
Abstract
Fixation-resistant NADPH-diaphorase (NADPHd) activity is used widely as a marker for nitric oxide synthase (NOS). In frozen sections, NADPHd histochemistry yields high anatomic definition. In whole-mounts, however, poor penetration of the reagents, background staining, and tissue opacity severely limit its application. Here we report a combination of new methods that significantly improves whole-mount NADPHd staining. We demonstrate these methods in the thoracic ganglia of a large insect, the locust Schistocerca gregaria, in which NADPHd has been analyzed previously using both whole-mounts and serial section reconstructions. The penetration of the staining reagents was markedly improved after fixation in methanol/formalin compared to phosphate-buffered formaldehyde. Methanol/formalin also reduced nonspecific NADPHd and enhanced the selective staining. Penetration was further enhanced by incubation regimens that exploit the temperature- or pH-dependence of NADPHd. In combination with methanol/formalin fixation, this permitted staining to develop evenly throughout these comparatively large invertebrate ganglia. These improvements were complemented by a new clearing technique that preserves the NADPHd staining, gives excellent transparency, and avoids distortion of specimen morphology. The new methods revealed the three-dimensional architecture of NADPHd expression in locust ganglia in unprecedented detail and may similarly improve wholemount detection of NADPHd in other invertebrate and vertebrate preparations.
Keywords
NADPH-diaphorase (NADPHd) activity in formaldehyde-fixed tissue can be largely attributed to nitric oxide synthase (NOS), the enzyme responsible for biosynthesis of the diffusible free radical messenger molecule nitric oxide (NO; Dawson et al. 1991; Hope et al. 1991; Masumoto et al. 1993; Elphick et al. 1994; Müller 1994; Müller and Bicker 1994). This discovery led to the extensive use of the NADPHd staining reaction as a marker for NOS after almost three decades during which its functional significance remained elusive (Thomas and Pearse 1964; Scherer-Singler et al. 1983). The NADPHd technique is sensitive, simple and inexpensive, but it demands careful interpretation, (a) because the NADPHd activity of enzymes other than NOS can give false-positive staining (e.g., Vincent and Kimura 1992 vs Kishimoto et al. 1993; Schürmann et al. 1997 vs Ott and Burrows 1999; Chiang et al. 2000 vs Skinner et al. 2000) and (b) because in certain invertebrates the apparent formaldehyde sensitivity of NOS-related NADPHd can result in false-negative staining (Ott and Burrows 1999; Ott and Elphick 2002). Antibodies against mammalian NOS isoforms are now widely available and enable localization of NOS by immunohistochemistry, but in invertebrates they may yield poor results (e.g., Elphick et al. 1995) or fail altogether (e.g., Schürmann et al. 1997). More recent studies have therefore used “universal” NOS (uNOS) antibodies that target the conserved amino acid motif QKRYHEDIFG of NOS (e.g., Gibbs and Truman 1998; Gibson and Nighorn 2000; Skinner et al. 2000; Zayas et al. 2000; Newcomb and Watson 2001). We have shown that, in the brain of the locust and the cockroach, strongly NADPHd-positive neurons are also immunopositive with uNOS antibodies (Ott and Elphick 2002). However, the sensitivity and anatomic resolution of the antibody staining were considerably poorer than that of NADPHd. In the cockroach, diffuse immunoreactivity putatively unrelated to NOS abounded. In the locust, the uNOS antibody revealed no arborizations except in the antennal lobes, a region of very high NOS activity. Seidel and Bicker (2002) have relied on NADPHd to study the embryonic expression of NOS in the locust “… because the antiserum does not resolve very clearly the fine neurites whereas the NADPHd staining resolves sharply the neuronal arborizations….”
Even when NOS antibodies yield well-defined staining, NADPHd is a valuable tool for independent evaluation. For example, affinity-purified uNOS antibodies give positive staining in locust skeletal muscle but no corresponding NADPHd staining has been found, suggesting that in locust muscle the uNOS antibodies may crossreact with material other than NOS (Ott et al. 2000). The NADPHd method therefore permits (a) the detailed anatomic analysis of NOS expression, particularly when antibodies of high specificity and affinity are not at hand, and (b) the validation of results obtained with anti-NOS immunohistochemistry.
