
Abstract
Mutations in the HFE gene and a newly identified second transferrin receptor gene, TfR2, cause hemochromatosis. The cognate proteins, HFE and TfR2, are therefore of key importance in human iron homeostasis. HFE is expressed in small intestinal crypt cells where transferrin-iron entry may determine subsequent iron absorption by mature enterocytes, but the physiological function of TfR2 is unknown. Using specific peptide antisera, we examined the duodenal localization of HFE and TfR2 in humans and mice, with and without HFE deficiency, by confocal microscopy. We also investigated potential interactions of these proteins in human intestinal cells in situ. Duodenal expression of HFE and TfR2 (but not TfR1) in wild-type mice and humans was restricted to crypt cells, in which they co-localized. HFE deficiency disrupted this interaction, altering the cellular distribution of TfR2 in human crypts. In human Caco-2 cells, HFE and TfR2 co-localized to a distinct CD63-negative vesicular compartment showing marked signal enhancement on exposure to iron-saturated transferrin ligand, indicating that HFE preferentially interacts with TfR2 in a specialized early endosomal transport pathway for transferrin-iron. This interaction occurs specifically in small intestinal crypt cells that differentiate to become iron-absorbing enterocytes. Our immunohistochemical findings provide evidence for a novel mechanism for the regulation of iron balance in mammals.
H
HFE is a non-classical major histocompatibility complex (MHC) class I molecule that interacts with β2-microglobulin for expression at the cell surface. The widespread C282Y mutation disrupts a disulfide bridge in the α3 helix of HFE, thus preventing its association and allowing the mutant protein to be sequestered within the cell (Feder et al. 1997; Waheed et al. 1997). The occurrence of iron overload in mice lacking β2-microglobulin, which occurs in a pattern akin to that of human hemochromatosis (De Sousa et al. 1994), provides corroborative evidence that the C282Y variant of HFE is responsible for human iron storage disease (Feder et al. 1996). Therefore, plasma and/or endosomal membrane expression of HFE is necessary to maintain normal iron balance. HFE molecules can bind the ubiquitous transferrin receptor (TfR1) in vitro and may influence cellular uptake of transferrin-bound iron (Feder et al. 1998; Gross et al. 1998; Roy et al. 1999). Moreover, HFE can form a ternary complex with TfR1 and its ligand, transferrin, in vitro (Lebron et al. 1998). Mutations in HFE that interfere with the putative TfR1-binding region prevent endocytic internalization of the HFE protein (Ramalingam et al. 2001). However, the physiological relevance of the HFE-TfR1 interaction in living animals and humans is not known.
It is generally accepted that undifferentiated small intestinal crypt cells possess a specific mechanism that determines requirements for transport of iron by mature enterocytes according to body iron status (Conrad et al. 1964; Pollack et al. 1964; Cheng and Le-blond 1974). These stem cells are considered to be subject to functional programming before they migrate along the villus and absorb iron as mature enterocytes. Within the small intestinal epithelium, HFE protein appears to be exclusively expressed in crypt cells (Parkkila et al. 1997). Here, HFE has been implicated in a sensor mechanism by which body iron status regulates iron absorption in mature enterocytes (Griffiths and Cox 2000; Griffiths et al. 2001). We have previously shown that, in the presence of transferrin-iron, HFE co-localizes with TfR1 in an early endosome compartment in cultured human intestinal cells (Griffiths et al. 2000). A report showing apparent co-localization of HFE with TfR1 in small intestinal crypt cells supports an in vivo role for HFE in regulating transferrin-iron uptake into these cells, with subsequent effects on mucosal absorption of iron (Waheed et al. 1999).
We focus here on a novel second transferrin receptor, TfR2, which has been reported to exhibit low-level expression overall in the intestine but is of unknown function (Kawabata et al. 1999). Mutations in the human TfR2 gene cause hemochromatosis, immediately suggesting a key role for the cognate TfR2 protein in iron homeostasis (Camaschella et al. 2000). Two transcripts of the human TfR2 gene have been identified: TfR2-α, predominantly expressed in the liver, and TfR2-β, which encodes a truncated polypeptide. The amino acid sequence of the TfR2-α transcript shows 66% homology to TfR1 in its extracellular domain. In vitro studies demonstrate that the expressed TfR2 protein mediates transferrin-iron uptake into cells but that TfR2-α has an approximately 30-fold lower affinity for holotransferrin than TfR1 (West et al. 2000). Unlike TfR1, TfR2 mRNA lacks iron regulatory elements and TfR2 expression may be regulated by the cell cycle rather than by intracellular iron status (Kawabata et al. 2000).
To test the hypothesis that TfR2 is a key regulatory molecule that interacts functionally with HFE in a specialized pathway controlling iron absorption in mammals, we raised antisera specific for HFE and TfR2 and examined, by confocal microscopy, the localization and molecular interactions of endogenous HFE and TfR2 proteins in human and murine intestinal tissue and cells.
Materials and Methods
Antibody Production and Purification
Polyclonal antibodies were raised to peptide sequences of murine and human HFE and TfR2 proteins in rabbits and chickens. For HFE, peptides were selected from both human and mouse sequences and specificity was ensured by comparison with homologous protein sequences and avoidance of conserved regions. For human HFE-derived peptides, sequences derived from the α1 and α3 extracellular domains of the HFE protein were compared with HLA-A2, HLA-G, and the neonatal Fc receptor (FcRn) (Griffiths et al. 2000). For mouse HFE-derived peptides, derived from α1 and α2 domains, the corresponding human sequences were first compared as for human peptides. For TfR2, a peptide was selected from the human sequence (amino acids 305–320) and compared with TfR1 and prostate-specific membrane antigen (PSMA) for specificity. The equivalent murine sequence was found to be similar, with cross-species reactivity anticipated. Peptides were synthesized and conjugated to keyhole limpet hemocyanin (molar ratio of 50:1 peptide:KLH in 0.05 M NaH2PO4, pH 7.0, at a final concentration of 1 mg/ml) to enhance immunogenicity (ISL; Paignton, UK). Commercial mouse monoclonal antibodies (MAbs) were obtained to human TfR1 (CD71; Lab Vision, Fremont, CA). A mouse MAb to human CD63 (late endosome marker) was kindly provided by Dr. Paul Luzio (Cambridge Institute for Medical Research; Cambridge, UK).
