
Abstract
Hyperhomocysteinemia, caused by a lack of cystathionine β synthase (CBS), leads to elevated plasma concentrations of homocysteine. This is a common risk factor for atherosclerosis, stroke, and possibly neurodegenerative diseases. However, the mechanisms that link hyperhomocysteinemia due to CBS deficiency to these diseases are still unknown. Early biochemical studies describe developmental and adult patterns of transsulfuration and CBS expression in a variety of species. However, there is incomplete knowledge about the regional and cellular expression pattern of CBS, notably in the brain. To complete the previous data, we used in situ hybridization and Northern blotting to characterize the spatial and temporal patterns of Cbs gene expression during mouse development. In the early stages of development, the Cbs gene was expressed only in the liver and in the skeletal, cardiac, and nervous systems. The expression declined in the nervous system in the late embryonic stages, whereas it increased in the brain after birth, peaking during cerebellar development. In the adult brain, expression was strongest in the Purkinje cell layer and in the hippocampus. Immunohistochemical analyses showed that the CBS protein was localized in most areas of the brain but predominantly in the cell bodies and neuronal processes of Purkinje cells and Ammon's horn neurons.
H
In humans, pathological concentrations of homocysteine are associated with hyperhomocysteinemia, an autosomal recessive disease. In hyperhomocysteinemic patients, several factors lead to pathological concentrations of homocysteine: mutations in the CBS gene, mutations in the methylenetetrahydrofolate reductase (MTHFR) gene that encodes an enzyme of the remethylation pathway, and deficiencies in vitamins B6, B12, and folate. The major clinical features of hyperhomocysteinemic patients are premature atherosclerosis and thromboembolism, lens dislocation, osteoporosis, major skeletal development abnormalities, and mental retardation with brain damage and a predisposition to schizophrenia and epilepsy (Harker et al. 1974; Grieco 1977; Vollenweider et al. 1990; Mudd 1985; Van den Berg et al. 1995). The biological factors that link hyperhomocysteinemia due to CBS deficiency to these developmental abnormalities are unknown.
A murine model of hyperhomocysteinemia has been generated by Cbs gene targeting (Watanabe et al. 1995). Both homozygous CBS-deficient mice [CBS(−/−)] and heterozygous CBS-deficient mice [CBS (+/−)], used as models of hyperhomocysteinemia, exhibit some pathological features of hyperhomocysteinemia, such as vascular defects (Dayal et al. 2001; Eberhardt et al. 2000; Lentz et al. 2000; Weiss et al. 2002) and major skeletal development abnormalities (Watanabe et al. 1995; and Janel et al. unpublished data).
Although central nervous system CBS expression is well documented, the available data are largely based on early biochemical studies by investigators who used enzyme and cell fractionation methods to describe developmental and adult patterns of transsulfuration and CBS expression in liver and brain in a variety of species (Gaitonde and Richter 1957; Hope 1959; Mudd et al. 1965; Sturman et al. 1970,1976; Volpe and Laster 1972; Rassin and Gaull 1975). Despite these previous publications, there is incomplete knowledge about the regional and cellular expression pattern of CBS, notably in the brain.
To complete the previous data, we used in situ hybridization (ISH) and Northern blotting to characterize the spatial and temporal patterns of Cbs gene expression during mouse development. To provide additional information on the expression of Cbs during the late developmental stages until adulthood, we characterized the expression pattern in the nervous system. We also characterized the expression pattern in the skeletal system, which has been shown to be affected in a murine model of hyperhomocysteinemia (Watanabe et al. 1995).
In the early stages of development, Cbs is ubiquitously expressed. At later stages, expression becomes more specific in the tissues that are mainly affected in hyperhomocysteinemia patients [liver, skeletal system, cardiac and nervous systems (Quere et al. 1999)] as well as those affected in the CBS(−/−) murine model [liver, skeletal system (Watanabe et al. 1995; and Janel et al. unpublished data)]. For example, in the skeletal system, Cbs mRNA was detected at early stages in chondroblasts in areas at which bone ossification is believed to be initiated. This high level of labeling persisted during later stages of development. It was still visible in the ribs at P0 and in the vertebrae at P7. In the nervous system, the expression pattern was uniform and very weak in the brain at P0. The expression level increased during later stages of brain development before becoming restricted to the Purkinje cell layer and the hippocampus in adult mice. We also used immunohistochemistry (IHC) to determine the location of the CBS protein in the adult brain. The protein was predominantly observed in the Purkinje cells and the Ammon's horn neurons, either in the cell bodies or in the neuronal processes.
Materials and Methods
Animals
All procedures were carried out in accordance with internal guidelines for animal handling. Mice were housed in a controlled environment with access to unlimited food and water. We made every effort possible to minimize suffering and the number of mice used. Heterozygous CBS(+/−) mutant mice were generously donated by Maeda Nobuyo (Department of Pathology, University of North Carolina, Chapel Hill, NC). The day of conception was designated as embryonic day (E) and the day of birth was designated as postnatal day (P). Animals were considered adult when they were older than 3 months. The presence of a vaginal plug on the next morning was defined as 0.5 day postconception. Pregnant mice were killed on days 9.5, 10.5, 12.5, 13.5, or 15.5 postconception. The legends are in accordance with the Atlas of Mouse Development (Kaufman 1995) and the Atlas of Histology with Functional Correlations (Eroschenko 1996).
