
Abstract
Tyro 3 family receptors contain three members—Tyro 3, Axl, and Mer—that are essential regulators of mammalian spermatogenesis. However, their exact expression patterns in testis are unclear. In this study, we examined the localizations of Tyro 3, Axl, Mer, and their ligand Gas6 in postnatal mouse testes by immunohistochemistry. All three members and their ligand were continuously expressed in different testicular cells during postnatal development. Tyro 3 was expressed only in Sertoli cells with a varied distribution during testis development. At day 3 postnatal, Tyro 3 was distributed in overall cytoplasmic membrane and cytoplasm of Sertoli cells. From day 14 to day 35 postnatal, Tyro 3 appeared on Sertoli cell processes toward the adlumenal compartment of seminiferous tubules. A stage-dependent Tyro 3 immunoexpression in Sertoli cells was shown by adulthood testis at day 56 postnatal with higher expression at stages I-VII and lower level at stages IX-XII. Axl showed a similar expression pattern to Tyro 3, except for some immunopositive Leydig cells detected in mature testis. In contrast, immunostaining of Mer was detected mainly in primitive spermatogonia and Leydig cells, whereas a relative weak signal was found in Sertoli cells. Gas6 was strongly expressed in Leydig cells, and a relative weak staining signal was seen in primitive spermatogonia and Sertoli cells. These immunoexpression patterns of Tyro 3 family receptors and ligand in testis provide a basis to further study their functions and mechanisms in regulating mammalian spermatogenesis.
R
Tyro 3 family receptors are widely expressed in adult tissues such as neural, lymphoid, vascular, smooth muscular, reproductive tissue, and retina, and in primary and tumor cell lines derived from these sources (Graham et al. 1995; Crosier et al. 1997; Prieto et al. 2000; Konishi et al. 2004; Valverde et al. 2004). It has been shown (Lu et al. 1999; Lu and Lemke 2001) that triple mutant (Tyro3−/-Axl−/-Mer−/-) mice displayed multiple major organ defects, neurological abnormalities, severe lymphoproliferative disorders, spermatogenesis failure, and physiological deficits, suggesting that Tyro 3 family play significant biological roles in multiple tissues. How they perform the function is unknown.
More attention deserves to be paid to the functions of Tyro 3 family receptors on mammalian spermatogenesis. The triple knockout male mice completely lose the production of mature sperm, whereas single- and double-gene mutant male animals were fertile (Lu et al. 1999). These findings suggest that these three receptors are essential regulators of spermatogenesis and that their functions in gonadal development are compensated by each other. However, the expression patterns of Tyro 3 family receptor and their ligand Gas6 have not been extensively studied. Although the mRNAs of Tyro 3, Axl, Mer, and Gas6 in testicular cells have been examined by in situ hybridization and Northern blot in a few studies (Lu et al. 1999; Chan et al. 2000; Wong and Lee 2002), the results have not been consistent. A study on the immunoexpression of Tyro 3 family receptors and Gas6 in their protein levels would be helpful to clear up this matter. Furthermore, a precise localization and expression kinetics of Tyro 3, Axl, Mer, and Gas6 proteins in testis would be a basis to further study their functions in spermatogenesis.
In this study, immunohistochemistry staining was used to examine the immunoexpression patterns of receptor Tyro 3, Axl, Mer, and ligand Gas6 in postnatal developing testis.
Materials and Methods
Animals
All C57BL/6 mice used in this study were obtained from the Laboratorial Animal Center of Beijing University (Beijing, China) and handled in compliance with the guideline for the care and use of laboratory animals established by the Chinese Council on Animal Care.
Antibodies
Goat anti-Tyro 3, Axl, and Mer polyclonal antibodies and rabbit anti-Gas6 polyclonal antibody were used for detecting protein Tyro 3, Axl, Mer, and Gas 6. All primary antibodies and reagents related to immunohistochemistry staining and Western blotting were obtained from Santa Cruz Biotechnology (Santa Cruz, CA), except where noted otherwise.
