
Abstract
Here we report a detailed analysis of the expression and localization of the giant protein AHNAK in adult mouse tissues. We show that AHNAK is widely expressed in muscle cells, including cardiomyocytes, smooth muscle cells, skeletal muscle, myoepithelium, and myofibroblasts. AHNAK is also specifically expressed in epithelial cells of most lining epithelium, but is absent in epithelium with more specialized secretory or absorptive functions. In all adult tissues, the main localization of AHNAK is at the plasma membrane. A role for AHNAK in the specific organization and the structural support of the plasma membrane common to muscle and lining epithelium is discussed.
T
Materials and Methods
Antibodies and Controls
Rabbit polyclonal antibodies were made against AHNAK using two different epitopes. The first, KIS (CKISMPDVDL HLKGPK), was chosen according to the strategy of Shtivelman and Bishop (1993). The second epitope, CQL (CQLPE VELSVSTKKE), is localized in the C-terminus of the protein (aa 5630–5643). The KIS-AHNAK and CQL-AHNAK antibodies used throughout our study are affinity-purified. Antibodies were purified from immununized rabbit sera by affinity chromatography using KIS or CQL peptide crosslinked to Sepharose beads. The purity of the immunoglobulins was controled by SDS-PAGE in the absence and in the presence of a reducing agent. The purified antibodies were stored in PBS and 50% glycerol at a concentration of 0.15 mg/ml. The specificity of the antibodies was tested in Western blotting on HeLa cell extracts and on the recombinant AHNAK protein. To control for antibody specificity, the KIS-AHNAK antibody and the CQL-AHNAK antibody were incubated with the corresponding KIS or CQL peptides before primary immunoreaction. No immunoreaction was detected using this method. For immunochemistry, the specificity of immunostaining was controlled by preincubation of the affinity-purified KIS-AHNAK antibody with the KIS peptide. For double immunofluorescence experiments, we used a monoclonal anti-PECAM, MEC13.3, from Sigma (St Louis, MO). Cyan3 and Alexa 488 antibodies used as second antibody were provided by Molecular Probes (Eugene, OR).
Immunoblotting
In adult mouse, specimens from kidney, heart, thymus, esophagus, pancreas, liver, and lung were removed. These specimens were cut into small pieces and quickly frozen with liquid nitrogen until used. Specimens were homogenized with a glass homogenizer and were then heated at 90C for 3 min in denaturation buffer composed of 2% SDS, 25 mM Tris-HCl, pH 6.8, 10% glycerol, 0.1% bromophenol blue, 1m M EDTA. Proteins were electrophoretically resolved by one-dimensional SDS-PAGE (6% polyacrylamide), the gel was then equilibrated in transfer buffer containing 0.03% SDS and proteins were transferred onto nitrocellulose membrane. The membrane was first incubated with CQL-AHNAK antibody (1:100 dilution) in TBS containing 0.3% Tween-20 (TTBS) at 4C for 16 hr. After washing, the membrane was incubated with a 1:10,000 dilution of horseradish peroxidase secondary antibody and the immune complex was revealed by chemiluminescence. Nitrocellulose membrane was then stripped in TTBS with 1% SDS and 0.1% sodium azide, and then reprobed with the KIS-AHNAK antibody (1:1000 dilution).
Immunochemistry
Tissues were screened for AHNAK localization using a tissue array produced by ResGen Co. (Invitrogen; Carlsbad, CA). We used paraffin-embedded tissue spots representing several mouse organs: adrenal, brain, heart, large intestine, small intestine, renal cortex, renal medulla, liver, lung, lymph node, skeletal muscle, pancreas, salivary gland, skin, spleen, stomach, thymus, prostate, testis, ovary, uterus, breast, and esophagus. Paraffin was removed and immunohistochemistry was then performed using the DAKO ABC horseradish peroxidase system (DAKO; Carpinteria, CA). After endogenous peroxidase inactivation with H2O2 3% and sodium azide 0.05%, followed by extensive washing, tissues were blocked in 5% normal goat serum in TBS for 30 min and incubated with the KIS-AHNAK antibody at 1:100 dilution in TBS plus 2% normal goat serum overnight at 4C. After rinsing with TBS, tissues were incubated with a biotinylated goat anti-rabbit Ig (DAKO) diluted 1:100 in TBS plus 5% normal goat serum for 30 min at room temperature. After extensive washing with TBS, the slide was incubated with the horseradish peroxidase complex according to the manufacturer′s recommendations. The chromogen used was AEC+ (DAKO). Slides were counterstained with hematoxylin. For indirect immunofluorescence localization of AHNAK, mouse testis, brain, and liver were frozen in isopentane at −80C and cut in 10-μm sections. Slides were fixed with 4% paraformaldehyde for 30 min and washed abundantly before permeabilization for 5 min with 0.5% Triton X-100 in TBS. After extensive washing and blocking with goat serum, primary antibodies were incubated overnight at 4C at the following dilutions: KIS-AHNAK 1:100; PECAM 1:5. Slides were then incubated with secondary antibody for 1 hr and nuclear countstaining was performed with Hoechst 33258 (1 μg/ml). Double immunofluorescence analysis was performed using confocal microscopy (Leica TCS-SP2) or fluorescence microscopy (Zeiss Axiovert 200M). Images were directly captured, saved, and transferred to Adobe Photoshop 5.5.