In Situ Hybridization
ISH of Cbs mRNA in Mouse Sections. [35S]-UTP-labeled single-stranded RNA was synthesized from the NotI fragment of pBluescript, which contains the rat Cbs cDNA fragment (nucleotides 652–945), using T3 RNA polymerase as an antisense riboprobe, and from the EcoRI fragment of pBluescript using T7 RNA polymerase as a sense riboprobe. A conventional protocol was used for ISH (Peuchmaur et al. 1990). Briefly, embryos (E12.5, E15.5) and brains from FVB/N mice (P0, P2, P7, and 3 months) cryosections (8 μm) were transferred onto SuperFrost/Plus slides, postfixed in 4% paraformaldehyde/PBS for 10 min, and washed in diethylpyrocarbonate (DEPC)-treated double-distilled water. To reduce background, slides were acetylated in triethanolamine-containing acetic anhydride, rinsed in PBS, dehydrated in 95% ethanol, and air-dried. Rat CBS sense and antisense probes were used at a concentration of 2 × 106 cpm/slices in the hybridization mixture [50% deionized formamide, 20% (w/v) dextran sulfate, 1 × Denhardt's solution, and 500 μg/ ml tRNA in 0.3 M NaCl; 20 mM Tris-HCl, pH 7.5, 5 mM EDTA, pH 8]. This solution was mixed and heated for 2 min at 80C before being spotted on slides. Hybridization was carried out overnight at 8C and was followed by several stringent posthybridization washes (the most stringent wash was in 2 × SSC/formamide 50%, DTT 1 mM) at 68C for 20 min). Hybridization signals were visualized by exposure to Kodak NTB2 photographic emulsion for 1 month at 4C. Tissues were then counterstained with hematoxylin-eosin, dehydrated through graded concentrations of alcohol, cleared in xylene, and coverslipped.
ISH of Cbs mRNA in Whole-mount Embryos. Briefly, single-stranded RNA probes containing digoxigenin were synthesized from previously described linearized template exactly as directed by the manufacturer (Roche Diagnostics; Maylan, France). E9.5, 10.5, 12.5, and 13.5 FVB/N embryos were fixed for 3 hr in 4% paraformaldehyde. The 12.5 and 13.5 FVB/N embryos were then cut in half mid-sagittally using a mouse brain matrix (Fracalosso, France) and stored immediately at −20C in 100% methanol (Vialard et al. 2000). Whole-mount hybridization was carried out as described by Conlon and Rossant (1992). The embryos were prepared as described (Conlon and Rossant 1992) and prehybridized for 1 hr at 68C in hybridization buffer (50% formamide, 0.75 M NaCl, 10 mM Pipes, pH 6.8, 1 mM EDTA, 100 μg/ml tRNA, 0.05% heparin, 0.1% BSA, 1% SDS). The hybridization buffer was replaced, single-stranded RNA probes labeled with digoxigenin were added to 2 μg/ml, and the embryos were hybridized overnight at 68C. The embryos were washed in stringent posthybridization solutions and detection of hybridization was performed as described by Conlon and Rossant (1992).
Northern Blotting Analyses
Liver and brain tissues were obtained from 3-month-old homozygous (−/−) and heterozygous (+/−) CBS-deficient mice and from wild-type (+/+) control mice from the same litter and were stored at −80C. Total RNA was prepared from two mixtures of three different tissues by the guanidinium thiocyanate procedure (Chomczynski and Sacchi 1987). The quantity and purity of the RNA were assessed by measuring absorbance at 260 nm and 280 nm. Samples of denatured RNA (15 μg) were separated by 2.2 M formaldehyde/1% agarose gel electrophoresis and transferred to nylon membranes. The RNAs were crosslinked to the membrane using a Stratalinker UV crosslinker before hybridization. Part of the rat Cbs cDNA fragment (nucleotides 652–945) was labeled using the Multiprime DNA labeling system (Amersham; Piscataway, NJ) with [α-32P]-dCTP (NEN Life Science Products; Boston, MA). We ensured that loading was equal and that the RNA from the liver samples had not been degraded by hybridization with a probe specific for mouse Cu/Zn superoxide dismutase 1 (SOD1). This gene was expressed at equal levels in the three groups of mice. We ensured that the RNA from the brain samples had not been degraded by hybridization with a probe specific for mouse cytochrome C oxidase subunit I (Cyt C Ox). Hybridization was performed at 68C overnight. The blot was then washed and exposed to X-ray film at −80C.
