
Abstract
Osteogenic cells express some matrix proteins at early culture intervals. The aim of this study was to determine if, and in what proportion, cells used for plating contain bone sialoprotein (BSP) and osteopontin (OPN), two matrix proteins associated with initial events in bone formation. Their pattern of expression, as well as that of fibronectin (FN) and type I pro-collagen, was also examined at 6 hr and at 1 and 3 days. The cells were obtained by enzymatic digestion of newborn rat calvariae, and grown on glass coverslips. Cytocentrifuge preparations of isolated cells and coverslips were processed for single or dual immunolabeling with monoclonal and/or polyclonal primary antibodies, followed by fluorochrome-conjugated antibodies. The cell labeling was mainly associated with perinuclear elements. OPN was also distinctively found at peripheral cytoplasmic sites. About 31% of isolated cells were OPN-positive and 18% were BSP-positive. After 1 day, almost 50% of cells were immunoreactive for OPN and for type I pro-collagen, and still less than 20% reacted for BSP. Approximately 7% exhibited peripheral staining for OPN. Almost all cells were associated with extracellular FN. However, only 15% showed intracellular labeling. These results indicate that an important proportion of cells used for plating contain BSP and OPN, a situation that should be taken into consideration in experimental analyses of osteoblast activity in vitro.
Keywords
O
The expression pattern of matrix proteins in vitro has been correlated with acquisition and maturation of the osteoblast phenotype (Owen et al. 1990; Stein and Lian 1993; Lynch et al. 1995; Moursi et al. 1996). For some of these proteins, expression levels fluctuate over time (Owen et al. 1990; Stein and Lian 1993; Yao et al. 1994; Aarden et al. 1996; Nefussi et al. 1997; Zohar et al. 1998; Malaval et al. 1999; Luan et al. 2000). Osteopontin (OPN) is first expressed during the period of active cell proliferation, decreases post-proliferatively, and then reincreases at the onset of mineralization, to achieve peak levels during mineralization (Owen et al. 1990; Stein and Lian 1993; Yao et al. 1994; Zohar et al. 1998). A transient expression of BSP in differentiating osteoblasts has also been proposed (Malaval et al. 1999). However, its production is generally believed to initiate in newly differentiated osteoblasts (Ganss et al. 1999). Therefore, although some of the observed early expression of OPN and BSP is associated with the osteoblast differentiation sequence in vitro (Owen et al. 1990; Stein and Lian 1993; Malaval et al. 1999), the possibility that it may derive, at least in part, from synthesis before cell isolation has also been raised (Owen et al. 1990).
In culture, ideally, one would want to have a single cell population that evolves synchronously over time. However, cell isolates used for osteogenic cultures usually comprise a mixture of populations, ranging from precursor cells to fully differentiated osteoblasts (Bellows et al. 1986; Bellows and Aubin 1989; Zohar et al. 1997a,b). Although this situation is recognized, the proportion of differentiated cells manufacturing BSP and OPN, two multifunctional proteins expressed early during bone formation, has to our knowledge not been established with precision. The aim of the present study was to determine if, and in what proportion, rat calvarial osteogenic cells isolated enzymatically contain BSP and OPN. In addition, we have examined the cytoplasmic and extracellular distributions of these two proteins, as well as those of fibronectin (FN) and type I pro-collagen, at early time points.
Immunolabeling was selected as the protein detection method because molecular signals do not always correlate with the timing of protein synthesis and secretion and do not provide any indication of localization. The results obtained indicate that a significant proportion of differentiated osteoblasts in cell isolates already produce matrix proteins and, most likely, secrete them as soon as they are placed in culture.
Materials and Methods
Cell Isolation and Primary Culture of Osteogenic Cells
Cells were isolated by sequential trypsin/collagenase digestion of calvarial bone from newborn (2–4 days) Wistar rats (Charles River Canada; St-Constant, QC, Canada) as previously described (Nanci et al. 1996; Irie et al. 1998). Osteogenic cells (viability of cells ranged between 85 and 95%) were plated on Fisherbrand 12-mm round glass cover-slips (Fisher Scientific; Nepean, ON, Canada) in 24-well plates at a cell density of 1.5 × 104 cells/well and grown for periods of 6 hr, 1 day, and 3 days in Gibco Minimum Essential Medium (MEM) with Earle's salts (Invitrogen; Burlington, ON, Canada) supplemented with 10% fetal bovine serum (Invitrogen) and 1% penicillin-streptomycin (Invitrogen) at 37C in a humidified atmosphere with 5% CO2. All animal procedures were in accordance with guidelines of the Comité de Déontologie de l'Expérimentation sur les Animaux of Université de Montréal.