Results
Immunoblotting Analysis and Tissue Distribution of AHNAK
Extracts from various mouse tissues were analyzed for AHNAK expression by immunoblotting using affinity-purified antibodies raised against the CQL and the KIS peptides (Figure 1). To normalize for total protein content of the tissue preparations, the extracts were also analyzed for α-tubulin content (Figure 1A). Both the CQL and the KIS antibody reacted strongly with a major protein band that migrated with the expected molecular weight for AHNAK, confirming the specificity of our antibodies (Figure 1B). A high level of expression of AHNAK was observed in heart, lung and esophagus, whereas kidney, pancreas, and liver expressed only moderate levels of AHNAK. AHNAK immunoreactivity was absent in the thymus. CQL and KIS antibodies also reacted with lower molecular weight protein bands. These protein bands are present only in tissue extracts that express the full-length AHNAK protein and may therefore represent proteolytic products of AHNAK. In support of that assumption, we found that AHNAK is highly sensitive to metal-dependent proteases. When tissues are lysed in the absence of the metal chelators EDTA and EGTA, the full-length AHNAK immunoreactivity decreases, with a concomitant increase in the lower molecular weight protein bands recognized by both the AHNAK KIS and CQL antibodies (data not shown). A band of approximately 100 kD was also detected with the CQL antibody but not with the KIS antibody. This band might correspond to a C-terminal AHNAK proteolytic fragment. The affinity-purified KIS antibody was next used for immunoperoxidase and indirect immunofluorescence analysis of AHNAK immunoreactivity in mouse tissues (Figures 2–5).
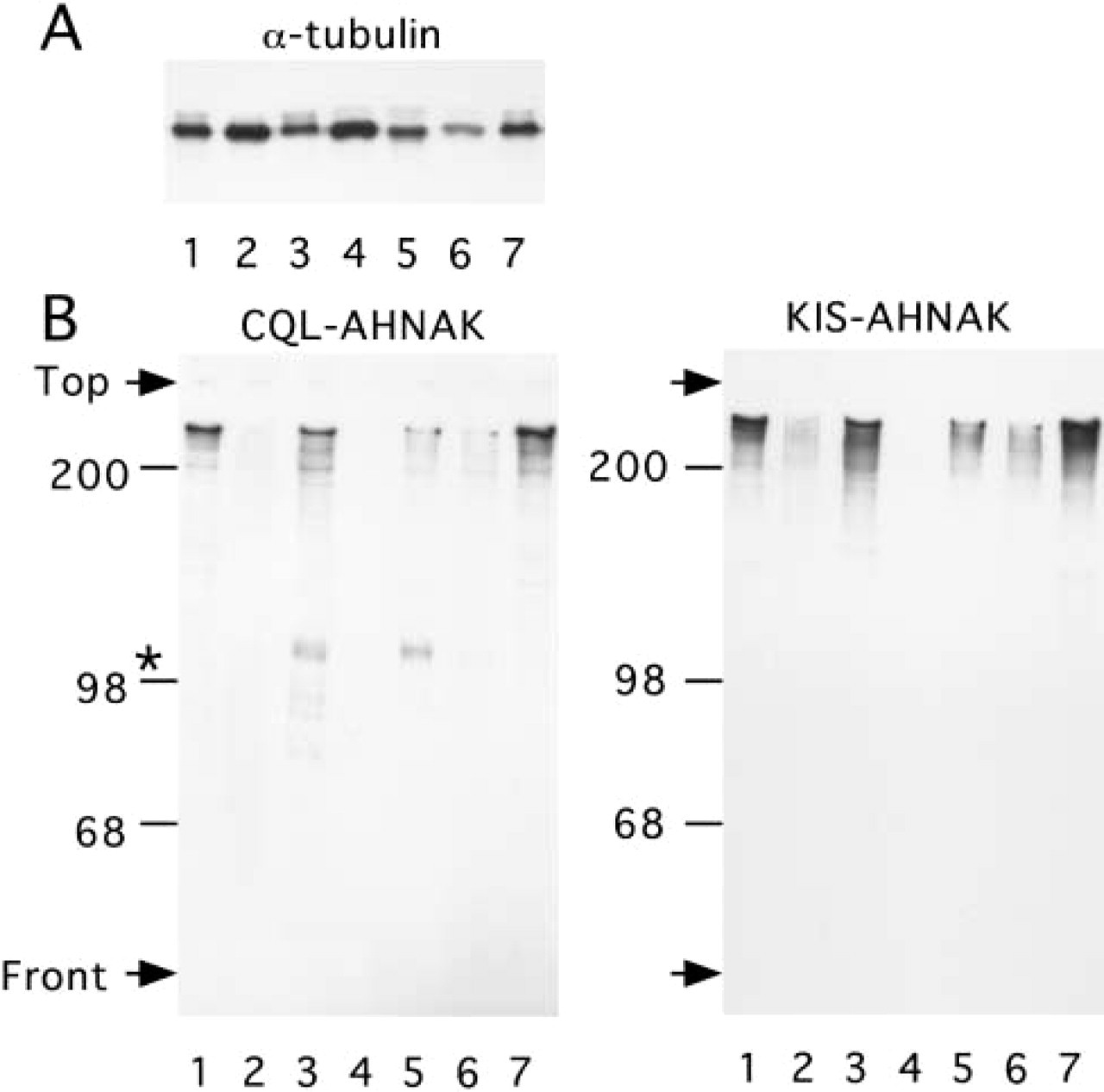
Immunoblotting analysis of AHNAK content in mouse organs or tissues. Tissue extracts were analyzed by Western blotting using α-tubulin (
AHNAK Localization in Muscle Cells
Immunohistochemical analysis of AHNAK immunoreactivity in different tissues revealed a specific localization of AHNAK in muscle cells (Figure 2). As previously reported by Haase et al. (1999), heart sections clearly show a plasma membrane localization of AHNAK in cardiomyocytes (Figure 2A). Figure 2B shows a muscle section incubated with affinity-purified KIS antibody that was preincubated with the KIS peptide and counterstained with biotinylated horseradish peroxidase. In this control, AHNAK staining was completely abolished, confirming our biochemical analysis that the KIS antibody used in our study is specific for AHNAK protein. AHNAK