Although high anatomic detail usually requires the NADPHd staining to be performed on frozen sections, it has also been applied to unsectioned specimens (wholemounts; e.g., Xenopus tadpole central nervous system, McLean and Sillar 2001; muscularis externa of rat intestine, Santer 1994; rabbit retina, Zemel et al. 2001). This whole-mount approach offers particular advantages in invertebrate nervous systems (e.g., Moroz et al. 1993; Müller and Bicker 1994; Bascal et al. 1995; Elphick et al. 1995; O'Shea et al. 1998; Sadamoto et al. 1998; Zayas et al. 2000; Newcomb and Watson 2001), first, because of their smaller size, and second, because many invertebrate neurons are individually identifiable by their branching morphology, which is more readily recognized in whole-mount preparations than in sections. However, three factors impose severe limitations on NADPHd staining in whole-mounts: (a) the staining reagents penetrate the tissue very slowly; (b) NOS-unrelated NADPHd staining adds up over the thickness of the preparation and interferes with the observation of the specific staining; and (c) clearing the whole-mounts for microscopic examination is a compromise between tissue transparency and preservation of staining (Elphick 1997). These three problems are mutually aggravating, particularly in larger preparations, in which whole-mount NADPHd reveals only structures close to the surface and thus provides an incomplete picture of NOS expression. Here we report a combination of novel techniques that reveals NADPHd-expressing neurons with unprecedented anatomic resolution in intact tissue. We demonstrate these techniques in the metathoracic ganglion of the locust Schistocerca gregaria, a comparatively large preparation in which NADPHd expression has previously been analyzed using both whole-mounts (Müller and Bicker 1994) and series of frozen sections (Ott and Burrows 1998).
Materials and Methods
Adult locusts (Schistocerca gregaria, gregarious phase) were bought from Blades Biological (Cowden, Kent, UK). Reagents were obtained from either Sigma-Aldrich, Poole, UK or BDH Chemicals, Poole, UK, unless stated otherwise. Thoracic nerve cords were dissected out in HEPES-buffered saline (Ott and Elphick 2002) at room temperature (RT) and transferred into ice-cold saline for short-term storage (2 hr maximum) until they were fixed. Throughout this work, pH 2 refers to the value at RT, regardless of the temperatures at which the solutions were used.
Fixation
Two different fixatives were compared, buffered formaldehyde (BF) and methanol/formalin (MF).
BF Fixation
Buffered formaldehyde (BF; 4% formaldehyde in 0.1 M phosphate buffer, pH 7.4) was prepared from paraformaldehyde powder, aliquotted, and stored at −20C; aliquots were thawed immediately before use. Ganglia were fixed in BF for 2 hr at 4C and then washed twice for 15 min in PB at 4C. This was followed by overnight permeabilization on ice in 0.2% Triton X-100 in 0.1 M Tris buffer, pH 8.0 (Tris-Tx).
MF Fixation
Methanol/formalin (MF; Ott and Elphick 2002) is a mixture of 9 volumes absolute methanol and 1 volume formalin (37% commercial formaldehyde solution containing approximately 10% methanol; Sigma F-1635, Poole, UK), resulting in approximately 91% methanol and 3.7% formaldehyde; the mixture is stable at RT. Ganglia were fixed in ice-cold MF for 2 hr and then washed twice for 15 min in ice-cold 0.1 M Tris buffer, pH 7.2. The strong glial NADPHd present in locust ganglia after MF fixation was blocked by low pH according to Ott and Elphick (2002), with the following modifications. Ganglia were incubated on ice at pH 4.0 (0.1 M acetate buffer) for 60 min and then overnight at pH 5 in the presence of 0.2% Triton X-100. They were then washed again for 15 min in ice-cold Tris buffer, pH 7.2.
Whole-mount NADPHd Staining
Three alternative NADPHd staining regimens were used: direct staining, staining after cold preincubation, and staining after acidic preincubation.
Direct Staining
Ganglia were directly incubated in an NADPHd staining solution of 0.2 mM β-NADPH and 0.2 mM nitroblue tetrazolium (NBT) in Tris-Tx for 1–3 hr at RT.
Staining After Cold Preincubation
Ganglia were incubated in the same NADPHd staining solution as for direct staining, but on ice rather than at RT, for 3 hr under constant agitation. Only very weak staining developed during this preincubation due to low enzyme activity at 0C. Staining was then triggered by bringing the preparations to RT and was allowed to develop for 30–60 min.