Cytocentrifuge Preparations of Isolated Cells
For preparation of cytocentrifuge slides, 60-μl aliquots of isolated cells suspended in MEM (2.5 × 105 cells/ml) were placed in each Cytofunnel of a Cytospin 3 centrifuge (Thermo Shandon; Pittsburgh, PA) and spun onto polylysine-coated microscope slides (Baxter Diagnostics; Deerfield, IL) at 900 rpm for 5 min. All slides were allowed to dry for 1 hr before fixation.
Immunofluorescence
Cells grown on coverslips and cytocentrifuge preparations of cells used for plating (time 0) were fixed for 15 min at room temperature (RT) in 4% paraformaldehyde buffered with 0.1 M PBS, pH 7.2. After washing in PBS, cells were then routinely processed for immunofluorescence labeling. Briefly, they were permeabilized with 0.5% Triton X-100 in PBS for 15 min, followed by blocking with 1% ovalbumin in PBS for 30 min. For extracellular labeling, cells were not permeabilized. Primary monoclonal and/or polyclonal antibodies to BSP, OPN, FN, type I pro-collagen, and tubulin (for detecting cell outlines) were used (Table 1), followed by corresponding Alexa Fluor 488 (green fluorescence)- or 594 (red fluorescence)-conjugated goat secondary antibodies (Molecular Probes; Eugene, OR), or by rhodamine (TRITC)-conjugated secondary antibodies (Jackson Immunoresearch Laboratories; West Grove, PA), diluted 1:200 in PBS. All incubations were performed in a humidified environment for 60 min at RT, followed by three 5-min washes in PBS. Replacement of the primary antibody with PBS was used as control. For type I pro-collagen labeling, incubations with pre-immune IgG were also carried out. Dual labeling was done with a 1:1 mixture of two primary antibodies followed by a 1:1 mixture of corresponding secondary antibodies. Mixtures were prepared to yield the same working dilutions as the ones used in single labeling (Table 1).
Coverslips were mounted on microscope slides with Air-vol 205 (Airproducts; Allentown, PA) and examined by epi-luminescence under a conventional fluorescence microscope (Axiophot; Carl Zeiss, Oberkochen, Germany), using Plan-Neofluar objectives (X40, NA 0.75, and X100, NA 1.30). For quantitative analysis, approximately 20 microscopic fields at X100 were randomly selected for a total of 350–450 cells. Proportions of immunoreactive cells were expressed as the mean percentage per microscopic field ± standard deviation and comparisons were carried out using the non-parametric Mann-Whitney U-test for independent samples. Photographic recordings were made on Kodak Elite Chrome 400 reversal film (Eastman Kodak; Rochester, NY) with different exposure times (5-15 sec), depending on the intensity of fluorescence labeling. Double photographic exposure was done for the simultaneous visualization of dual labeling. Slides were then digitally recorded at 1200 dpi using an Epson Expression 1600 scanner and processed with Adobe Photoshop software. The results described below are representative of at least three different sets of primary cultures, except for cytocentrifuge preparations, which were prepared from two different digests only.
Primary antibodies used and their working dilutions
Results
Cell Morphology
Phase-contrast visualization of enzymatically released calvarial cell preparations (time 0) fixed after cytocentrifugation revealed partially spread, rounded cells with variable dimensions, nucleus-to-cytoplasm ratios, and vacuolation (Figure 1). Fixation of cell suspensions before cytocentrifugation resulted in refractile, rounded cells (data not shown). Although the latter preparation permitted immunolabeling of cells with different antibodies, it was not used in this study because of the difficulty in assessing nuclear and cytoplasmic morphology by phase-contrast.
Calvarial cells in primary cultures exhibited variable phenotypes and dimensions (Figures 2 and 3), and, as expected, became progressively larger and more spread over the 3-day culture period. By 6 hr, cells attached and partially spread on the coverslip surface, showing predominantly a polygonal morphology with relatively short cytoplasmic extensions (Figure 2). By days 1 and 3, morphology was variable and cells were polygonal, stellate, or fusiform (Figure 3). Occasionally some of them had long cytoplasmic extensions, which frequently contacted other cells. Mitotic figures were also found at all time points.