Western Blotting Analyses
The brains from 3-month-old homozygous (−/−) and heterozygous (+/−) CBS-deficient mice and from wild-type (+/+) control mice were quickly removed and dropped in Laemmli solubilizing buffer (50 mM Tris-HCl, pH 6.8, 100 mM DTT, 2% SDS, 10% glycerol) and boiled for 10 min. Then 60 μg of each protein sample was subjected to SDS electrophoresis on 5–12.5% gels and transfered to nitrocellulose membranes (Immuno-blot PVDF Membrane; Bio-Rad Laboratories, Hercules, CA). The membrane was incubated in a blocking buffer consisting of 5% (w/v) nonfat dried milk in Tris-buffered saline (1.5 mM Tris base, pH 8, 5 mM NaCl, 0.1% Tween-20) for 1 hr at room temperature (RT). The membranes were then incubated in the presence of the anti-CBS antibody (Omura et al. 1984) (1:10,000 dilution) or the anti-calbindin antibody (Chemicon; Temecula, CA) (1: 2000) in 5% BSA Tris-buffered saline at 4C overnight. After washes, membranes were incubated with a 1:2000 dilution of anti-rabbit Ig peroxidase-conjugated antibody (Cell Signaling Technology; New England Biolabs, Beverly, MA) in blocking buffer for 1 hr. Bound antibody was visualized with the ECL chemiluminescence system (Amersham Pharmacia Western Blotting Detect System).
Immunohistochemistry
Brains were removed from the mice and sagittal cryosections (8 μm) were fixed in 4% paraformaldehyde for 10 min. Brain cryosections from both homozygous (−/−) CBS-deficient mice and wild-type (+/+) mice from the same litter were loaded on each slide. Because a second unspecific band was present on the Western blot, the anti-CBS antibody (Omura et al. 1984) was preincubated with a protein preparation (3 μg/μl) from the brains of CBS(−/−) mice [20 μl per μl of anti-CBS antibody solution in 10 ml of Tris-buffered saline (TBS: 0.1 M Tris-HCl, pH 7.4, 0.15 M NaCl)] for 12 hr at 4C to mask nonspecific antigens.