expression is not restricted to cardiomyocytes, but appears to be a general feature of the different muscular cell types. As observed in cardiomyocytes, in the skeletal myocytes forming the striated skeletal muscle fiber, AHNAK immunoreactivity also accumulated at the plasma membrane (Figure 2C). In the small intestine, AHNAK antibodies decorated the layers of smooth muscle cells of the tunica underlining the epithelium (Figure 2D). Both the longitudinal and the circular layer of smooth muscle cells showed strong AHNAK immunoreactivity. A cross-section of the circular layer clearly showed that AHNAK is localized at the plasma membrane. This pattern of AHNAK immunoreactivity in smooth muscle cells was also observed on the muscularis propria of the body of the stomach, which also consists of an inner circular and outer longitudinal layer (data not shown). Other structures composed of smooth muscle cells found in various tissue also express AHNAK (Figure 2E and 2F). In the ovary, theca cells showed strong plasma membrane AHNAK immunoreactivity (Figure 2E). Theca cells, which are important for steroid secretion, also have muscle characteristics (Motta and Familiari 1981; Self et al. 1988). At ovulation, these cells by their contraction may be capable of squeezing the follicular wall or may favor follicle collapse and subsequent involution during atresia. Smooth muscle cells that compose the tunica media and ensheathe large blood vessels in various tissue were also stained with AHNAK antibodies. In testis, smooth muscle cells surrounding small and large arteries were AHNAK-positive (Figure 2F). AHNAK was also expressed by PECAM-positive endothelial cells, as revealed by yellow pixels in merged confocal double-labeled fluorescent image (Figure 2F). We also observed apparently higher AHNAK immunoreactivity in endothelial cells of small arterioles than in large arteries. Slender smooth muscle-like cells surrounding seminiferous tubules also expressed AHNAK (Figure 2F). These cells are responsible for the production of collagen and elastin fibers present in the lamina propria, while their contractile activity helps the movement of spermatogonia along the tubule.
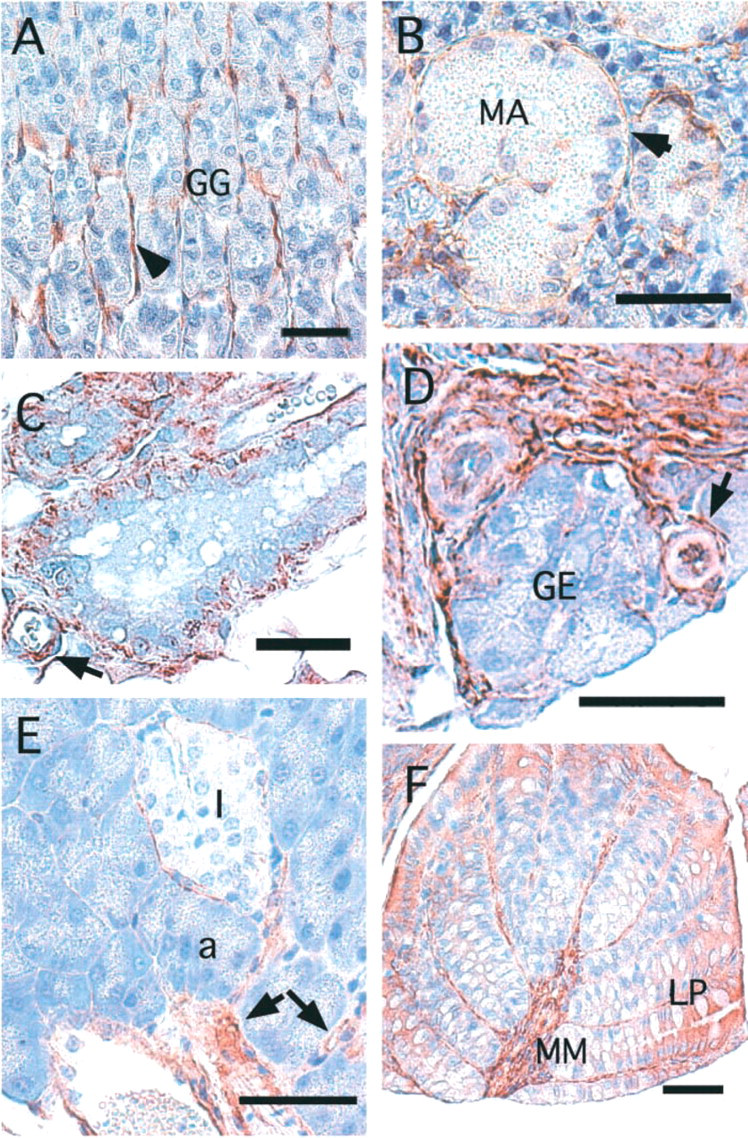
AHNAK Localization in Lining Epithelium
In the stratified epithelium of the skin, AHNAK was expressed at the plasma membrane of keratinocytes of the stratum basale and the stratum spinosum (Figure 3A). AHNAK staining was not observed in the cornified layer. AHNAK is also expressed in cells of papillary dermis and in some cells of the hair follicle (Figure 3A).