Staining After Acidic Preincubation
This is effectively a modified cold preincubation that is carried out at low pH to further reduce enzyme activity. Overnight incubation in acetate buffer, pH 5.0, was omitted. Instead, ganglia were pre-incubated overnight on ice with 0.2 mM NADPH and 0.2 mM NBT in 0.1 M Sørensen phosphate buffer, pH 5.0, containing 0.2% Triton X-100 (phosphate buffer was used because NADPH appeared unstable in acetate buffer). Development was then triggered by placing the tissue in the pH 8.0 staining solution (see above; 0.1 M Tris buffer) at RT.
Clearing of Whole-mounts
The staining reaction was stopped by extensive washes in distilled water. Whole-mounts were then cleared using one of the following three approaches: glycerol clearing, ethanol/xylene clearing, and methanol/cedar oil clearing.
Glycerol Clearing
Ganglia were cleared and subsequently mounted on slides, in a 9:1 mixture of glycerol and 0.1 M phosphate buffer, pH 7.4 (e.g., Müller and Bicker 1994; Bascal et al. 1995).
Ethanol/Xylene Clearing
Ganglia were mounted on slides in DPX after dehydration in ethanol and clearing in xylene or methyl salicylate (e.g., Moroz et al. 1993; Elphick 1997; O'Shea et al. 1998; Sadamoto et al. 1998; Zayas et al. 2000). Because nitroblue formazan (NBF) is soluble in these agents, the exposure had to be kept to the necessary minimum.
Methanol/Cedar Oil Clearing
We developed this new technique to avoid the shortcomings in the two above approaches. Ganglia were transferred directly into a freshly prepared 3:1 mixture of absolute methanol and glacial acetic acid for 15–30 min, followed by three times for 15–30 min in absolute methanol and 30–60 min in cedar oil (Cedarwood Oil for Clearing; RA Lamb, London, UK). Preparations were then mounted permanently in cedar oil on cavity slides.
Frozen Sections