New Zealand White rabbits, weighing 2–3 kg, and hybrid laying hens were immunized using a standardized protocol with KLH-conjugated human HFE peptides (1:1 Freund's complete adjuvant for the initial injection and incomplete adjuvant used for fortnightly boost injections). Further hybrid laying hens were similarly immunized with each of the two KLH-conjugated mouse HFE peptides, and a single rabbit was immunized with the KLH-conjugated TfR2-derived peptide. Serum was extracted from venous samples taken before and after immunization. Blood samples were left overnight at 4C and clot-free liquid pipetted out from around the detached clot into a separate tube. The clot was centrifuged at 2500 × g for 30 min at 4C and the expressed liquid transferred to the tube, which was then centrifuged at 1500 × g for 15 min. Serum was removed and stored in aliquots at −20C.
For purification of serum antibodies from chicken and rabbit, the free peptide was first covalently linked to bovine serum albumin (BSA) and then to cyanogen bromide-activated Sepharose (CL-4B; Amersham, Little Chalfont, UK) to prepare immune-specific antibody by immunoaffinity chromatography as described below. To 6 ml of dissolved BSA (10 mg/ml in PBS, pH 6.0) was added 300 μl of m-maleimidobenzoyl-N-hydroxysuccinimide ester (MBS) in dimethyl sulfoxide (30 mg/ml). The MBS solution was added dropwise under the surface at room temperature (RT), stirring continuously for 30 min. Then 2.1 ml of the mixture was made up to 2.5 ml with PBS (pH 7.4) and added to a pre-equilibrated PD-10 gel-filtration column to remove unbound MBS. To 1.75 ml of eluate was added 5 mg of peptide in 250 μl of PBS, stirring for 1 hr at RT. A gel was then prepared by mixing 1 g of Sepharose 4B in 200 ml of 1 mM HCl (pH 3.0), stirring continuously for 1 hr at RT. The gel was separated through a sintered glass funnel and 1.75 ml of gel was transferred to a 25-ml plastic tube. One ml of peptide/BSA conjugate and 3.5 ml of coupling buffer (0.1 M NaHCO3, 0.5 M NaCl, pH 8.6) was added and the mixture was rotated overnight at 4C. To recover the gel, the suspension was left standing for 10 min and then centrifuged at 1100 × g for 15 min. The gel was incubated in 25 ml of 0.1 M Tris (pH 8.0) for 1 hr at RT to block further linking sites and was retrieved by further centrifugation. To remove non-covalently bound protein, the gel was first mixed with 25 ml of acid buffer (0.1 M NaAc, 1 M NaCl, pH 4.5) for 5 min, washed and retrieved by centrifugation, and then mixed with 25 ml of coupling buffer. This washing cycle was repeated three times and the final supernatant discarded. The gel was resuspended in 9 ml of coupling buffer and 15 ml of immune serum was added. The tube was rotated for 5 hr at RT. After centrifugation, the recovered gel was exhaustively washed with coupling buffer and resuspended in coupling buffer diluted 1:1 with distilled water before transfer to an empty column. Immunoaffinity-purified antibody was eluted with 0.1 M glycine-HCl (pH 2.8) and immediately neutralized with saturated Tris base. The concentration of the antibody solution was noted by measuring the optical density at 280 nm, and the solution was stored in aliquots at −20C, or at 4C after addition of sodium azide (0.1%) to prevent microbial growth.
Human Tissue Samples
Samples of human liver and intestine were obtained by biopsy from patients undergoing investigation for suspected gastrointestinal or hepatic diseases. Written informed consent was obtained and the studies were subject to full institutional ethical review and approval.
Western Immunoblotting
For HFE antibodies, the protein samples used for electrophoresis were a human liver lysate, diluted 1:20 in reducing SDS sample buffer, and mucosal extract derived from mouse duodenum, which was reconsituted after freeze-drying in reducing SDS sample buffer (Laemmli 1970). For the TfR2 antibody, a lysate of K562 cells (erythroleukemic cell line known to express TfR2) was prepared and diluted in 3 × reducing SDS sample buffer. To prepare cell lysates, a single vial of frozen cells was thawed rapidly and centrifuged at 1100 × g for 7 min. The pellet was resuspended in 1 ml of lysis buffer [0.02 M Tris, 0.1 M NaCl, 1 mM EDTA, 1% (v/v) Triton X-100, pH 6.8] containing one tablet of protease inhibitor (Boehringer Mannheim; Mannheim, Germany) per 50 ml and incubated at 4C for 1 hr with intermittent vortexing. The suspension was centrifuged at 10,000 × g for 15 min at 4C and the supernatant stored at −20C. For tissue lysates, 10 mg of frozen human liver or mouse duodenal mucosa was homogenized and lysed for 1 hr at 4C in 0.1 ml lysis buffer as above.
The lysed samples were denatured at 95C for 5 min before being electrophoresed in a 10% SDS-polyacrylamide mini-gel (BioRad; Hercules, CA) with prestained polypeptide molecular weight markers in parallel (BioRad). The polypeptides were transferred in buffer at 4C onto polyvinyl difluoride (Immobilon-P) membranes (Millipore; Bedford, MA). Each membrane was blocked [5% dried milk in buffer containing 0.3 M NaCl, 10 mM Tris base, 0.05% (v/v) 100% Tween-20, pH 7.4 with HCl] and incubated with primary antibody in blocking buffer containing 0.5% dried milk or a control preparation for 1 hr at RT. After three 10-min washes in PBS containing 0.1% (v/v) Tween-20 (PBS-Tween), the membrane was incubated with anti-rabbit or anti-chicken horseradish peroxidase (HRP)-conjugated secondary antibody, diluted according to the manufacturer's instructions (DAKO; Ely, UK). After three 10-min rinses with PBS-Tween, a chemiluminescent substrate (LumiGLO; Amersham) was added for 1 min and the membrane immediately exposed to autoradiographic film.