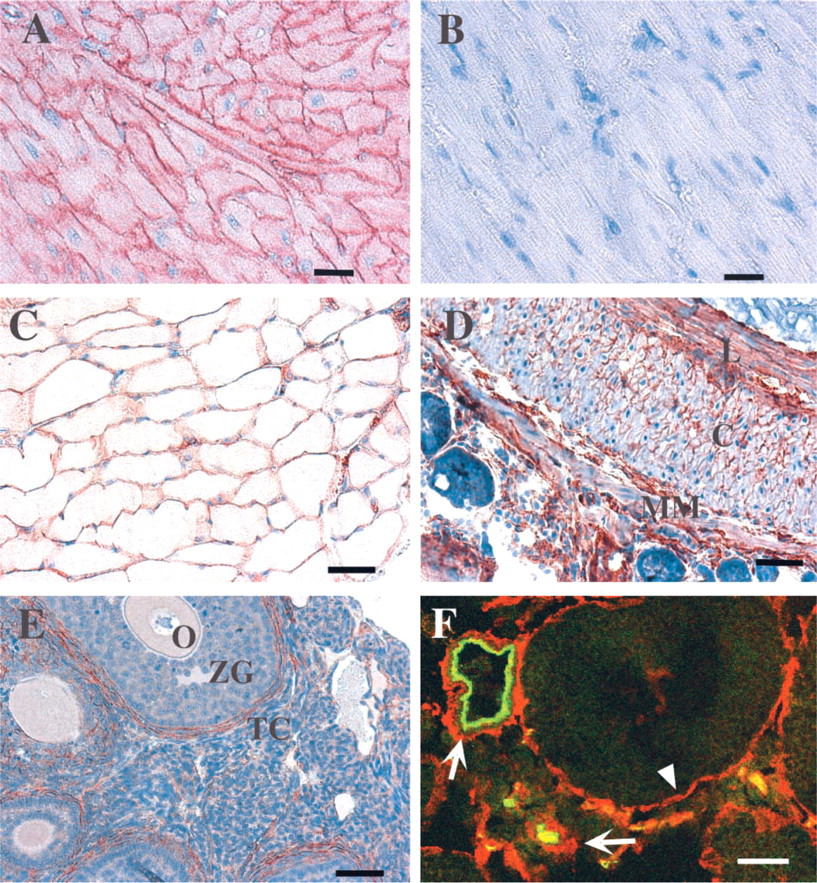
AHNAK localization in muscle tissue. (
In the upper part of the digestive tract, i.e., the esophagus, AHNAK was found in the stratified squamous epithelium, which in rodents may be keratinized (Figure 3B). Here again, AHNAK immunoreactivity was concentrated mainly at the plasma membrane. As shown in Figure 3B, AHNAK was also present in the muscle cells of the striated muscularis layer. In kidney, AHNAK immunochemistry showed that AHNAK was localized in a special type of epithelium of the urinary system that lines the collecting ducts, ducts of Bellini, and the pelvicalyceal system (Figure 3D; and data not shown). This transitional epithelium, called urothelium, is present along the conducting passages of the urinary system and protects the organism from acid and hypertonic urine. In liver, AHNAK immunoreactivity was confined to the portal tract. Figure 3E focuses on a typical portal tract containing three main structures. The largest is a terminal branch of the hepatic portal vein (PV), which has a thin wall lined by endothelial cells labeled with PECAM antibody. These endothelial cells were AHNAK-positive. Smaller-diameter vessels are arterioles, which show lower AHNAK immunostaining. Finally, the bile collecting duct, lined by simple columnar epithelium, was strongly immunostained with AHNAK antibody. In these lining epithelial cells, AHNAK was concentrated at the plasma membrane. There was no AHNAK immunostaining of the liver hepatocytes. Immunohistochemistry with the lung (Figure 3E) confirmed the Western blotting analysis, showing that a high level of AHNAK protein is present in lung (Figure 1, Lane 3). AHNAK was highly expressed in parenchymal cells (epithelium of alveoli) and in the smooth muscle layer surrounding the bronchiolar epithelium. Faint AHNAK immunoreactivity was also present in pseudostratified bronchial epithelium. The labeled epithelial cells were mainly ciliated cells characterized by large nuclei. Their soma was weakly immunostained but concentrated immunoreaction deposit was observed at the apical plasma membrane and in contact with the basement membrane (white arrow).