Some ganglia that had been whole-mount-stained for NADPHd were subsequently sectioned rather than cleared. These ganglia were cryoprotected at RT in 20% sucrose in 0.1 M phosphate buffer containing 0.005% sodium azide, embedded in Jung Tissue Freezing Medium (Leica; Nussloch, Germany), and frozen on the surface of liquid nitrogen. A Leica CM1800 cryostat was used to cut 30-μm frozen sections, which were collected on chromalum/gelatin-coated slides and mounted in glycerol jelly.
Imaging
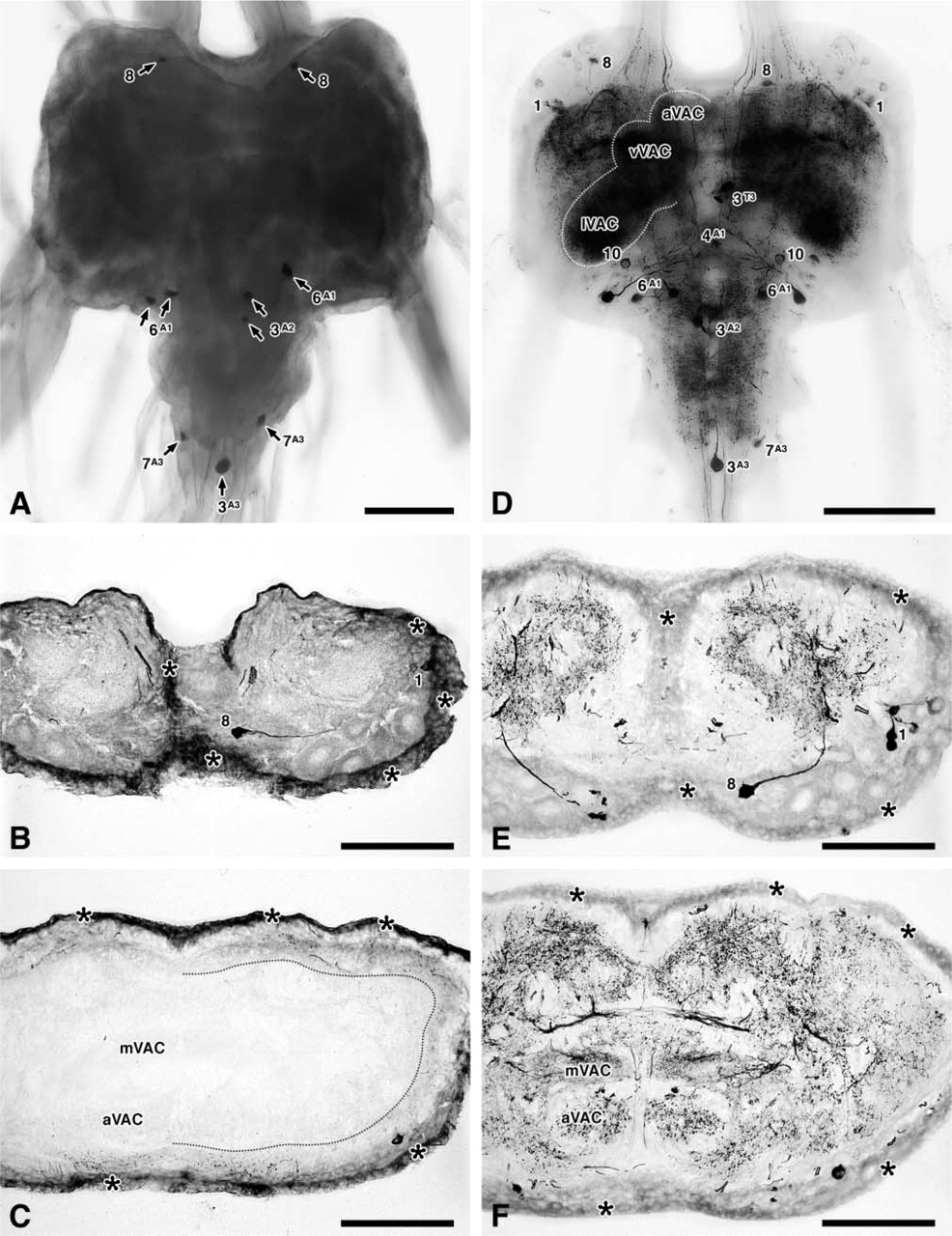
Whole-mounts and sections were viewed under a Leica DMRA2 compound microscope fitted with a motorized Z-stage. Digital images were captured using a Retiga 1300 12-bit monochrome camera (QImaging; Burnaby, BC, Canada) and QCapture 1.1.6 (QImaging) or OpenLab 3.0.8 (Improvision) software running on a Macintosh G4 computer (Apple Computer; Cupertino, CA). Color images were acquired using the camera in combination with an RGB Liquid Crystal Color Filter module (QImaging). All further processing was done in ImageJ 1.27z (Wayne Rasband, National Institutes of Health, <http://rsb.info.nih.gov/ij/>; ImageJ is in the public domain) and Photoshop 6.0 (Adobe Systems; San Jose, CA) software. For Figures 1A and 1D, the captured 12-bit images were log-transformed before mapping their dynamic range to 8 bit; multiple focal planes were combined in Photoshop using the “Darken” layer mode. For Figures 1B and 1C, 1E and 1F, the dynamic range of the captured 12-bit images was mapped linearly to 8 bit.
Comparison of conventional (
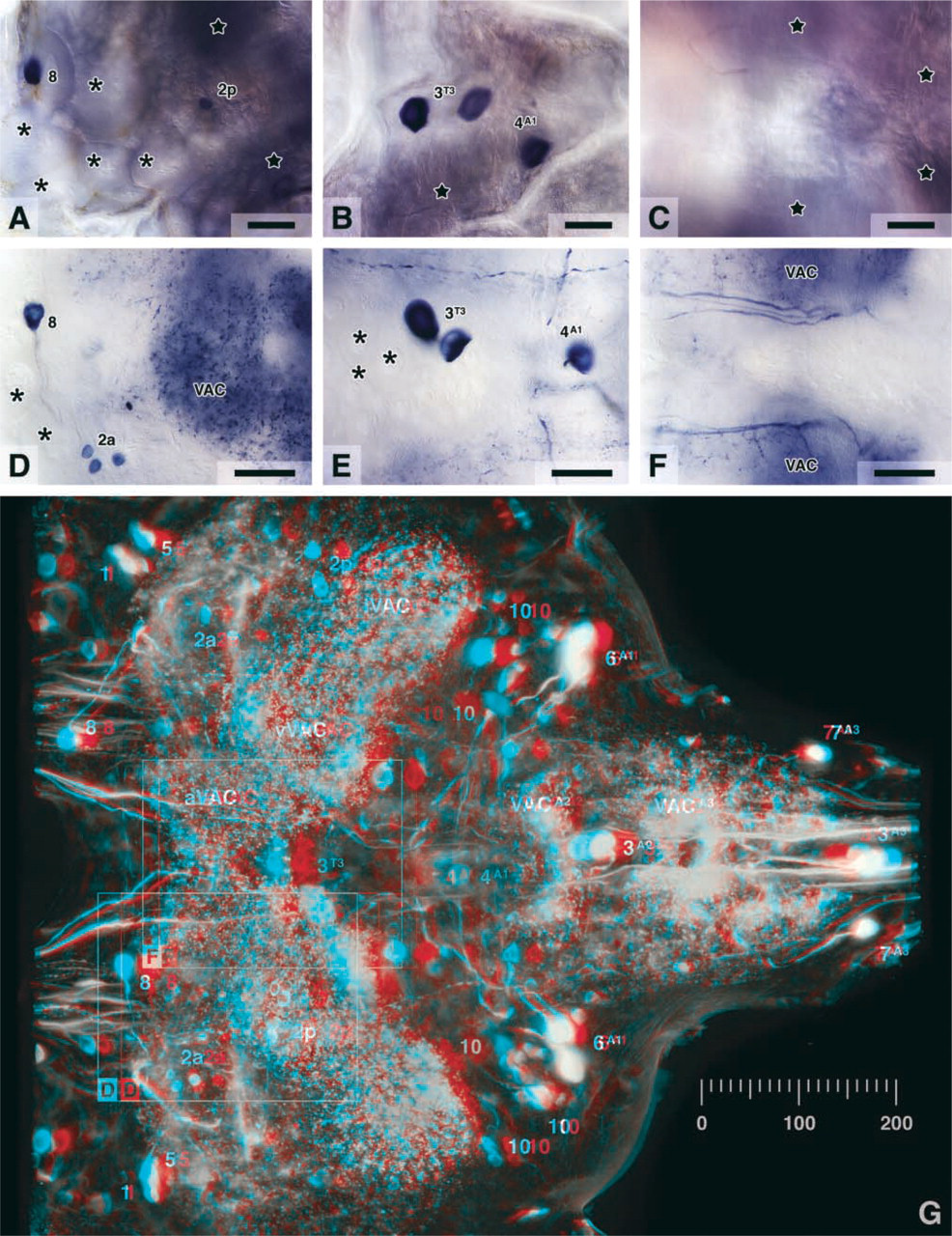
The 3D image (anaglyph) in Figure 2G was constructed from 75 focal planes taken at 5-μm intervals along the Z-axis (dorsoventral axis of the ganglion). The XY-resolution was increased by using a X20 objective and capturing six Z-stacks that overlapped in the XY-plane and together covered the entire ganglion. The 12-bit camera frames were log-transformed and downsampled to 8-bit resolution, and each set of six frames belonging to one focal plane was merged into a single high-resolution image. Next, each focal plane was brightness-inverted, and out-of-focus blur was reduced by Rolling Ball background subtraction (75 pixel radius) in ImageJ. Labels were applied to individual focal planes. The anaglyph was then constructed from two Brightest Point projections that simulate a binocular ventral view with an interocular viewing angle of 12°.