Confocal Microscopy in Tissues and Cells
The tissue distribution of HFE and transferrin receptors (TfR1 and TfR2) was investigated by confocal microscopy in frozen sections prepared from duodenal mucosal samples from humans and mice. Histologically normal duodenal biopsy specimens were taken with consent, after ethical approval, from two patients with normal iron status and one with hemochromatosis (C282Y homozygous) undergoing routine investigative gastrointestinal endoscopy. Duodenal sections were obtained from four matched pairs of wild-type and HFE knockout mice (kindly supplied by Dr W.S. Sly; St Louis School of Medicine, St Louis, MO). Mice deficient in HFE had been generated by targeted disruption of the murine HFE gene in the region of the C282Y mutation in embryonic stem cells (Zhou et al. 1998). The mutant allele, after embryo transfer, was bred to homozygosity on a svj-129 wild-type background from which the offspring were maintained on regular pelleted chow after weaning. Congenic-strain wild-type mice (Central Biomedical Services; University of Cambridge, UK) were housed and maintained under identical conditions. Animals fasted for 14 hr before sacrifice.
The cellular localization of endogenous iron proteins was examined in human Caco-2 cells, which have a small intestinal phenotype, and in human K562 cells. Caco-2 cells were cultured in Dulbecco's modified eagle medium (DMEM) containing 10% FCS, 2% l-glutamine, and penicillin/streptomycin. K562 cells were cultured in RPMI 16/40 medium supplemented as for DMEM. Undifferentiated human Caco-2 cells were grown to subconfluence on coverslips in 24-well plates. The medium was aspirated, the wells rinsed briefly with PBS, and cells were fixed with 4% paraformaldehyde in PBS for 2 min at RT. Cells were permeabilized in 0.1% Triton X-100 in PBS for 1 min at RT and rinsed with PBS. The cells were then blocked with 10% FCS in PBS-Tween for 1 hr at RT before incubation for a further hour at RT with primary antibody (immune-specific polyclonal or mouse monoclonal) or control antibody preparation diluted 1:100 in PBS-Tween containing 2% FCS. After four 5-min washes in PBS-Tween, the cells were incubated for 1 hr at RT with FITC-conjugated and/or TRITC-conjugated secondary reagents diluted according to the manufacturer's instructions (Jackson ImmunoResearch; West Chester, PA; Sigma-Aldrich, Dorset, UK). An anti-mouse Texas Red secondary antibody was used for CD63 localization (kindly provided by Dr. Paul Luzio). After four further washes, coverslips were mounted in glycerol-PBS (Citifluor; London, UK) and slides examined using laser confocal microscopy (Leica). K562 cells were air-dried onto microscope slides and fixed at −20C in a 2:1 methanol to acetone mixture for 10 min before blocking and staining as above.
For in situ localization of proteins in the duodenal epithelium, fresh-frozen sections were prepared to preserve both morphology and antigen accessibility. Duodena from newly sacrificed animals and human duodenal biopsy specimens were incubated in 2% paraformaldehyde in PBS for 1 hr at 4C. After three rinses in PBS, tissue was incubated in 20% sucrose in PBS for 1 hr at 4C. A glass beaker containing 2-methylbutane (isopentane) was cooled in liquid nitrogen and the tissue frozen first in the isopentane before storing in liquid nitrogen. For immunochemical analysis, slides were thawed and tissue ringed with a wax pen before fixation with 4% paraformaldehyde in PBS for 5 min. Sections were then rinsed with PBS-Tween and blocked with 10% FCS in PBS-Tween for 1 hr, followed by primary antibody (1:100 in PBS-Tween with 2% FCS) or control preparation for 1 hr at RT. Sections were washed and incubated with FITC- and/ or TRITC-conjugated secondary reagents for a further hour (Jackson ImmunoResearch/Sigma), followed by rinsing and mounting for laser confocal microscopy as discussed earlier for the cell preparations.
Results
Intestinal Localization of TfR2 Protein
Antibody specificity was confirmed by Western immunoblotting. Immunopurified rabbit and avian antibodies to human HFE reacted with a single 45-kD polypeptide, the predicted size for HFE. A stronger signal was obtained with the α1 domain-derived peptide antibody (Figure 1A). Antibodies raised against murine HFE peptides were also specific for the HFE protein, particularly the α1 domain-derived peptide antibody, which reacted with a single 47-kD peptide (Figure 1B). A single immunoreactive species estimated at 90 kD (expected size for the TfR2 polypeptide) was observed using immunopurified rabbit antibody to TfR2 (Figure 1C). The specificity of the antiserum for human TfR2 was confirmed independently in the laboratory of Dr. W.S. Sly by Dr A. Waheed. An immunoreactive polypeptide species at the appropriate molecular weight was identified in TRVb-2 cells, which overexpress human TfR2, K562, and Caco-2 cells, with no signal observed in TRVb cells (no expression of TfR1 or TfR2) or TRVb-1 cells expressing human TfR1 (Figure 1D).