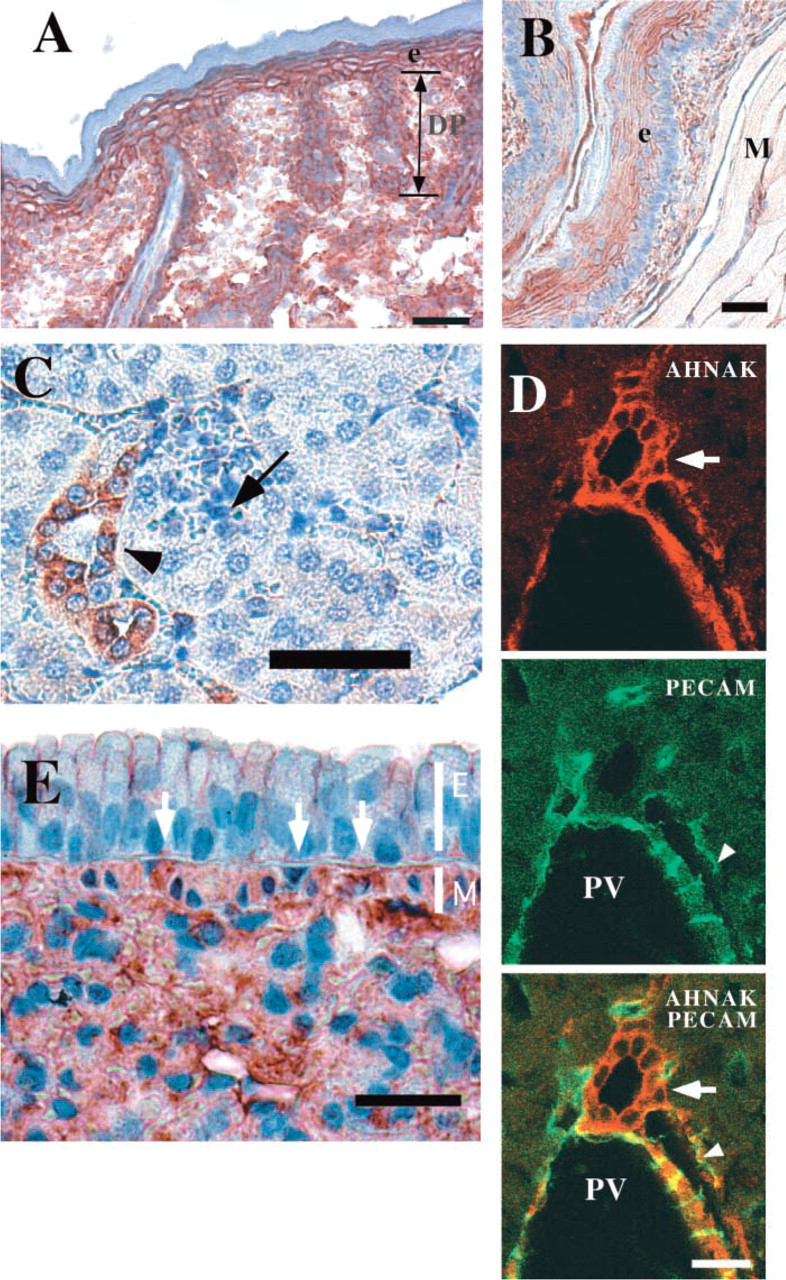
AHNAK localization in lining epithelium. (
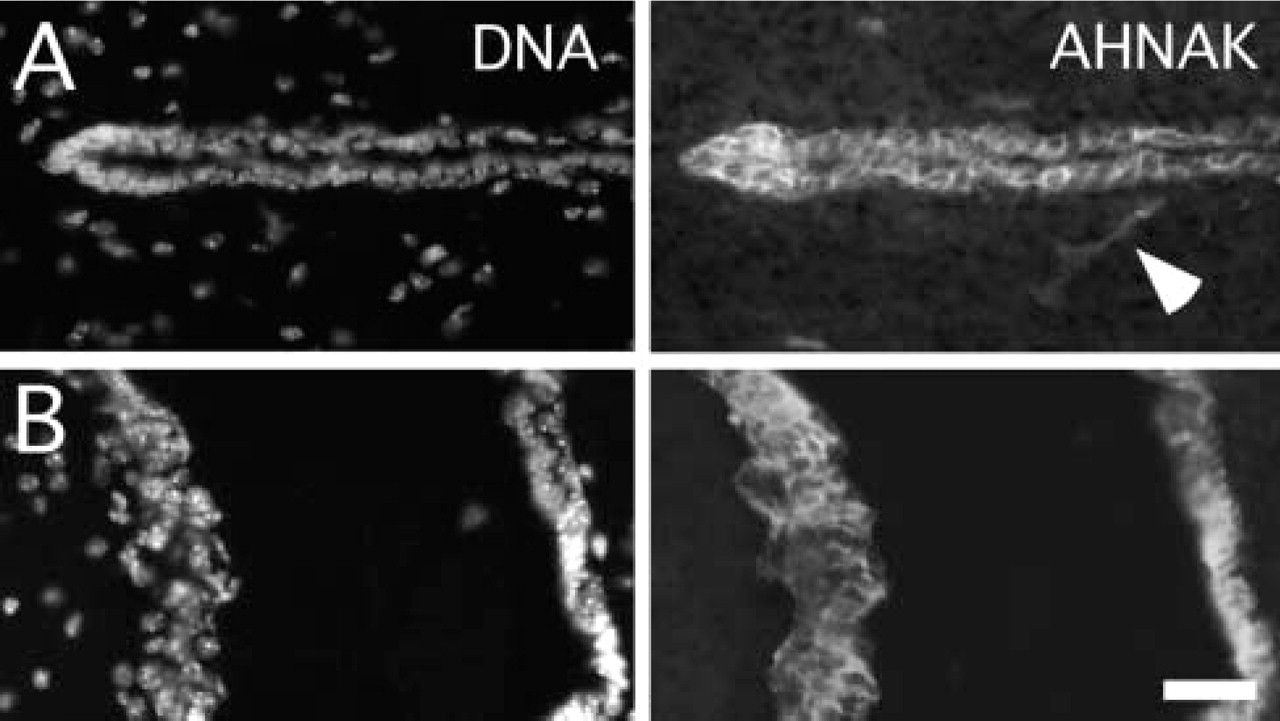
AHNAK is localized in the ventricular wall of adult brain. Double immunofluorescence analysis of AHNAK and HOECHST (DNA) labeling in the third ventricle (
We also found AHNAK immunoreactivity associated with the plasma membrane of several other lining epithelia, such as the stratified epithelium of the tunica albuginea of the testis (data not shown), and also in the epithelial cells lining the ventricles of the central nervous system (Figure 4). In the adult mouse brain, AHNAK immunoreactivity was abundant in epithelial cells that bordered all the ventricles, including the lateral ventricles (not shown), the third ventricle (Figure 4A), and the fourth ventricle (Figure 4B). There was no apparent AHNAK immunostaining of neuronal and glial cell soma. Brain capillary endothelial cells were also immunostained (see Figure 4A).
In contrast to muscle cells, not all epithelial cells synthesize AHNAK. As shown in Figure 5A, the different secretory epithelial cells that constitute the stomach mucosa, including mucus-secreting cells, acid-secreting cells, and pepsin-secreting cells, were not AHNAK-immunoreactive. AHNAK staining appeared as a thin strand between the gastric gland, which corresponds to the muscularis mucosae that extends from the base of a gastric gland, to the lumen, whose contraction expels gastric secretion into the stomach lumen. The absence of AHNAK in secretory epithelium was confirmed in the salivary gland (Figure 5B), the mammary gland (Figure 5C), and the prostate gland (Figure 5D). In the salivary gland, the epithelium of serous acini, with nuclei flattened against the basement membrane, and the