After several washes in TBS containing 0.1% Triton X-100, the sections were treated with freshly made 1% H2O2 for 30 min. The nonspecific binding sites were blocked with 3% BSA in TBS/0.1% Triton X-100 for 1 hr. The sections were then incubated with the anti-CBS antibody (1:1000 dilution) at 4C overnight. Biotinylated goat anti-mouse IgG (Amersham; 1:400 dilution) used as secondary antibody was incubated for 1 hr at RT and detected with streptavidin-peroxidase complex. The immunoreaction was visualized by treating the sections with DAB+ substrate chromatogen reagent (DAKO; Glostrup, Denmark) for 2 min at RT. Tissues were then counterstained with hematoxylin, dehydrated through graded concentrations of alcohol, cleared in xylene, and coverslipped.
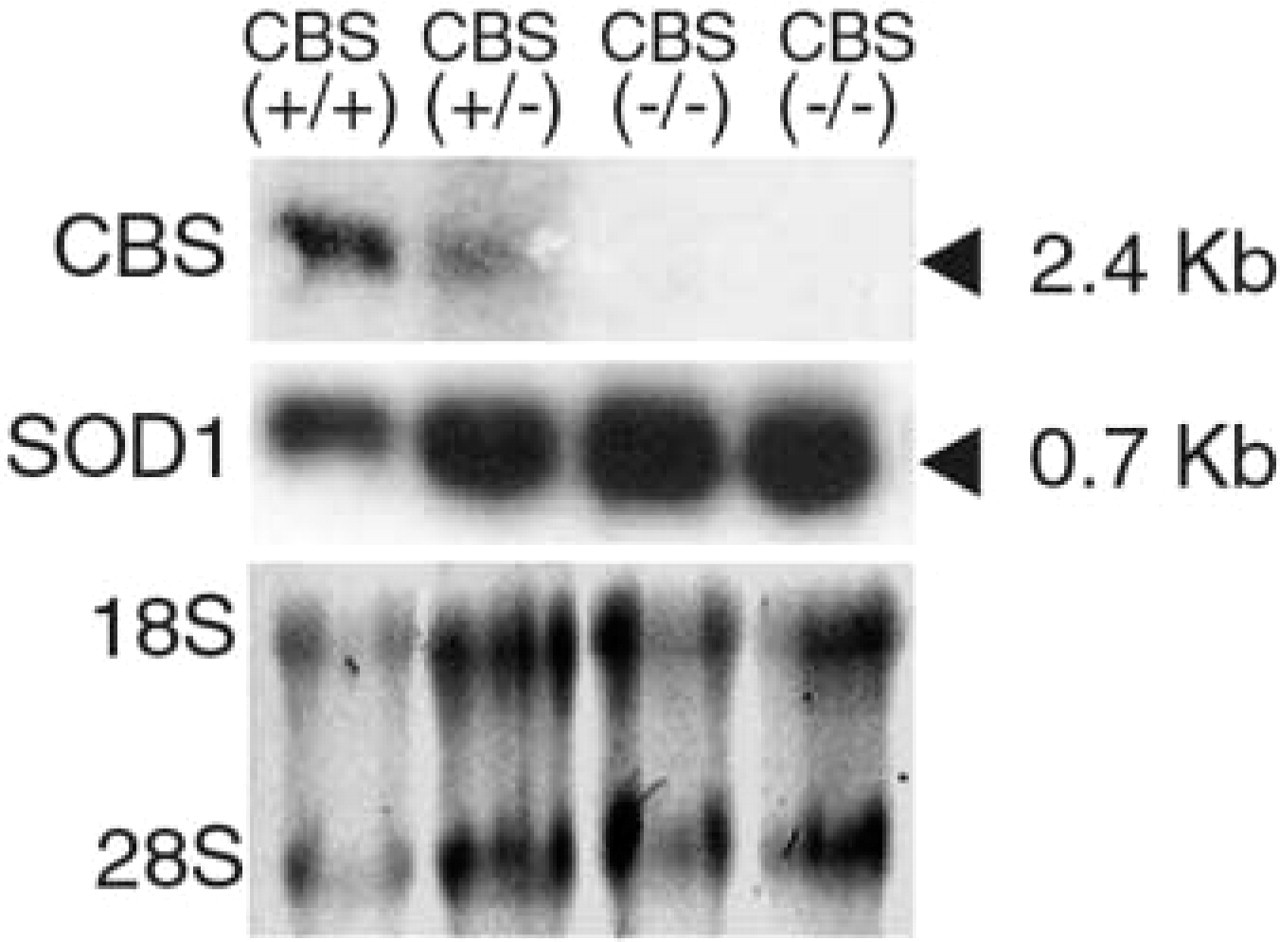
Expression of the Cbs gene in liver tissues from homozygous (−/−) CBS-deficient mice, heterozygous (+/−) CBS-deficient mice and from wild-type (+/+) control mice from the same litter. (
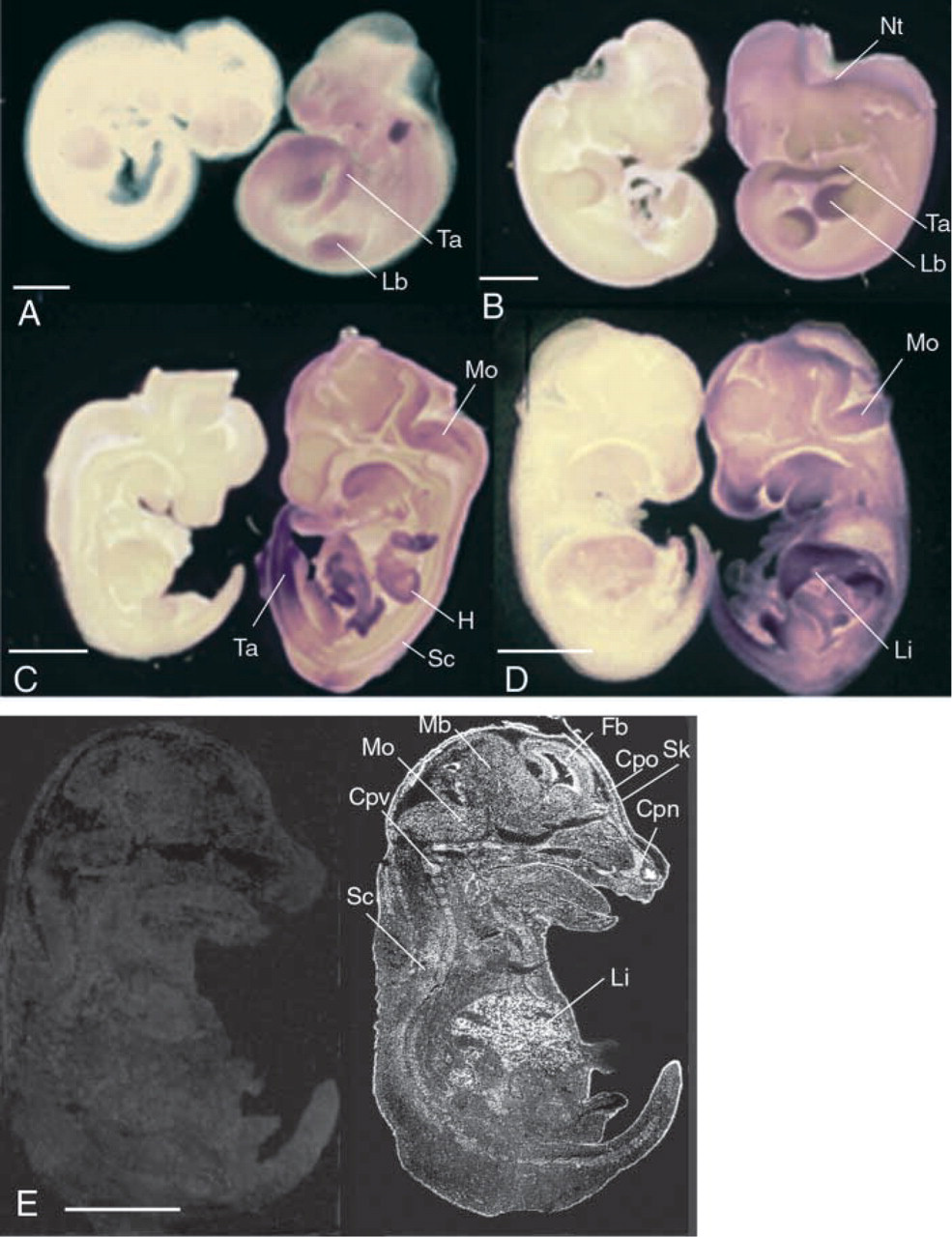
Expression pattern of Cbs in E 9.5–E13.5 FVB/N mouse fetuses. The rat Cbs sense riboprobe was used as a negative control (
Results
ISH Analyses
A 350-bp cDNA fragment containing exons 5–8 of the Cbs gene was isolated from the rat Cbs orthologue (Swaroop et al. 1992). This fragment shares 95% sequence identity with the recently identified mouse cDNA counterpart (Genbank accession number BC013480). We confirmed the specificity of this cDNA fragment by Northern blotting analysis (Figure 1) using mRNA isolated from the livers of CBS(−/−) mice, CBS(+/−) mice, and wild-type (+/+) control mice from the same litter. A single 2.4-kb band was observed in the CBS(+/+) and CBS(+/−) mice but not in the CBS(−/−) mice. The riboprobe synthesized from this fragment was then used to investigate the expression of the Cbs gene during development from 9.5 days after conception (E9.5) until adulthood (3 months).