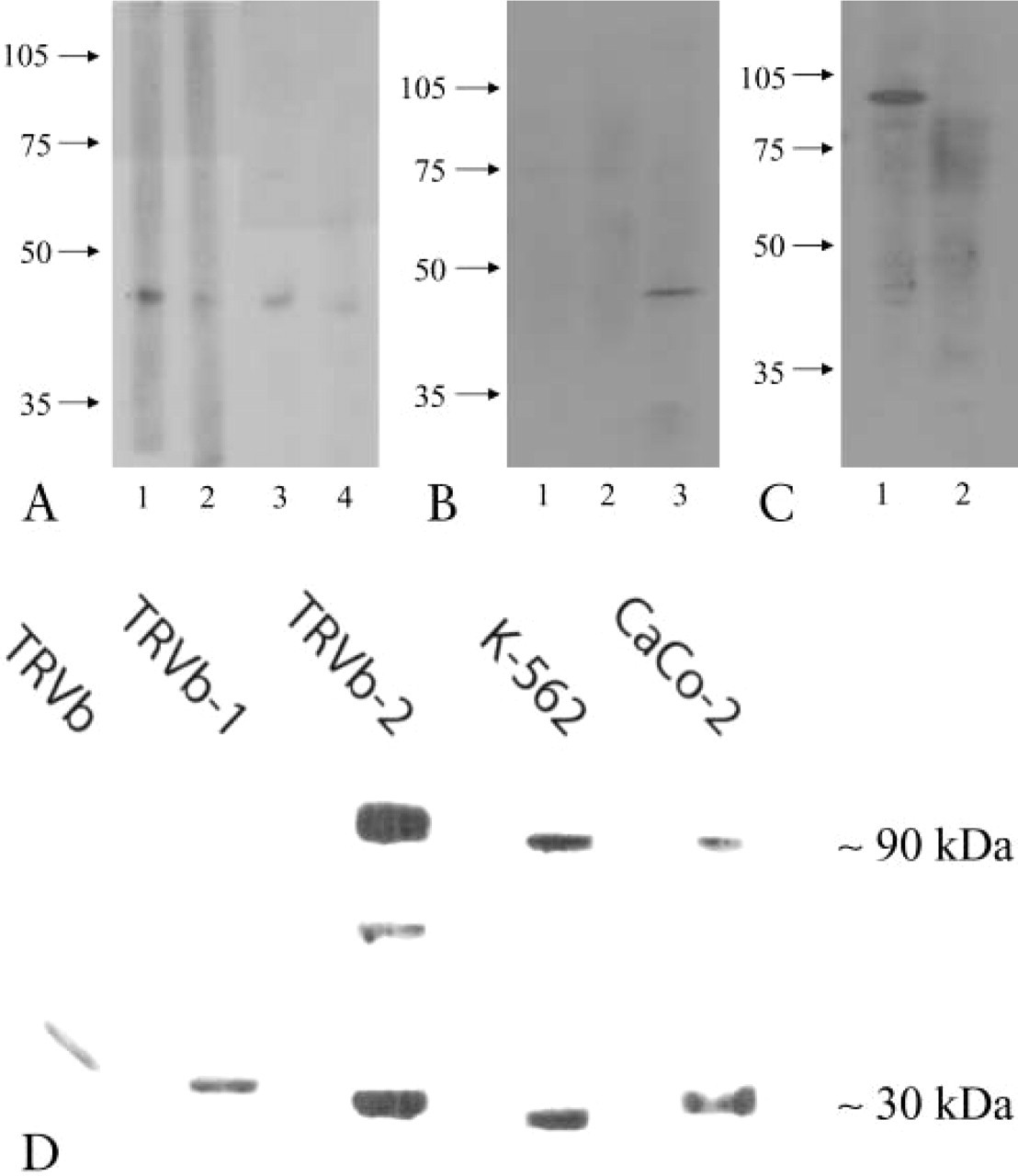
Characterization of antibodies by Western immunoblot-ting. Human liver lysate was electrophoresed and probed with antibodies to human HFE (
Sections of mouse duodenum derived from four wild-type animals were stained for the presence of TfR2. With preimmune serum no signal was observed (not shown), but with antibody to TfR2 there was clear signal restricted to crypt areas of the epithelium (Figure 2A). The crypt-specific localization of TfR2 was confirmed in samples of human duodenal mucosa (Figures 2B and 2C). The antibody does not differentiate between the two transcript products of TfR2, both of which are believed to be expressed in the intestine (Kawabata et al. 1999). HFE was similarly restricted to the duodenal crypts, using the rabbit anti-HFE antibody generated (Figure 2D). The distribution of TfR2 within crypt areas was distinct from that of the ubiquitous TfR1 as demonstrated using a double-staining method (Figures 3A-3C). This further confirms the specificity of the antibody to TfR2 and indicates a specific role and localization for TfR2 that is distinct from that of TfR1 in the intestinal epithelium.