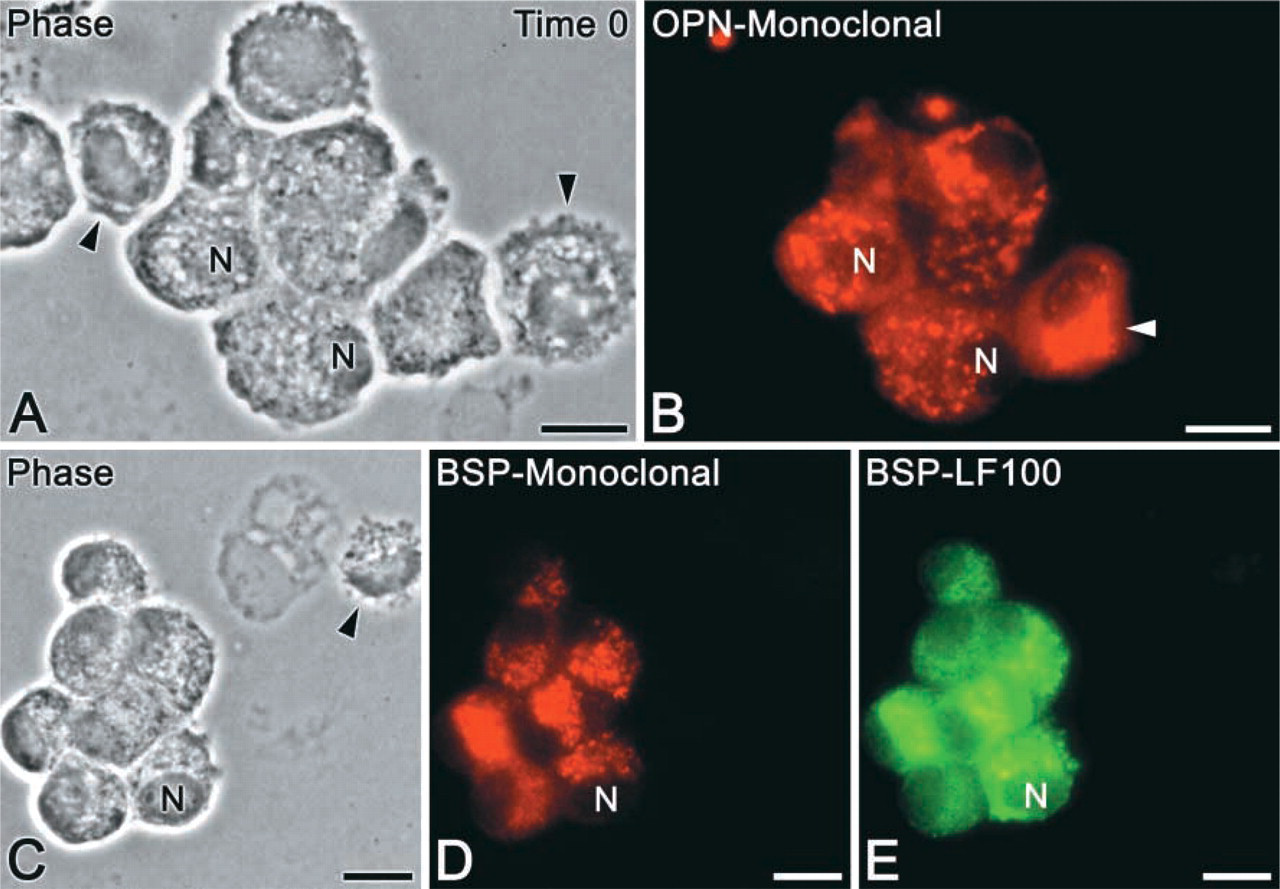
Cytocentrifuge preparations of isolated calvarial cells (time 0). (
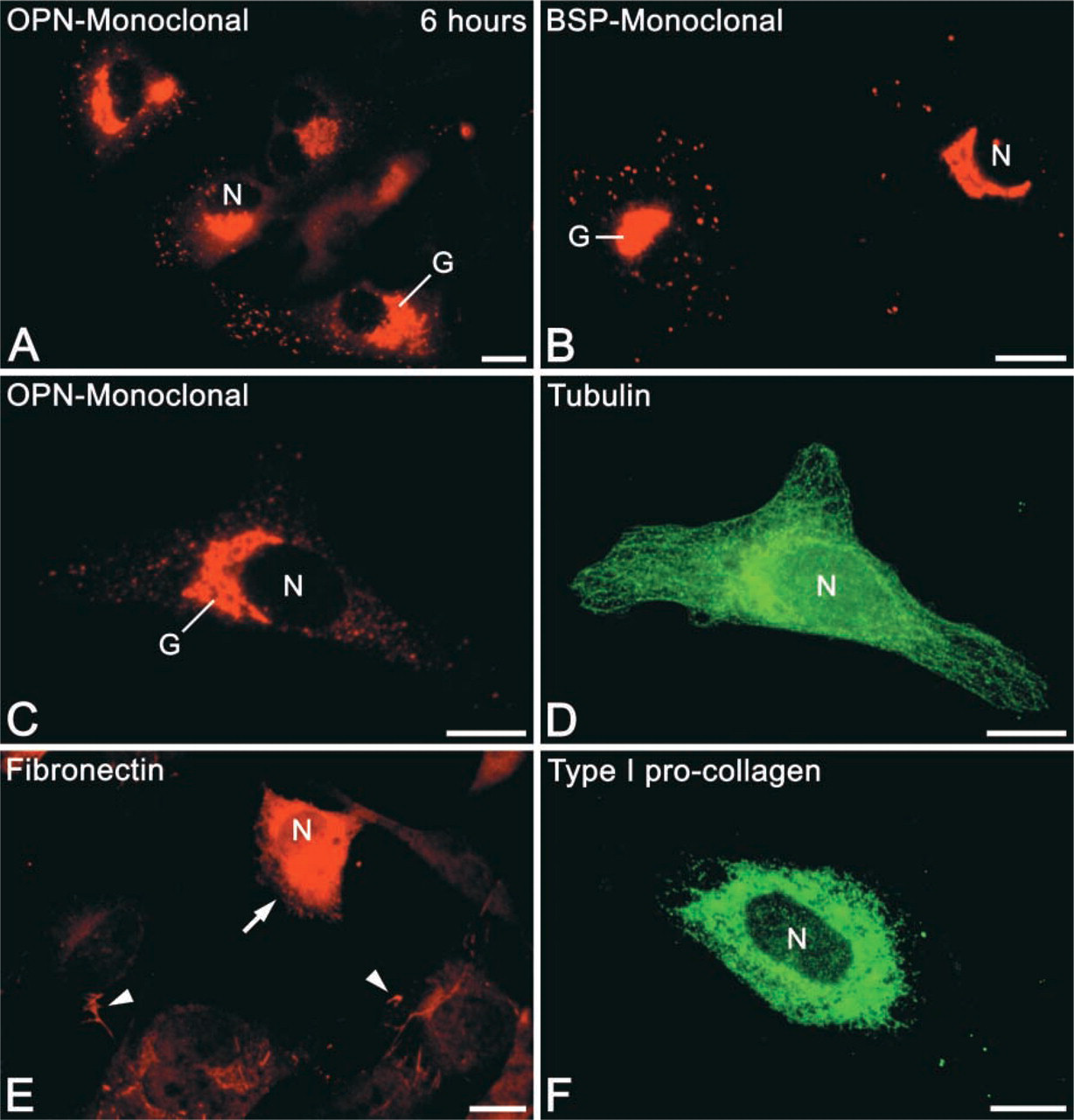
Immunofluorescence preparations of osteogenic cell cultures at 6 hr. (
Antibodies
The various primary antibodies used labeled different proportions of cells and yielded different fluorescence intensities. Dual labeling with monoclonal and polyclonal antibodies against BSP or OPN showed similar labeling patterns, as confirmed by the presence of yellow fluorescence with double photographic exposure. However, immunoreactivity appeared more defined with monoclonal antibodies (compare Figures 1D and 1E). Polyclonal antibodies against chicken tubulin and against recombinant N-propeptide of the α1 (I) human collagen chain recognized epitopes in rat osteogenic cells and resulted in characteristic labeling patterns. No significant labeling was observed in control incubations.
