multiple organ systems.
Footnotes
Acknowledgements
Supported by the Susan G. Komen Breast Cancer Foundation DRA99-003037 and BCTR0100345 and DOD grants DAMD 17-01-1-0252, DAMD 17-02-1-0391, and DAMD 17-00-1-0264.
We acknowledge the Histopathology and Tissue Shared Resource of the Lombardi Cancer Center at Georgetown University Medical Center for help in selecting tissue specimens. We would also like to acknowledge the assistance of the Tissue Culture Shared Resource and the Microscopy and Imaging Resource. These resources are supported by NIH grants P50-CA58185 (TCSR) and P30-CA51008 (TCSR and MIR).