Spatial Distribution of the Cbs mRNA During Development
Whole-mount hybridization revealed that Cbs was ubiquitously expressed between E9.5 and E10.5. However, the strongest expression was observed in limb buds, tail, and brain vesicles (Figures 2A and 2B). In Figure 2A the purple spot at the base of the neck was an artifact. Between E12.5 and E13.5, Cbs was expressed in developing organs, including the liver, cartilage primordium, cardiac system, and nervous system (Figures 2C and 2D). At these stages, Cbs transcripts were still detected in embryonic limb buds and in the tail. To determine the cellular expression pattern, we performed ISH analyses on embryo sections (Figure 2E). In the digestive system at E12.5, high levels of Cbs mRNA were first detected in certain developing mesoderm regions, including the liver (Figure 2E). Closer analysis showed that primitive hepatocytes were labeled at this stage. In later embryonic stages and in the adult mouse, strong expression was observed in hepatocytes (data not shown). In the cardiovascular system, expression was observed from E12.5 (Figure 2C). At this stage, Cbs transcripts were detected in endocardial cells (data not shown). Cbs mRNA was also detected in the aorta at later stages of development (data not shown).
In the skeletal system, Cbs mRNA was also detected in somitic derivatives and in tissues originating from the mesoblast at E12.5. The developing vertebral bodies and arches were labeled, whereas the developing intervertebral disks were not (Figures 3.1A and 3.1B). The original core of each disk, which is composed of cells originating from the notochord, was not labeled. Cbs mRNA was also detected in chondroblasts of the developing bones of the head, pharyngolaryngeal cartilage, ribs, vertebrae, and in areas at which bone ossification is believed to be initiated (Figure 2E). This dense labeling persisted during later stages of development in the perichondrium and the chondroblasts of ribs at P0 (Figure 3.1C) and vertebrae at P7 (Figure 3.1D). In adult mice (data not shown), the signal persists in the perichondrium in cartilage and in periosteum in bone.
Spatial distribution of Cbs mRNA in the Mouse Nervous System
The Cbs gene was expressed in the CNS from an early stage. At E9.5, the entire neural tube was labeled (Figure 2A), with the strongest labeling in the neural tube closure region (Figure 3.2A). This expression changed during neural tube formation and appeared to be more caudal at E10.5 (Figure 3.2B). At E10.5, the telencephalon was strongly labeled.
Between E12.5 and E15.5, strong signal was observed in most proliferating zones of the nervous system. The strongest labeling was observed in the ventricular zone of many structures, such as the neopallial cortex, the striatum and the fourth ventricle, and the medulla oblongata. Weak labeling was observed in the neopallial cortex, the midbrain, the medulla oblongata and the spinal cord (Figures 2E, 4A, and 4B). Although the strong expression in the ventricular zone of brain persisted at E15.5, expression in the nervous system declined in intensity.
From P0, the expression was uniform and very weak (Figure 4C). At P2, the expression became stronger and more limited to specific regions of the brain, such as the cerebellum and the olfactory bulb (Figure 4D). This expression changed during the later stages of cerebellar development (Figure 4E). We also used Northern blotting to study the expression of Cbs in the nervous system between P0 and adulthood. Northern blotting was performed with mRNA extracted from the brain and cerebellum of P0, P2, P7, P10, and P15 mice and of 3-month-old CBS(+/+) mice (Figure 5). With both brain and cerebellar extracts, a single 2.4-kb band was observed. This band was weak in P0 mice. The intensity then increased from P2 to P10, and decreased from P15 to 3 months. In the adult mice, weak signal was present in the whole brain, whereas strong signal appeared to be restricted to the Purkinje cell layer and to the hippocampus, including Ammon's horn and the dentate gyrus (Figure 4F).