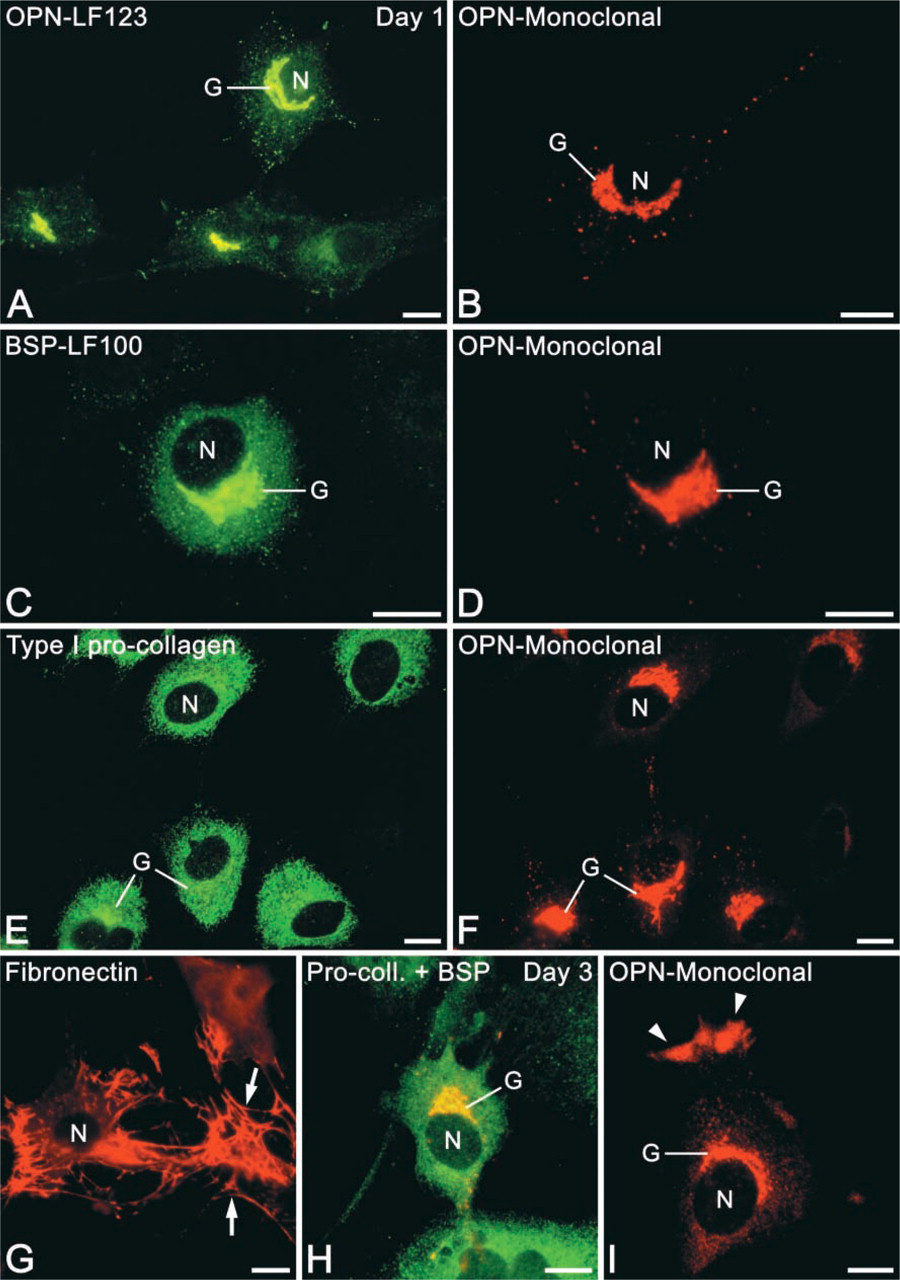
Immunofluorescence labeling of osteogenic cells after 1 day (
Distribution of Labeling
In a majority of cases, OPN was localized in an extensive perinuclear tubular network, vesicular structures, and punctate deposits throughout the cytoplasm (Figures 1B, 2A, 2C, and 3A, 3B, 3D, 3F, and 3I). Some cells showed labeling of a large, juxtanuclear region reminiscent of the Golgi area in active osteoblasts (Figures 2A, 3D, and 3F). A small proportion of OPN-positive cells (see below) also contained a peripheral cytoplasmic labeling (so-called “perimembranous in-tracellular OPN”; Zohar et al. 1997a) that appeared as focal subplasmalemmal accumulations of immunofluorescence with a dense, finely granular texture (Figure 3I, arrowheads). The frequency and intensity of labeling of these accumulations increased over the 3-day culture period and was often observed together with juxtanuclear labeling (Figure 3I). There was no discernible extracellular accumulation of OPN-reactive material at any of the time points.
Anti-BSP fluorescence was distinctively and mainly detected in the juxtanuclear Golgi area and as punctate deposits throughout the cytoplasm (Figures 1D, 1E, 2B, 3C, and 3H). Some rare cells showed perinuclear labeling. Extracellular accumulations of BSP-reactive matrix were sporadically detected at day 3, mainly in areas with relatively higher cell density.