(
Results and Discussion
NADPHd histochemistry in whole-mounts is severely limited by the poor penetration of the staining reagents NADPH and NBT, by NOS-unrelated NADPHd, and by tissue opacity. Naturally, these limitations are the more pronounced the thicker the specimens are. Background, for example, adds up over the thickness of the preparation and is therefore much more critical in whole-mounts than in sections. The problems are readily demonstrated in the metathoracic ganglion of the locust Schistocerca gregaria. In adult locusts, this ganglion measures about 1800 × 1400 × 800 μm (unfixed). In addition to its considerable size, two previously published studies make this preparation a very instructive test object. The first report on NADPHd in locust thoracic ganglia was based on whole-mount preparations that showed stained neuronal cell bodies (Müller and Bicker 1994). Since that report, the branching morphologies of these and several other NADPHd-positive neurons have been reconstructed in detail from serial frozen sections (Ott and Burrows 1998). In this preparation, therefore, the improvements in whole-mount staining afforded by the new procedures can be gauged both against the results obtained with previously available techniques and against the much more detailed picture that has emerged from rather laborious serial reconstructions. Throughout this article, the numerical nomenclature of Ott and Burrows (1998) is used to refer to previously identified NADPHd-positive neurons or groups of neurons. In the metathoracic ganglion, cell group “2” is not homogeneous (Ott and Burrows 1998). It comprises an anterior and a posterior population of neurons with different projections, here referred to as “2a” and “2p,” respectively.