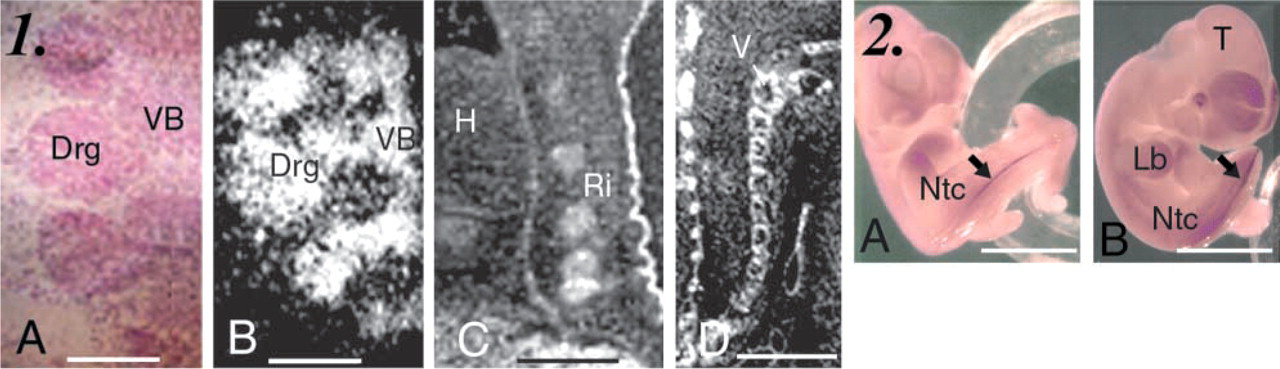
Distribution of Cbs mRNA in developing skeletal and neural system in FVB/N embryos and mice. (
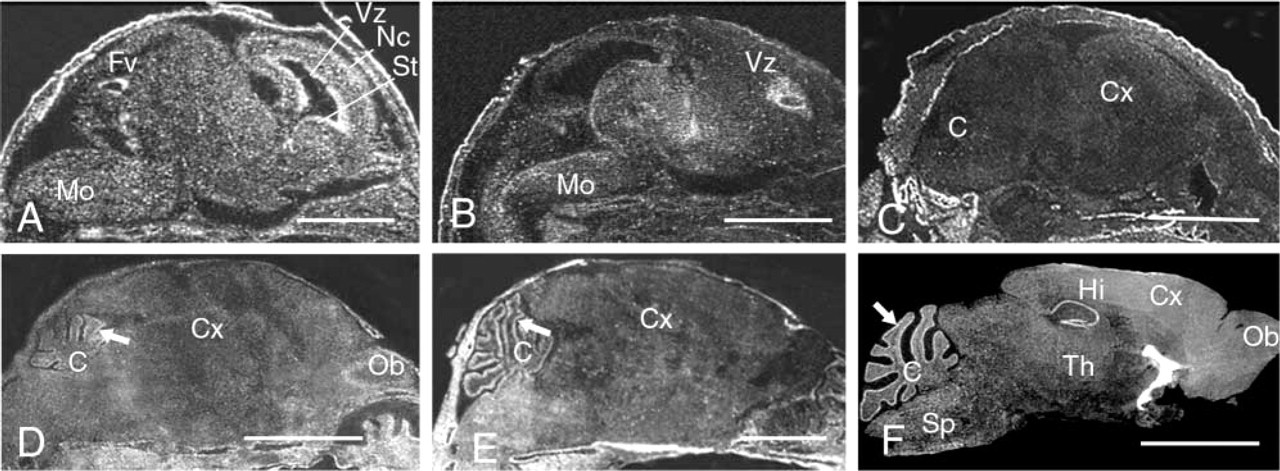
Expression of Cbs in the mouse central nervous system. (
Thus, during the earlier stages of brain development, Cbs is highly expressed in neuroblast cells. In the later stages of development (from birth to adulthood), the Cbs expression pattern changes as cells differentiate. This suggests that CBS is involved in neuronal development and function in some neurons.
Spatial Localization of CBS Protein in Mouse Nervous System
Because Cbs expression might be specific to some neurons, we used IHC methods to localize the protein in the adult brain. We carried out Western blotting experiments using an anti-rat CBS antibody (Omura et al. 1984) to probe protein samples isolated from the brains of CBS(−/−) mice, CBS(+/−) mice, and wild-type (+/+) control mice from the same litter. Two bands were observed, one at 68 kD and one at 50 kD (Figure 6A). The specific 68-kD band was strong in CBS(+/+) mice, weak in CBS(+/−) mice, and absent in CBS(−/−) mutant mice. The intensity of the unspecific 50-kD band was the same in the wild-type and mutant mice. To obtain specific labeling, the polyclonal mix was incubated with CBS(−/−) brain proteins for 12 hr before immunoreaction. All immuno-reactions were performed on two different brain sections from 3-month-old CBS(−/−) and CBS(+/+) mice from the same slide and were repeated at least four times with three or four different samples.
The protein profile closely mirrored the RNA profile. A weak signal was present in the whole brain. Despite the high level of general background staining in the section, we observed uniform and weak expression in the neurons of the cerebral cortex, the striatum, the thalamus, and the spinal cord, whereas no significant expression was detected in the non-neuronal cells of these brain regions. The strongest expression was observed in the hippocampus and the cerebellum (Figure 6B). In the cerebellum, signal was observed in the cell bodies and neuronal processes of the Purkinje cells (Figures 6C–6E). Weak signal was observed in cells of the granular cell layer. In the hippocampus, strong signals were detected in the cellular bodies and the neuronal processes of the CA2 and CA3 neurons (Figures 6F and 6H) and weaker signals were observed in the other regions, such as the CA1 neurons (Figure 6F and 6G) and the dentate gyrus.