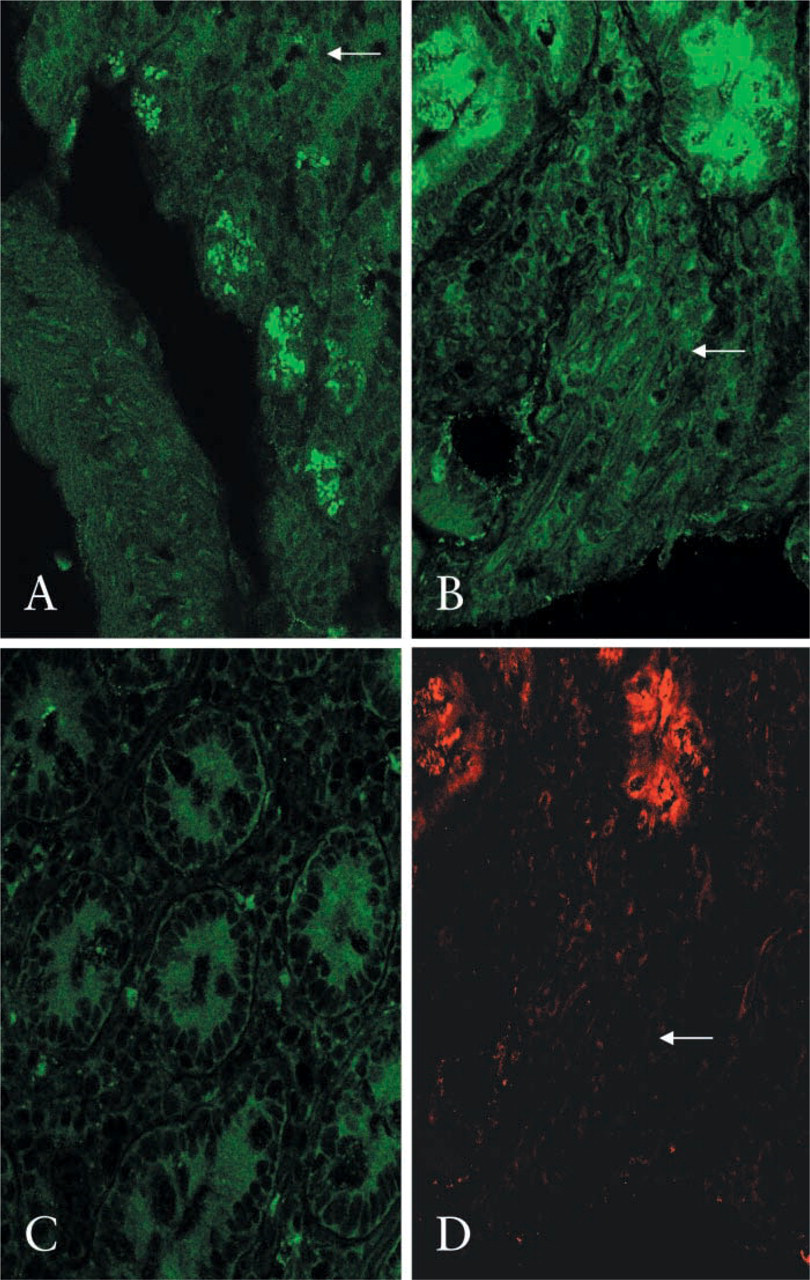
Localization of endogenous TfR2 in mouse duodenum (
In situ localization of HFE, TfR1, and TfR2 in normal human duodenal crypts. Sections were co-stained with rabbit anti-TfR2 and mouse anti-TfR1, followed by anti-rabbit FITC and anti-mouse TRITC (
Interaction of TfR2 and HFE in Duodenal Crypts
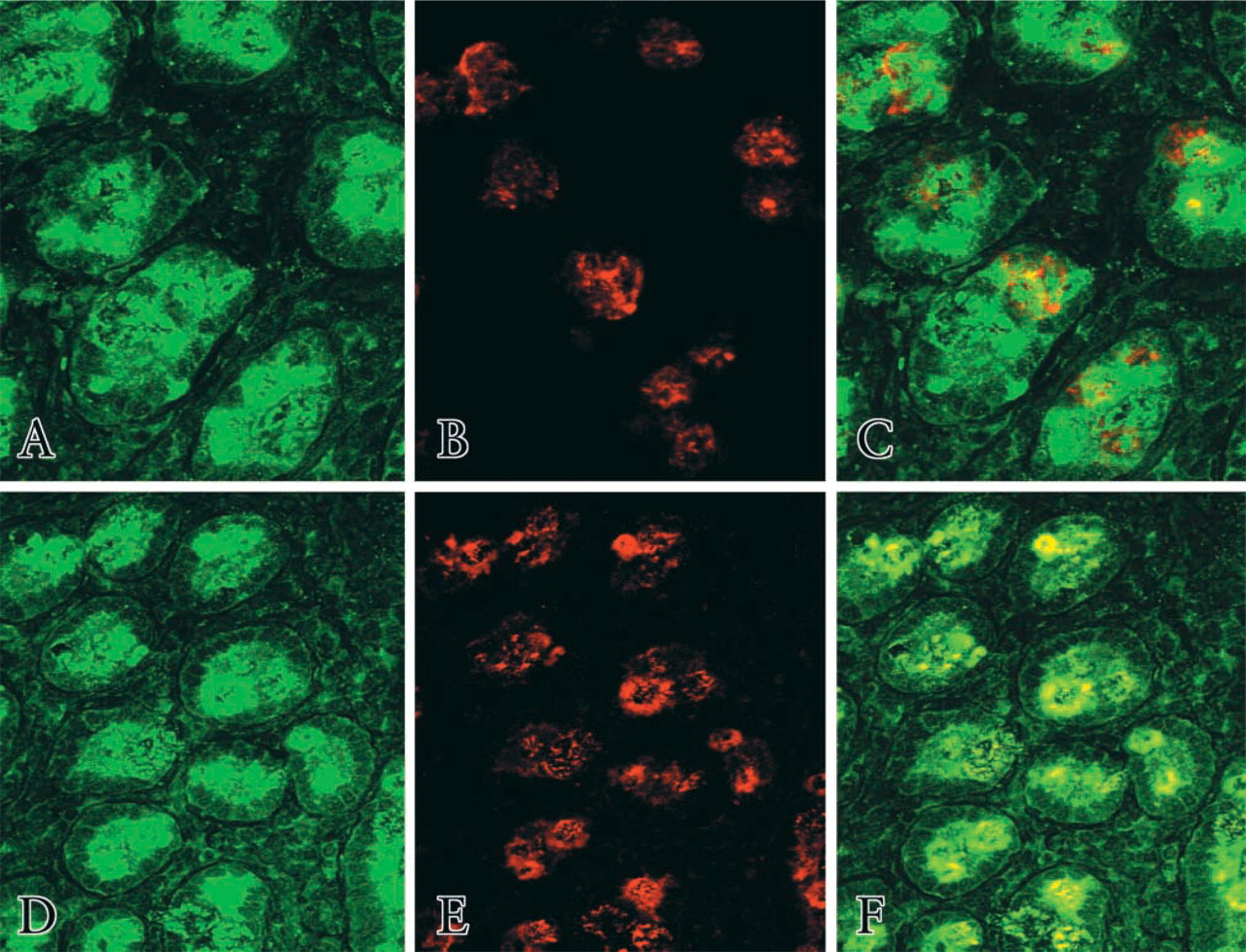
Using an avian antibody to HFE and the rabbit antibody to TfR2, sections of normal human duodenum were examined for potential intracellular interactions of these proteins in situ by confocal microscopy. With a double-immunofluorescence method, TfR2 and HFE were seen to co-localize within duodenal crypts (Figures 3D-3F). These experiments were repeated using duodenal sections derived from a patient with HFE deficiency (C282Y homozygous) and clinical hemochromatosis. Here, HFE staining and the co-localization signal were almost absent (Figure 4A). Furthermore, whereas in wild-type duodenal tissue the distribution of TfR2 appeared mainly intracellular, in the HFE-deficient samples TfR2 localized principally to the plasma cell membrane, particularly the basolateral surface. In mouse duodenum, similar findings were observed. HFE and TfR2 co-localized in crypt areas in the wild-type animals (Figure 4B) whereas in HFE-knockout mice the co-localization signal was absent (not shown).