epithelium of mucous acini were not AHNAK-immunoreactive. The thin AHNAK immunoreactivity surrounding serous acini corresponds to the processes of contractile myoepithelial cells. In mammary glands, myoepithelial cells surrounding the gland and endothelial cells of capillaries were also labeled by the AHNAK antibody (Figure 5C). The prostate glandular epithelium was not positive for AHNAK, whereas the supporting tissue, which contains numerous smooth muscle cells and capillaries, was labeled by the AHNAK antibody (Figure 5D). Figure 5E shows the immunolabeling of AHNAK in the pancreas, a large gland composed of glandular epithelium with exocrine and endocrine functions. The endocrine tissue formed by the islets of Langerhans was not stained, nor were the glandular acini, which constitute the exocrine part of the pancreas. Only the endothelial cells of the capillaries were immunolabeled. AHNAK immunostaining was also not observed in epithelium with absorptive functions, such as the epithelium of the small intestine (data not shown; see also Figure 2D) and the absorptive epithelium lining the colon, which also has protective functions (Figure 5F). The mucosa of the colon, formed of straight tubular glands consisting of cells specialized for water absorption and of mucus-secreting globlet cells, was devoid of AHNAK immunoreactivity. However, significant cytoplasmic immunoreactivity was present in the cells of the luminal portion of the epithelium. These cells, which are derived from precursor cells present in the basal portion of the epithelium, are more mature and are subjected to the stretching forces of the luminal content. The muscularis mucosae, which extends into the lamina propria between the folds of the gland, also expressed AHNAK.