Whole-mount NADPHd After BF Fixation
Figure 1A shows a typical example of the whole-mount staining in the metathoracic ganglion of an adult locust after application of “conventional” techniques (BF fixation, direct staining, glycerol clearing; for details see Materials and Methods). This experiment closely replicates the results of Müller and Bicker (1994; cf. their Figure 2a) and reveals the same set of cell bodies (arrows in Figure 1A). The fact that the ganglion appears considerably darker than that in Figure 2a of Müller and Bicker (1994) is largely a reflection of the linear response of the CCD camera vs the nonlinear response of photographic film. CCD imaging thus amplifies further the problem of background in whole-mount preparations. Stained cell bodies are more readily seen when the exposure of the CCD is increased (Figures 2A and 2B; same preparation as in Figure 1A), but at these settings the lighter parts of the preparation (lateral nerves, connectives, and the margin of the ganglion) would be far outside the dynamic range of the CCD. For example, interneuron “8” (which corresponds to “S” in Figure 2a of Müller and Bicker 1994) is difficult to make out at exposure settings that do not swamp the CCD chip at the margin of the ganglion (“8” in Figure 1A), but is clearly visible in Figure 2A.
Like Müller and Bicker (1994), we were unable to reveal the neuropilar arborizations of NADPHd-positive neurons (except in the tail end of the third abdominal neuromere where the ganglion peters out into the posterior connectives). It is important to note that the dark appearance of Figure 1A and the dark regions in Figures 2A–2C (stars) are caused exclusively by neuronal background and non-neuronal staining near the surface of the ganglion, and not by selective fiber staining in the neuropil. Frozen sections cut after whole-mount staining (Figures 1B and 1C) show very clearly the intense label in a superficial layer of non-neuronal cells known as the perineurium (Figures 1B and 1C, asterisks). This perineurial NADPHd is in all probability unrelated to NOS (Ott and Elphick 2002). Figure 1B also shows that considerable background has developed in neuronal cell bodies and in neuropil. Perineurial staining and neuronal background thus combine to obscure the selective NADPHd in whole-mounts. Moreover, sectioning after whole-mount staining shows that the penetration of the staining reagents is extremely poor. The section in Figure 1B cuts tangentially through the anterior cortex. Here, background extends throughout the section, evidence that the staining reagents have reached the tissue. In the more central section in Figure 1C, however, such background occurs only in a narrow zone along the surface: the reagents have penetrated only some 70 μm into the tissue. Consequently, arborizations of NADPHd-expressing neurons are also revealed only within this superficial layer (dotted line in Figure 1C). The best part of the neuropil, including the mechanosensory ventral association centers (VACs), which are known to contain very dense aggregations of NADPHd-positive fibers (Ott and Burrows 1998), is completely devoid of any staining. The neuropil was still unstained after several hours of development at RT. By that time, nonspecific staining near the surface had reached levels that made whole-mount observation impossible.
Whole-mount NADPHd After MF Fixation
Poor penetration and background are thus mutually aggravating because the latter is amplified by the long incubations necessitated by the former. This prompted us to explore whether fixation in methanol/formalin (MF) might improve the results in whole-mounts. We have recently developed MF fixation as a means to preserve the strongly formaldehyde-sensitive NOS/NADPHd of the cockroach Periplaneta americana (Ott and Elphick 2002). Although MF fixation has thus far been used only for detection of NADPHd in frozen sections, we expected that MF might facilitate the penetration of the NADPHd staining reagents in intact tissue because it coagulates the cytoplasm and increases the permeability of the cell membranes. Furthermore, MF has also been shown to improve the sensitivity and selectivity of NADPHd staining in sectioned locust brain (Ott and Elphick 2002), which should help combat the background problems in whole-mounts.