At all time points, type I pro-collagen labeling appeared diffuse around the nucleus (Figures 2F, 3E, and 3H). Near the periphery of the cells, the labeling assumed an almost reticular pattern and, at day 3, fluorescent globular structures were sometimes found in this region. In addition, at 6 hr, a subset of immunoreactive cells exhibited labeling of the Golgi area and some granules. Both labeling patterns of type I pro-collagen were not observed when the primary antibody was replaced with pre-immune IgG.
In permeabilized and non-permeabilized cells, FN labeling revealed an extracellular fibrillar network extending from one cell to another, but not exclusively associated with the cell surface. These findings occurred in areas of high cell density at all time points (Figure 3G). In areas of low cell density, only small amounts of FN could be observed and appeared as small dots or short fibrillar structures associated with the cell surface. In permeabilized cells, strong diffuse perinuclear labeling was occasionally observed (Figure 2E). Cells showing such labeling were frequently associated with trails of FN on the substrate adjacent to them.
It is generally accepted that cell fixation with para-formaldehyde does not preserve microtubules very well. Nevertheless, tubulin labeling was detected throughout the entire cytoplasm (Figure 2D), revealing cell outlines that closely matched the ones observed by phase-contrast.
Proportions of immunoreactive cells at time of isolation and after 1 day in culture a
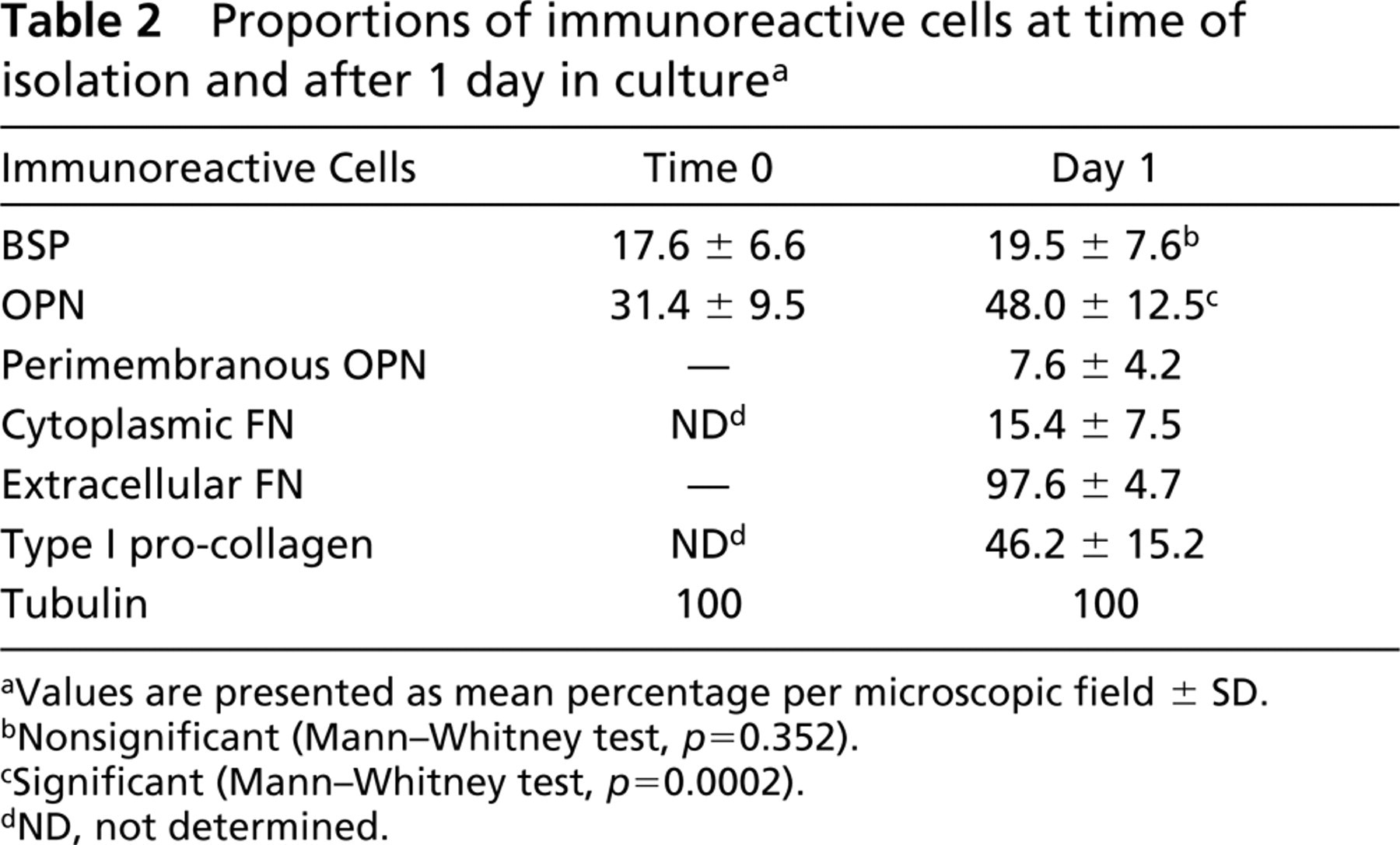
aValues are presented as mean percentage per microscopic field ± SD.