TfR2 and HFE were localized in duodenal biopsy specimens from a C282Y homozygote with hemochromatosis (
Intracellular Interactions of TfR2 and HFE
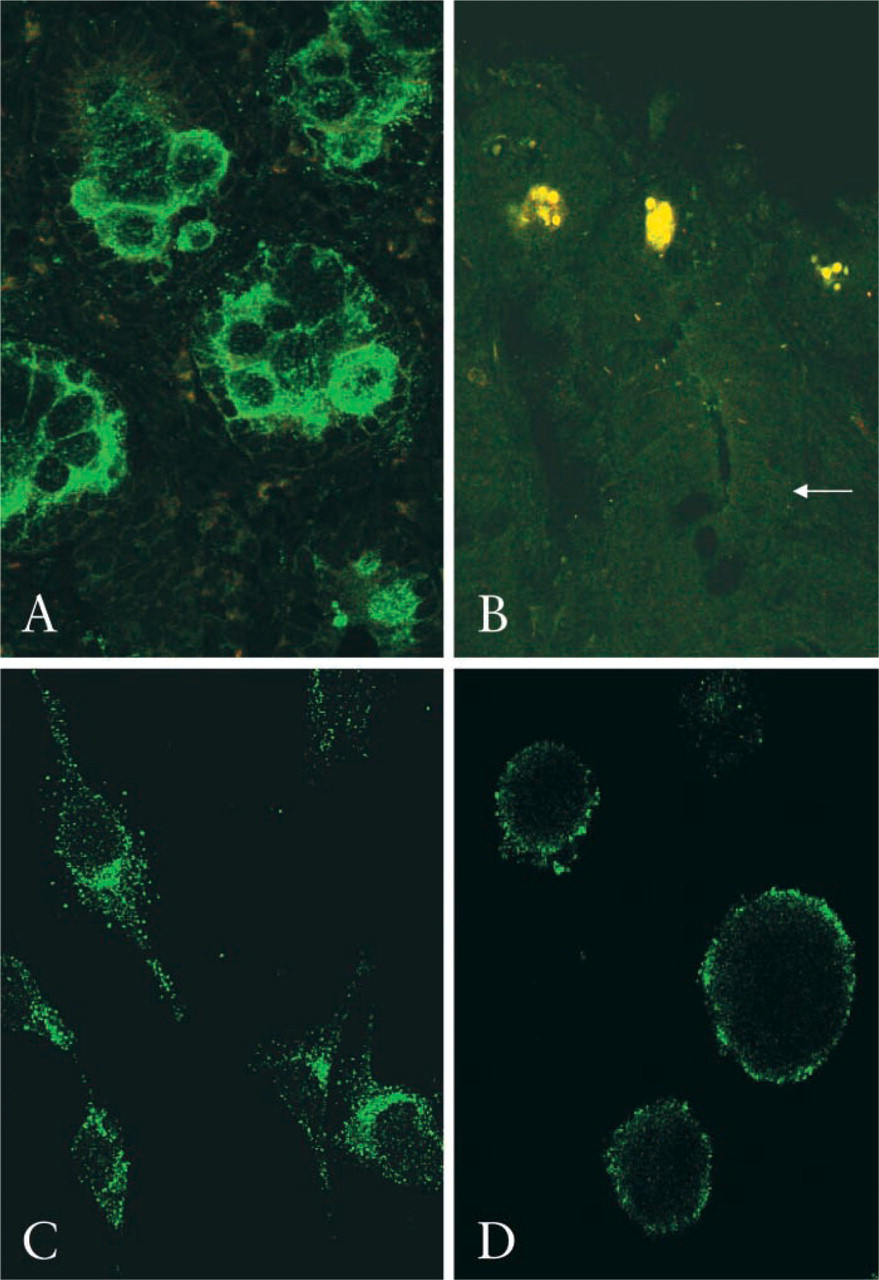
The subcellular localization of the human TfR2 protein was investigated further in the cultured Caco-2 cell line, which has a small intestinal phenotype. In undifferentiated human Caco-2 cells, strong staining of endogenous TfR2 was observed in a vesicular cytoplasmic compartment (Figure 4C). Specificity was confirmed because a greatly reduced signal was observed when the antibody was preincubated with the TfR2 peptide immunogen (not shown). These experiments were repeated in the K562 cell line, in which a similar vesicular staining pattern was observed with TfR2 antibody (Figure 4D). There was no co-localization of TfR2 with either TfR1 or the late endosome marker CD63 in Caco-2 cells (Figures 5A and 5B), suggesting that TfR2 localizes to a distinct early endosomal compartment in these cells. Without additional transferrin ligand, a small population of endogenous TfR2 molecules co-localized with HFE in human Caco-2 cells. However, on exposure to human transferrin-iron there was a marked increase in both the number and the proportion of vesicles showing specific TfR2/HFE co-localization (Figures 5C and 5D).