MF fixation yielded pronounced improvements in the detection of NADPHd in intact thoracic ganglia. First, the staining extended much further into the neuropil, reflecting the facilitated penetration of the staining reagents in MF-fixed tissue. However, direct staining (as defined in Materials and Methods) still failed to reveal the NADPHd-positive fibers in the core of the ganglion with incubation times that avoid overstaining the periphery (not shown). Only by combining MF fixation with cold or acidic preincubation was it possible to reveal the NADPHd fiber architecture evenly throughout the ganglion (Figure 1D and 2D–2G; the fainter appearance of the staining “inside” the image in Figure 2G is largely an artifact of the Rolling Ball background subtraction used to reduce out-of-focus blur). The rationale behind cold preincubation is that diffusion is linearly dependent on temperature, whereas enzyme reactions have higher-order thermal kinetics. Low temperatures thus favor diffusion over enzyme activity and allow incubation in the staining reagents without concomitant development of staining. Slight staining that may develop during prolonged cold preincubation can be completely suppressed by acidic preincubation, which also exploits the pH dependency of NOS enzyme activity (cf. Spessert and Claassen 1998; Ott et al. 2001). However, equivalent results were obtained with the simpler cold preincubation technique. The improvement in penetration of the staining reagents over conventional methods is evident from Figure 2F, which shows a central section cut after whole-mount staining (cf. result after conventional processing, Figure 2C).
Second, and equally important, the signal-to-background ratio in intact ganglia was far better after MF than after BF fixation. NADPHd-positive cell bodies and a plethora of sharply stained arborizations were revealed within a virtually unstained matrix (compare Figures 2D–2F to Figures 2A–2C). As expected from previous analysis in sections (Ott and Burrows 1998), the VACs contained the highest density of stained arborizations (Figures 1D, 2D, 2F, and 2G). All dark regions of blurred “background” in Figures 1D and Figures 2D–2F are exclusively due to arborizations that are not in focus (cf. Figures 2G, in which out-of-focus blur has been digitally reduced). Therefore, the abundance of selective staining itself was now the main factor limiting the visualization of detail in whole-mounts. The strong NADPHd in the perineurium which, after BF fixation, obscures the selective neuronal staining, was virtually eliminated (Figures 1E and 1F, asterisks). By contrast, the intensity of the selective neuronal NADPHd was markedly increased, confirming our recent findings in the locust brain (Ott and Elphick 2002). As in the brain, putatively NOS-unrelated NADPHd in glia was amplified by MF fixation but was blocked successfully by preincubation in acidic buffer (see Ott and Elphick 2002). This treatment also further reduced the neuronal background and resulted in preparations of exceptional clarity.
Clearing the Tissue for Whole-mount Viewing
NADPHd staining that extends into the depth of the tissue is of little use if the tissue cannot be cleared sufficiently for whole-mount viewing. Buffered glycerol induced heavy anisometric shrinkage and the ganglia collapsed dorsoventrally (this flattening is not very apparent from the ventral view in Figure 1A, but note the resultant folding of the surface in Figure 2B). This finding is in agreement with Bucher et al. (2000), who reported that glycerol produces extreme deformations, even when applied through a finely graded series of concentrations. Moreover, we found that MF-fixed tissue does not clear properly in glycerol. The traditional alternative to glycerol has been some variant of the procedure here referred to as ethanol/xylene clearing (e.g., Moroz et al. 1993; Elphick 1997; Sadamoto et al. 1998; Zayas et al. 2000). However, nitroblue formazan (NBF), the product of the NADPHd staining reaction, is soluble in ethanol and in organic clearing agents such as xylene and methyl salicylate. The solubility in ethanol necessitates rapid dehydration, which inevitably distorts the ganglion morphology (Bucher et al. 2000) but can still decolorize even strongly stained structures. Further fading occurs during the clearing and in the solvent of the mounting medium. Curiously, weak but widespread staining (e.g., background after BF fixation) was often less affected than sharp selective staining. This “low-pass filter effect” of ethanol/xylene clearing may be explained by steep NBF concentration gradients that arise at sharply stained structures and that drive the diffusion of NBF, whereas less diffusion may occur in larger, more homogeneously stained regions of tissue. Ethanol/xylene-cleared preparations deteriorated further on storage when NBF that had dissolved reprecipitated as the solvent evaporated from the mounting medium.