bNonsignificant (Mann-Whitney test, p = 0.352).
cSignificant (Mann-Whitney test, p = 0.0002).
dND, not determined.
Quantitative Analysis of Labeled Cells
Quantitative analysis of immunoreactive cells was carried out at two time points (time 0 and day 1). Different proportions of osteogenic cells were immunoreactive for BSP, OPN, FN, type I pro-collagen, and tubulin (Table 2). Cytocentrifuge preparations (time 0) contained about two times more OPN- than BSP-positive cells (p<0.0001). Almost all BSP-positive cells (97%) were also immunoreactive for OPN. Tubulin labeling was detected in all isolated cells.
After 1 day in culture, almost half the cell population was OPN- and type I pro-collagen-positive (p = 03192) but only 27% of cells were dual-labeled with these antibodies (Figures 3E and 3F). The proportion of cells labeled with BSP antibodies was significantly lower than that of OPN- and type I pro-collagen-positive cells (p<0.0001 for both). About 20% of cells were immunoreactive for BSP and virtually all of these cells also expressed OPN (Figures 3C and 3D) and type I pro-collagen. Cells with cytoplasmic FN labeling occurred in almost the same proportion as BSP-positive cells (p = 0.0427), whereas extracellular FN was associated with almost all cultured cells. Peripheral accumulations of OPN were found in about 7% of cells. All cells in the culture were immunoreactive for tubulin.
Discussion
In the present study, we demonstrated that a significant proportion of cells enzymatically released from newborn rat calvarial bone are immunoreactive for some major non-collagenous matrix proteins (NCPs). In addition, at early time points, different proportions of osteogenic cells express collagenous and NCPs. Therefore, the early expression of some matrix proteins by cultured osteogenic cells does not necessarily represent de novo gene expression but may also reflect pre-existent synthetic activity taking place during bone matrix formation and mineralization in vivo, before sacrifice of the animal.
Trypsin/collagenase digestion of fetal or newborn rat calvarial bone yields a heterogeneous cell population, which comprises osteoprogenitors, pre-osteoblasts, differentiated osteoblasts, and fibroblasts (Bellows et al. 1986; Bellows and Aubin 1989; Zohar et al. 1997a,b). In this context, because BSP expression begins in rat fetal tissues at day 17 (Chen et al. 1992) and has been associated with fully differentiated osteoblasts (Ganss et al. 1999), the proportion of BSP-immunoreactive cells in our isolates (time 0) must represent osteoblasts and osteocytes (Candeliere et al. 2001; Kato et al. 2001). About the same proportion of isolated cells expressed BSP after 1 day in culture and these showed a labeling pattern suggestive of active osteoblasts (Bianco et al. 1993), consistent with the above interpretation. In the case of OPN, its expression begins earlier during the osteoblast differentiation sequence (Owen et al. 1990; Stein and Lian 1993; Yao et al. 1994; Zohar et al. 1998). Therefore, OPN-positive cells in isolates may comprise pre-osteoblasts, differentiated osteoblasts, and osteocytes (Aarden et al. 1996; Kato et al. 1997; Candeliere et al. 2001). In this context, the variability in the pattern of intracellular OPN labeling around the nucleus, likely representing Golgi labeling, suggests different levels of cell differentiation, functional state, and/or the presence of different cell types. For example, cultured fibroblasts can also express OPN (Nomura et al. 1988; Zohar et al. 1997a,2000) and are difficult to distinguish morphologically from osteogenic cells at early time points (Malaval et al. 1999; Ducy et al. 2000). Dual labeling for BSP and OPN was important here because it allowed identification of the subset of active osteoblasts among OPN-positive cells. Indeed, virtually all BSP-positive cells also contained OPN and type I pro-collagen.