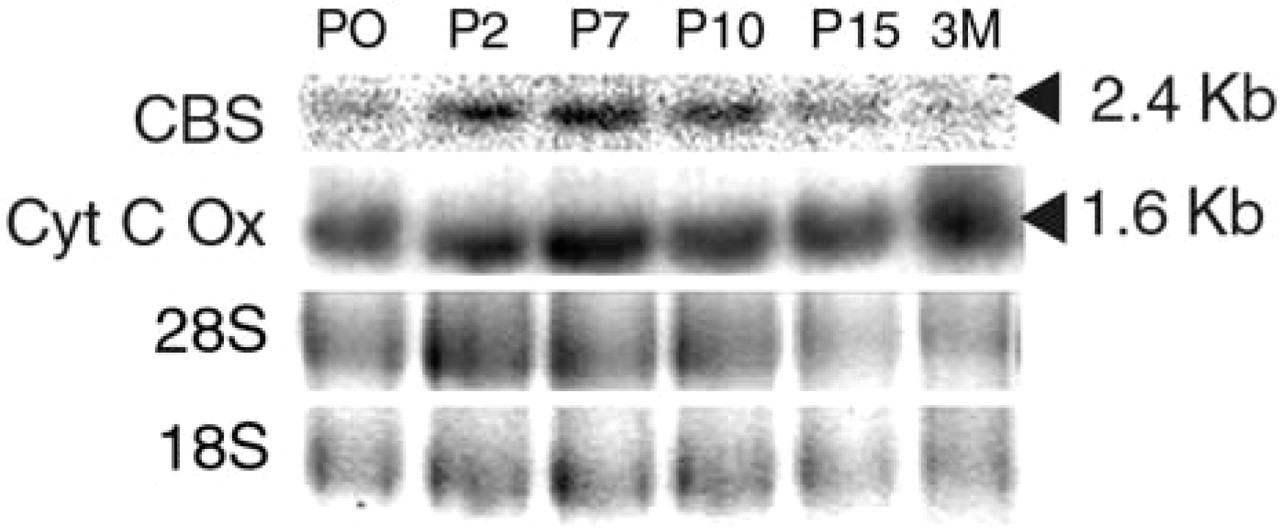
Expression of Cbs in the mouse CNS during late development. (
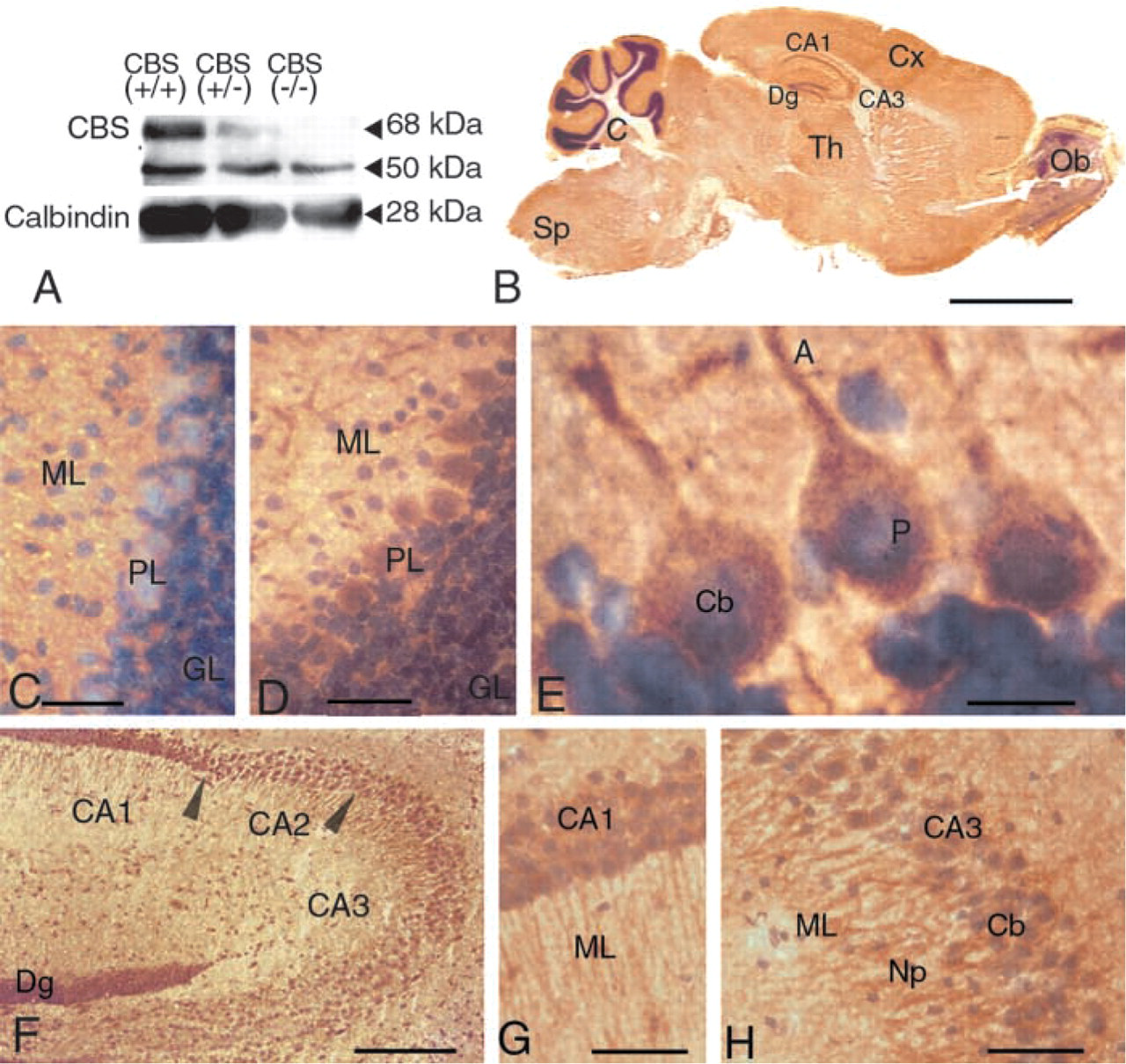
CBS immunostaining in the mouse CNS. (
Discussion
We characterized the expression profile of the mouse Cbs gene and demonstrated that its spatial and temporal expression patterns are developmentally regulated. In the early stages of development, Cbs was ubiquitously expressed. It then predominantly accumulated in specific tissues, especially those that are affected in homocysteinemic patients (liver, cardiac, skeletal and nervous systems) and a murine model of hyperhomocysteinemia (Watanabe et al. 1995; and Janel et al. unpublished data). These results are consistent with the previously published results obtained on early human embryos (3–6 weeks post conception) (Quere et al. 1999), which suggest that CBS expression during mammalian development is functionally conserved. The demonstration that the expression of Cbs in the murine nervous system is developmentally regulated provides further information about the expression of Cbs. From E9.5 to E12.5, strong signal was observed in many zones of the nervous system, including the ventricular zone, the neopallial cortex, the medulla oblongata, the midbrain, and the spinal cord (Figure 2). At later stages of brain development, the expression declined in the nervous system (Figure 4). After P0, the expression changed as the cerebellum developed (Figures 4D–4F). In the adult CNS, the signal was restricted to the Purkinje cell layer and the hippocampus.
We also showed that the CBS protein is localized in the brain in adult mice. A low level of expression was observed in the whole brain cerebral cortex, the striatum, the thalamus, and the spinal cord, whereas the protein was strongly expressed in the cell bodies and the neuronal processes of the Purkinje cells and hippocampal neurons. This pattern is consistent with the pattern of CBS activity in nine regions of the adult rat brain (Kohl and Quay 1979).
The expression of Cbs therefore appears to be developmentally regulated. A high level of expression was observed in neuroblasts during the early stages of development and in some differentiated neurons in the adult CNS, suggesting that CBS is involved in the growth and maturation of neural networks. This regulated pattern during CNS development should be related to a regulated rate of homocysteine and methionine. In addition, it might explain why high homocysteine levels result in developmental defects of the neural tube in most animal models (Rosenquist and Finnell 2001). In the adult brain, Cbs is highly expressed in some neurons, such as Purkinje cell neurons and hippocampal neurons, and more particularly in the cell bodies and the neuronal processes of these cells.