We therefore developed a new clearing technique (methanol/cedar oil clearing; Figures 1D and 2D–2G) on the basis of three observations. First, the blue form of NBF, which accounts for the selective staining, is very stable in methanol, while pink background NBF is extracted by it. Second, the swelling action of acetic acid counteracts the morphological distortions that occur during dehydration. Third, cedar oil makes an excellent clearing medium which, unlike xylene and methyl salicylate, does not dissolve the NBF. For dehydration, the ganglia were transferred from water directly into a single bath of methanol/acetic acid 3:1 for 15–30 min, followed again directly by three baths of 15–30 min in absolute methanol. This rapid and convenient procedure gave better preservation of morphology than dehydration through a finely graded methanol series (10% steps, 30 min each). After clearing in cedar oil, the preparations showed 30–40% isometric shrinkage (compared to unfixed ganglia in physiological saline) but virtually no deformation artifacts.
The combination of MF fixation, cold or acidic preincubation, and methanol/cedar oil clearing results in NADPHd whole-mount staining of unprecedented anatomic resolution. Particularly under differential interference contrast illumination, the preparations yield images of spectacular definition that could be mistaken for physical sections (Figure 2D–2F), even when focusing deep into the ganglion (Figure 2F). However, the greatest advantage lies in the rich 3D detail, which is only poorly conveyed by 2D images. The 3D anaglyph in Figure 2G has been created from 75 focal planes taken through an intact metathoracic ganglion. Although it must necessarily fall short of direct inspection through the microscope, it demonstrates the power of the techniques presented here. Moreover, it both complements and condenses into a single three-dimensional image the neuroanatomical analysis of Ott and Burrows (1998) in sectioned material.
The improvements in whole-mount NADPHd staining shown here are in part due to the use of MF in place of BF as a fixative, which improves whole-mount staining for two independent sets of reasons. First, MF fixation enhances the histochemical detection of putatively NOS-related NADPHd per se. Second, it renders the tissue much more permeable to the staining reagents. In a previous report (Ott and Elphick 2002), which was based exclusively on frozen sections, we have characterized MF as a fixative with regard to the specificity and sensitivity of NADPHd staining in the nervous system of two insects: the cockroach, a species in which NADPHd after BF fixation fails almost completely, and the locust, a species in which BF fixation has previously been used with considerably greater success. In both species, MF fixation resulted in much stronger and more selective staining than BF fixation, and also in lower background. This suggests that the NADPHd activity of NOS is strongly sensitive to formaldehyde-fixation in the cockroach and is also sensitive, to a lesser extent, in the locust, and that MF fixation improves the preservation of the NADPHd activity of NOS in both species. Moreover, the lower background suggests a more efficient suppression of other types of NADPHd activity. These advantages obviously apply to sections and whole-mounts alike, although whole-mounts benefit more than sections from the increased signal/background ratio because of their thickness. However, the poor penetration of the NADPHd staining reagents poses a problem not present in sectioned material. Here we have shown that, as a strong coagulant and a solvent of lipids, MF renders the tissue much more accessible for the staining reagents than BF. In whole-mounts, this increased penetrability of the fixed tissue is at least as important as the improvement in selectivity and sensitivity. This is evident from the observation that, after BF fixation, only superficial staining developed even after prolonged preincubation in the staining reagents (not shown).
The improved penetration of the staining reagents after MF fixation and cold preincubation, and the advantages of methanol/cedar oil clearing, do not depend on species- or tissue-specific enzyme properties and are hence universally applicable in vertebrate and invertebrate tissue. We have applied the method successfully to other parts of the locust nervous system, including abdominal ganglia and the brain, and to the nervous system of other invertebrates, including crayfish (Ott et al. 2001) and snail (unpublished data). Selective elimination of NOS-unrelated NADPHd by low pH, used in this study to suppress glial staining, has previously been reported in vertebrates (Spessert and Claassen 1998) and could therefore improve the NOS signal in a variety of species and tissues. However, the duration and pH of the treatment (if any) need to be determined empirically for the species and tissue at hand to avoid loss of NOS-related staining. Finally, MF fixation may improve the detection of NOS-related NADPHd in other species in which NOS is formaldehyde-sensitive (Ott and Burrows 1999; Ott et al. 2001; Ott and Elphick 2002). This may not necessarily be the case in vertebrates, in which the NADPHd activity of NOS is very robust and resists formaldehyde fixation. However, as in locusts, methanol and formaldehyde in combination could result in lower background, and it will be interesting to explore MF fixation in vertebrate tissues. We therefore propose that our new techniques may be very useful for revealing the three-dimensional architecture of NOS expression in other invertebrate and vertebrate preparations.
Footnotes
Acknowledgements
Supported by grant S11816 from the BBSRC to MRE.