In vivo and in vitro studies have demonstrated that differentiated osteoblasts are phenotypically diverse. Expression of OPN, BSP, and osteocalcin by these cells is extremely variable (Liu et al. 1997; Aubin 1998; Malaval et al. 1999; Candeliere et al. 2001). It has been suggested that only type I pro-collagen, alkaline phosphatase, and pth/pthrp receptor are ubiquitous markers of the osteoblastic lineage (Malaval et al. 1999; Candeliere et al. 2001). Therefore, type I pro-collagen-positive cells that did not express BSP could not only be pre-osteoblasts and/or fibroblasts but some of them may also represent a particular state of differentiated osteoblasts.
Although OPN and BSP are generally regarded as multifunctional extracellular matrix molecules involved in cell adhesion and migration and in the regulation of mineral deposition (McKee and Nanci 1996a,b; Ganss et al. 1999; Byzova et al. 2000; Sodek et al. 2000; Giachelli and Steitz 2000; Harris et al. 2000; Denhardt et al. 2001; Fisher et al. 2001), it has recently been proposed that an intracellular form of OPN, so-called “perimembranous intracellular OPN,” is an integral component of a hyaluronan-CD44-ERM (ezrin, radixin, moesin) attachment complex (Zohar et al. 1997a,1998,2000). This complex is believed to be involved in the migration of embryonic fibroblasts, fetal calvarial cells, activated macrophages, and metastatic cells in vitro. It can therefore be assumed that the small proportion of cells exhibiting peripheral cytoplasmic labeling for OPN observed in our study likely represent migrating cells. Modulation of the proportion of osteogenic cells that exhibits such a behavior may be important in determining the rate of osteogenic events in normal and pathological conditions and may also influence bone healing events.
Even though only 15% of cells showed intracellular FN labeling, the association of almost all cells with extracellular FN at early time points is not unexpected because (a) cultured cells are capable of assembling FN fibrillar matrix in the presence of soluble FN molecules (Sottile et al. 1998; Schwarzbauer and Sechler 1999) and (b) FN is important for the initial attachment of osteogenic cells in culture (Winnard et al. 1995; Yang et al. 2002). Tubulin labeling was found throughout the cytoplasm of all cultured cells, and the cell outlines revealed by the immunofluorescence matched closely those in phase-contrast images. Immunodetection of this microtubular protein therefore represents an advantageous marker in dual labeling studies of cells cultured on non-transparent surfaces (i.e., metal and hydroxyapatite surfaces), in which cell morphology cannot be assessed by phase-contrast microscopy.
In conclusion, although it is generally agreed that enzymatically isolated cells comprise mixed populations, we have analyzed quantitatively the percentage of cells in the isolate and after short-term culture that express matrix proteins. A non-negligible proportion of cells isolated from newborn rat calvaria were found to already contain, at least, BSP and OPN. Most likely, they also express other matrix proteins such as FN and type I pro-collagen. Based on the labeling pattern in the Golgi area (Mark et al. 1987; Bianco et al. 1993), it can be concluded that the cells that immunoreacted for BSP and OPN are probably osteoblasts and/or osteocytes. Such differentiated cells may exert an influence on progenitor or differentiating cells and affect their kinetics (discussed in Malaval et al. 1999). In addition, because these two proteins are associated with initial osteogenic events in vitro (Ganss et al. 1999; Sodek et al. 2000), any pre-existent synthetic activity must be taken into consideration in experimental analyses of osteoblastic cell differentiation and bone-like matrix formation in culture.
Footnotes
Acknowledgements
Supported by the Canadian Institutes of Health Research (CIHR). Paulo Tambasco de Oliveira is the recipient of a postdoctoral fellowship (00/11604–4) from FAPESP (Brazil) and is also supported by the University of São Paulo (Brazil).
We thank Ana Carina da Paula (Department of Immunology and Microbiology, Université de Montréal) for technical assistance in cytocentrifuge preparations, Dr Larry W. Fisher (NIH) for providing anti-BSP (LF-87 and LF-100) and anti-OPN (LF-123) antibodies, Drs M.C. Farach-Carson and W.T. Butler (University of Texas at Houston, TX) for the anti-rat OPN antibody, and Drs Malcolm Collins and Paul Bornstein (University of Washington at Seattle, WA) for an antibody to recombinant N-propeptide of the α1 (I) human collagen chain. The mouse monoclonal anti-rat osteopontin (MPIIIB10–1) and anti-rat bone sialoprotein (WVID1-9C5) antibodies, developed by Michael Solursh and Ahnders Franzen, were obtained from the Developmental Studies Hybridoma Bank formed under the auspices of the NICHD and maintained by The University of Iowa, Department of Biological Sciences (Iowa City, IA).