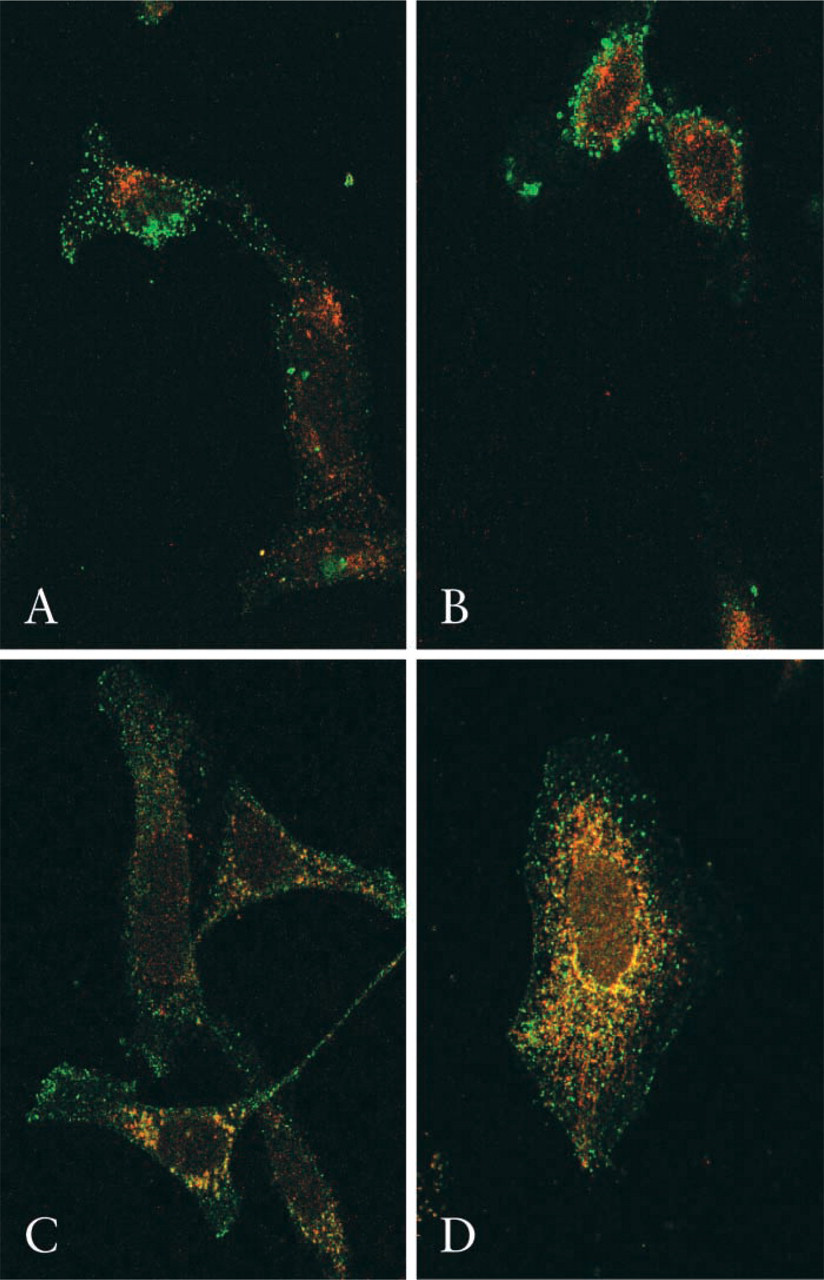
Subcellular localization of endogenous TfR2 in human Caco-2 cells. Undifferentiated Caco-2 cells were co-stained with rabbit anti-TfR2 and mouse anti-TfR1, followed by anti-rabbit FITC and anti-mouse TRITC (
Discussion
The physiological function of the recently identified transferrin receptor homologue TfR2 has hitherto been unknown, but the involvement of TfR2 in iron homeostasis, as demonstrated by the occurrence of iron storage disease (HFE3) in patients carrying disabling mutations in the human TfR2 gene (Roetto et al. 2001; Girelli et al. 2002; Mattman et al. 2002), has immediately focused attention on this novel protein of iron transport. Here we show for the first time that TfR2 localizes specifically to intestinal crypt cells in human and mouse duodenum. The pattern of expression of TfR2 is quite distinct from the distribution of TfR1 protein expression along the entire villous axis (Lombard et al. 1990). We further show here that TfR2 co-localizes with wild-type HFE protein in mammalian crypt cells that constitute a renewing population of intestinal villous cells involved in the uptake and absorption of dietary iron. In cultured human Caco-2 cells, addition of transferrin-iron markedly stimulates intracellular interactions between endogenous TfR2 and HFE within a vesicular compartment. These vesicles are likely to represent a distinct subpopulation of endosomes involved in the uptake and recycling of transferrin-iron in the intestinal crypts.
Previous studies have shown that HFE may interact with TfR1 in vitro. When overexpressed, HFE negatively modulates TfR1-mediated iron uptake in cultured HeLa cells. Therefore, mutations in HFE might be expected to increase intracellular iron stores in crypt cells if a similar HFE-TfR1 interaction predominates at this site. However, enterocytes have been shown to be paradoxically iron-deficient in hereditary hemochromatosis (Francanzani et al. 1989; Pietrangelo et al. 1995), and uptake by the duodenal epithelium of parenterally administered radiolabeled transferrin-iron was reduced in HFE knockout mice compared with wild-type (Trinder et al. 2002). Additional recent studies have demonstrated increased transferrin receptor-dependent iron uptake in cultured Chinese hamster ovary cells after concomitant overexpression of HFE and β2-microglobulin (Waheed et al. 2002). These experiments therefore support positive modulation of transferrin-iron uptake by wild-type HFE in the intestine. We now show that within the duodenal epithelium TfR2, but not TfR1, is selectively expressed in crypt cells. Our confocal studies further demonstrate a population of vesicles containing the novel TfR2 protein that is negative for CD63 antigen, a marker for late endosomes (Metzlaar et al. 1991). In the presence of iron-saturated transferrin, endogenous HFE co-localizes with these TfR2-positive vesicles, and marked attenuation of the co-localization signal in the presence of C282Y-mutated protein further implicates HFE in this membrane compartment.