Discussion
Table 1 summarizes the tissue distribution and the cellular localization of AHNAK in mouse tissues. The protein AHNAK is a hallmark of muscular and muscle-like cells with contractile properties organized in contractile tissue or scattered in the tissue as fibromyocytes or myoepithelial cells. AHNAK is also specifically expressed in epithelial cells of the skin, the esophagus, the tunica albugina of the testis, the bile ducts, and the brain ventricles. In contrast to muscle cells, not all epithelial cells express AHNAK. We did not find significant AHNAK immunoreactivity in the glandular epithelium of the mammary gland, the salivary gland, the stomach, the prostate, or the exocrine and endocrine pancreas, all of which have secretory functions. We were also unable to detect AHNAK in the absorptive epithelium of the proximal part of the nephron and of the intestine. In addition to muscle cells and lining epithelial cells, AHNAK is also present in endothelial cells of several capillary and blood vessels (see Figures 2F, 3D, 4A, and 5C–5E). However, in fenestrated endothelium of the kidney glomeruli (Figure 3C), the hepatic sinusoid (data not shown), and the continuous capillaries of lung (data not shown), AHNAK immunoreactivity was below the detection limit. This suggests that the level of AHNAK expression in endothelial cells might depend on the blood vessel properties.
AHNAK localization is restricted to lining epithelium. (
Tissue and cell localization of AHNAK
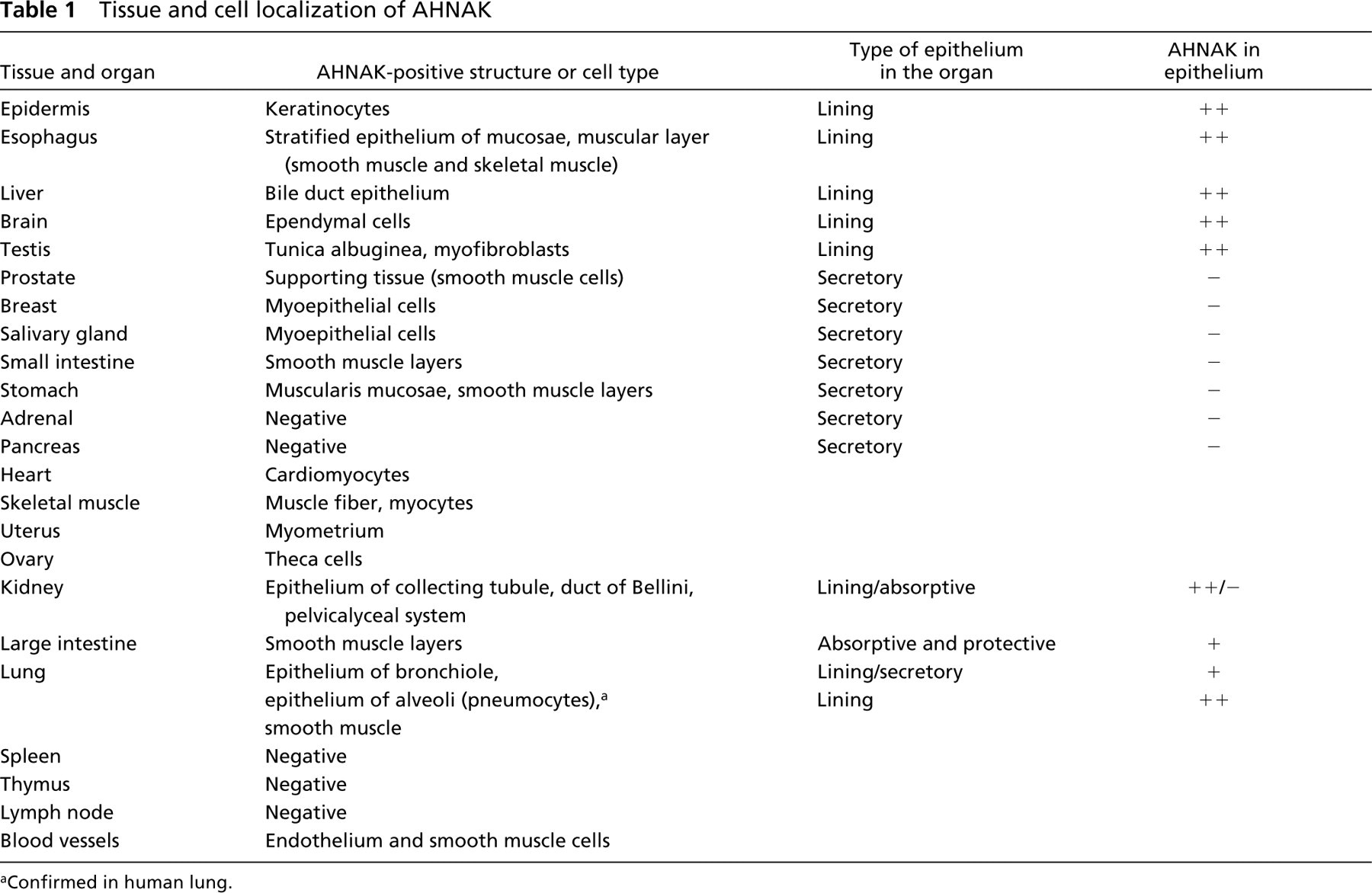
aConfirmed in human lung.