Homocysteine, the concentration of which is regulated by CBS activity, is known to be excitotoxic in vitro, and there is evidence for synthesis of homocysteine and its derivatives, such as homocysteic acid, in the brain (Gaitonde and Richter 1957). In spite of many studies performed in vitro (Sawada et al. 1982; Ito et al. 1991), there is scant evidence that homocysteine is actually involved in neurotransmission or modulation of neuronal activity in vivo. In fact, the observation of developmentally regulated and specifically localized CBS expression suggests a role for homocysteine metabolism in normal brain function and particularly a putative role of homocysteine in synaptic transmission, which has been observed in vitro (Yuzaki et al. 1996). It is not known whether homocysteine is cytotoxic. However, high concentrations of homocysteine and its derivatives, which act in vitro as natural transmitters at the N-methyl-
It has been also suggested that high homocysteine concentrations promote hypersensitivity to excitotoxicity and apoptosis in many cultured neuronal cells, such as rat embryonic neuronal cells, human neuronal cell lines, rat cerebrocortical cells, rat cerebellar granule cells, and rat hippocampal neurons (Kim and Pae 1996; Lipton et al. 1997; Parsons et al. 1998; D'Emilia and Lipton 1999; Kruman et al. 2000). We demonstrated that many of the neural cells that are sensitive to high concentrations of homocysteine strongly express Cbs. Therefore, CBS might protect the neurons by reducing the endogenous concentrations of homocysteine. These changes in the homocysteine concentration might explain the correlation between high homocysteine concentrations and some neurodegenerative diseases.
It is not clear whether the endogenous concentration of homocysteine is involved in neuronal function or development, but the putative role of homocysteine in synaptic transmission might explain the importance of the regulation of homocysteine metabolism in the CNS through CBS activity. In addition, the observed association of elevated plasma homocysteine with some neurodegenerative diseases makes it more pressing to better understand the role of homocysteine metabolism in normal brain.
Footnotes
Acknowledgements
Supported by a grant from the Fondation Jérôme Lejeune and by an EU grant (BMH4-CT98–3039). KR was supported by the Fondation Jérôme Lejeune. ET was supported by the Fondation pour la Recherche Médicale (FRM).
We thank Dr M. Sakaguchi (Department of Molecular Biology, Graduate School of Medical Science, Kyushu University, Japan) for providing the anti-CBS antibody and Dr M. Nobuyo (Department of Pathology, University of North Carolina, Chapel Hill, NC) for providing heterozygous CBS(+/−) mice. We also thank Nicole Créau and Carmela Lopes for helpful advice and discussions.