In the light of these findings, we propose that HFE functionally interacts with TfR2 within this early endosomal compartment specifically to enhance receptor-mediated iron entry into small intestinal crypt cells. We further propose that the transferrin ligand is of central importance in informing crypt cells about body iron status. Iron absorption is inversely related to the level of transferrin saturation in serum (Taylor and Gatenby 1966), and inherited transferrin deficiency is associated with increased absorption of iron, correctable with parenteral transferrin, suggesting a reversible failure of homeostatic signaling within the intestine (Heilmeyer et al. 1961; Raja et al. 1999). Lately, increased duodenal DMT1 protein expression has been demonstrated in hypotransferrinemic mice, the same downstream upregulation response found in HFE deficiency (Canonne-Hergaux et al. 2001; Griffiths et al. 2001). Therefore, the steady-state iron saturation of plasma transferrin may directly determine the delivery of iron to proliferating crypt cells in the upper small intestine, thereby signaling an instantaneous evaluation of iron demand. It is recognized that iron-deficient erythropoiesis in the hypotransferrinemic mouse may directly influence iron absorption, but the mechanism for this is not known. Mutations in the human HFE gene, such as the C282Y mutation examined here, may therefore disrupt crypt HFE/TfR2 interactions, thus affecting the capacity of serum transferrin-iron to influence molecular programming of the DMT1 iron uptake pathway in mature enterocytes.
HFE is therefore likely to have a specific effect on iron entry into crypt cells, which may be mediated by a distinct pathway that is independent of the ubiquitous transferrin receptor (TfR1) isoform and that has a specialized function in iron signaling and homeostasis. Within the mucosal epithelium, HFE and TfR1 appear to interact in a minor tissue compartment, as reported previously (Griffiths et al. 2000), and colocalization has been demonstrated in the crypts (Waheed et al. 1999). TfR1 may have only a housekeeping role. Indeed, we suggest here that the TfR1 mechanism mediates the high-affinity uptake of iron from the plasma solely to meet intrinsic metabolic requirements of enterocytes. TfR1 protein expression is directly regulated by intracellular iron concentrations, as a result of interactions with iron regulatory protein, thus facilitating rapid homeostatic responses to changes in cellular iron demand (Hentze et al. 1989). Compared with TfR1, TfR2 has a relatively low affinity at physiological pH for iron-containing transferrin, and therefore an endocytic pathway for transferrin-iron uptake mediated by TfR2 (and HFE) would be highly sensitive to changes in the saturation level of plasma apotransferrin. Preferential co-localization of HFE with TfR2 supports this as the principal iron ligand/receptor compartment in the intestinal crypts. A specialized endosomal transport pathway in the intestine, of which HFE and TfR2 are key components may, according to body iron status, alter crypt cell iron concentrations and thereby modulate expression of rate-limiting iron-transport proteins, such as DMT1, by iron-regulatory protein interactions.
Although we believe that the finding of HFE/TfR2 co-localization within an intestinal cell vesicular compartment represents an important step in understanding the components of the regulatory mechanism for iron homeostasis, few mechanistic insights as to how these molecules interact functionally within cells are available. West et al. (2000) were unable to demonstrate direct binding of HFE to TfR2 in vitro. However, binding at hydrogen ion concentrations other than pH 7.5, e.g., acidic endosomal conditions, was not evaluated. Furthermore, the truncated protein constructs generated in their overexpression systems may have had altered binding characteristics compared with the native proteins. The authors also showed that residues in TfR1 involved in HFE binding were not conserved in TfR2. However, these same residues may not necessarily be involved in interactions between HFE and TfR2. The overlapping co-localization signal of endogenous HFE and TfR2, observed in mammalian crypt cells and in human Caco-2 cells, suggests close proximity of these molecules in situ, and our findings indicate the presence of these proteins within the same vesicular compartment. Although powerful in demonstrating the behavior of these proteins in vivo, the precise nature of any interactions between HFE and TfR2 cannot be resolved solely by microscopic methods. Questions remain as to how a signaling pathway involving HFE and TfR2 might operate at a molecular level, and studies are required to address this, e.g., further evaluation of the recently described TfR2-deficient mouse, which develops a pattern of iron overload akin to HFE deficiency (Fleming et al. 2002). The role of TfR2 in iron homeostasis may include sites other than the intestine and, in particular, TfR2 may regulate hepatocyte iron uptake, because it has been reported that in HFE knockout mice hepatic TfR2 mRNA expression is not downregulated and remains high (Fleming et al. 2000).
In the context of these findings, HFE may serve to promote transferrin-iron uptake by TfR2 into the crypt cell cytoplasm or to another specialized transport system within recycling endosomes. A key regulatory function of the transferrin ligand-receptor pathway implicated in our hypothesis is confirmed by the persistently increased uptake and absorption of iron by the intestine in hpx mice and in hypotransferrinemic humans, despite the development of marked systemic iron storage disease (Heilmeyer et al. 1961; Simpson et al. 1991). Mutations in HFE, TfR2, or transferrin would reduce crypt cell uptake of transferrin-iron and lead to inappropriate upregulation of enterocyte iron transport proteins with toxic accumulation of iron in tissues. Thus, an early endosomal compartment within mammalian crypt cells involving HFE and TfR2, distinct from that which involves TfR1, may constitute a selective transport pathway for the delivery of transferrin-iron to the intestine. We propose that this signaling pathway influences the subsequent absorption of iron by mature enterocytes and is therefore critical for the maintenance of iron homeostasis.
Footnotes
Acknowledgements
Supported by a Wellcome Trust Clinical Training Fellowship (WJHG), a Sackler Award (WJHG), and European Union funding.
We are grateful to Dr A. Waheed and Dr W.S. Sly for their assistance in confirming the specificity of our TfR2 antibody.