In adult tissue, the main subcellular localization of AHNAK is at the plasma membrane. Cytoplasmic staining was also observed in fibroblast or certain epithelial cells, such as those in the colon (Figure 5F). Exclusive membrane location of AHNAK has also been recently reported in rat cardiomyocytes by Hohaus et al. (2002), using two region-specific AHNAK antibodies raised against either amino-terminal or carboxyl-terminal epitopes. These findings contrast with the predominant nuclear localization of AHNAK observed in cultured cells lines of nonepithelial origin (Shtivelman and Bishop 1993). However, recent data reported that in MDCK cells derived from canine kidney epithelium, membrane targeting of AHNAK is regulated by cell density and the formation of cell-cell contacts (Sussman et al. 2001). The authors proposed that the plasma membrane-anchored AHNAK may be involved in the growth arrest of normal epithelial cells (Sussman et al. 2001). In adult mouse tissues, AH-NAK-expressing cells are terminally differentiated cells and therefore do not proliferate. This status might account for the predominant membrane localization of AHNAK in most adult tissues. Alternatively, recent studies indicate that AHNAK may be proteolytically cleaved in vivo (Skoldberg et al. 2002) and that AHNAK fragments may have functional roles intracellularly (Sekiya et al. 1999). It is therefore possible that, in cell culture, cytoplasmic AHNAK is more susceptible to proteolysis, and that AHNAK proteolysis could contribute to the apparent nuclear localization of the protein. In normal keratinocytes, endogenous full-length AHNAK accumulates at the cell membrane, but the transfected C-terminus of AHNAK accumulates within the cell nuclei (Nie et al. 2000).
The predominant cellular expression of AHNAK in muscles and in lining epithelium that we report in this study might help to elucidate the membrane function of this giant protein. First, it allows restriction of the search for AHNAK function to essentially two specialized cell types. Moreover, several structural and functional features are shared between muscle and lining epithelium. These tissues are subjected to stretching force and have elastic properties. For example, the epithelium of the esophagus is subjected to the stretching force of the bolus, and muscle cells to stretching forces during the contraction-relaxation cycles. This implies that common cellular organizations are involved in both tissues to respond and control the stretching forces. The first is the cell-cell interactions necessary for maintaining tissue integrity. Several molecular structures are known to participate in cell-cell interactions, such as desmosomes, tight junctions, and adherens junctions. Although initially AHNAK was co-purified with the desmosomal component during cell fractionation of keratinocytes (Hieda and Tsukita 1989), and therefore was been named desmoyokin, later studies clearly demonstrated that AHNAK (desmoyokin) is not directly associated with desmosomes (Masunaga et al. 1995). We have investigated the possible association of AHNAK with tight junctions in endothelial cells of brain capillary and ependymal cells (data not shown). Double immunofluorescence labeling and confocal microscopy analysis for AHNAK and the tight junction-associated protein ZO1 revealed a distinct localization of the two proteins (unpublished data). Finally, the observation that AHNAK can migrate from the cytoplasm to the plasma membrane in cell culture without any effect on cell-cell adhesion (Hashimoto et al. 1995) does not support a direct role for AHNAK in cell-cell interactions. We envision that AHNAK may be more likely associated with the structural support of the membrane during changes in membrane shape, the reinforcement of the cortical framework, and signaling pathways. It is significant that AHNAK shares structural similarity and subcellular localization with two other giant proteins abundantly expressed in muscle cells, titin and dystrophin. Both of these giant proteins lie close beneath the plasma membrane (Shimada et al. 1993; Mussini et al. 1995). Dystrophin is part of a multimeric protein complex that links actin on the inside of the cells to extracellular matrix protein through transmembrane proteins (Samitt and Bonilla 1990; Wakayama et al. 1993; Norwood et al. 2000). Titin is a giant muscle protein. Its size and location within the sarcomere structure suggests that it plays an important role in the mechanism of muscle elasticity (Freiburg et al. 2000). It will be very interesting to investigate if AHNAK may also act as a scaffold protein, recruiting and maintaining the organization of cytoskeletal proteins at the plasma membrane. This will obviously require the identification and characterization of AHNAK-associ-ated proteins at the plasma membrane. Two membrane-associated signaling pathways regulated by AHNAK have already been characterized. In the cardiomyocyte, AHNAK modulates membrane-bound L-type calcium channel activity (Haase et al. 1999). Intriguingly, organization of the cortical actin network has been implicated in modulation of voltage-gated ion channel activity (Maguire et al. 1998), and in vitro interaction between the carboxyl-terminal region of AHNAK and actin has recently been described (Hohaus et al. 2002). Peptides from the central domain of AHNAK also bind and activate PLCγ in the presence of arachidonic acid (Sekiya et al. 1999). The activation of this enzyme mediates several cellular responses, including cytoskeletal rearrangements that lead to protection of the plasma membrane in response to mechanical stress (Ryan et al. 2000; Ruwhof et al. 2001). Finally, taking into account that AHNAK is an abundant muscular protein with possible important functions associated with structural support of the plasma membrane, it will be now of primary interest to investigate if its function is altered in muscular dystrophies that are characterized by cell membrane defects.
Footnotes
Acknowledgements
Supported in part by a grant from Association pour la Recherche contre le Cancer (Christian Delphin) and a Ligue Nationale contre le Cancer fellowship (Benoit Gentil).
We thank Dr Peoc'h for valuable advice and Dr La Marre Jonathan for critical reading of the manuscript.