Western Blotting
Testis from d35 mouse was lysed on ice with lysis buffer (25 mM Tris-HCl, pH 8.0, 150 mM NaCl, 1 mM EDTA, 0.1% SDS, 0.1% sodium deoxycholate, 1% Triton X-100, protease inhibitors: 1 mU/ml aprotinin, 0.1 mM leupeptin, 0.5 mM PMSF) for 1 hr. Insoluble materials were removed by centrifugation at 12,000 × g at 4C for 15 min; supernatant was mixed with loading buffer and boiled for 5 min. Then the samples were loaded on 10% SDS-PAGE and subsequently electrotransferred onto poly(vinylidene difluoride) transfer membranes (Millipore; Bedford, MA). After blocking with 2% BSA in PBS containing 0.1% Tween-20 (PBS-T) for 1 hr, the electrotransferred membranes were incubated with anti-Tyro 3, anti-Axl, anti-Mer, or anti-Gas6 antibody at 1:1000 dilution at 4C overnight. After washing with PBS-T, the membrane was incubated with horseradish peroxidase-conjugated rabbit anti-goat IgG (for Tyro 3, Axl, and Mer) or goat anti-rabbit IgG (for Gas6) antibody (Zhongshan; Beijing, China) at room temperature for 1 hr. After washing with PBS-T, antigen-antibody complex was visualized by using an enhanced chemiluminescence detection kit (Zhongshan).
Tissue Section Preparations
Postnatal day 3 (d3), d7, d14, d21, d35, and d56 normal mice were anesthetized with CO2 and then killed by cervical dislocation. The testes were immediately removed and fixed with 4% paraformaldehyde in 0.1 M PBS, pH 7.4, for 12 hr at room temperature. The samples then were dehydrated in ethanol, cleared with xylene, embedded in paraffin, sectioned at 5 μm thickness in a microtome, mounted onto poly-L-lysine precoated glass slides, and dried overnight at 37C for immunohistochemistry staining.
Immunohistochemistry Staining
Immunohistochemistry staining was conducted by the avidin-biotin-peroxidase complex method. Samples were deparaffinized in xylene at room temperature and rehydrated with a graded ethanol and then with distilled water. Next, they were incubated in PBS containing 3% H2O2 for 15 min to inactive endogenous peroxidase activity. After a brief wash in PBS, they were soaked in a citrate buffer and microwaved at 100C for 10 min for antigen retrieval. Before addition of the primary antibodies, the samples were incubated with normal rabbit serum for Tyro 3, Axl, and Mer or normal goat serum for Gas6 to block nonspecific binding. They were then incubated overnight with the primary antibody; goat anti-Tyro 3, Axl, and Mer antibodies; and rabbit anti-Gas6 antibody in a moist chamber at 4C. This was followed by three 5-min washes in PBS. Next, the samples were incubated with the biotinylated rabbit anti-goat IgG for Tyro 3, Axl, and Mer or goat anti-rabbit IgG for Gas6 for 30 min and then with streptavidin-peroxidase complex for 30 min. After three 5-min washes in PBS, the peroxidase-binding sites of all the samples were demonstrated by the diaminobenzidine method. All sections were counterstained with hematoxylin and mounted with Canada balsam (Sigma; St Louis, MO) for observation after dehydrated in ethanol and clarification with xylene. The primary antibodies were used at 1:400 dilutions for Tyro 3 and Axl or 1:800 for Mer and Gas6. Negative control sections were incubated with preimmune goat serum or rabbit serum instead of primary antibody, or with primary antibody preabsorbed with blocking peptide.
Isolation of Sertoli Cells and Germ Cells
The procedure for the isolation of Sertoli cells and germ cells was based on previous description (Cheng et al. 1986) with a modification. Briefly, decapsulated testes from d35 mouse were incubated with collagenase (0.5 mg/ml) for 15 min with gentle oscillation, then were filtered through 80-μm copper meshes to eliminate Leydig cells. Tubules were suspended in collagenase for another 20-min incubation to remove myoid cells. Tubules were then incubated with hyaluronidase (1 mg/ml) for 25 min with gentle oscillation and pipetting. The resulting cells were washed four times with F12/DMEM, then were plated on Lab-Tek Chamber slides (Nunc Inc.; Naperville, IL) at 4 × 106 cells/ml of F12/DMEM supplemented with sodium bicarbonate (1.2 mg/ml), penicillin (100 U/ml), streptomycin (100 μg/ml), and 10% FBS. Cells were maintained in a humidified atmosphere of 95% air: 5% CO2 (v/v) at 32C for 48 hr. Thereafter, Spermatogenic cells were collected by gentle pipetting from Sertoli cells and dropped onto poly-L-lysin-coated slides for immunostaining. The germ cells adhered to Sertoli cells were lysed by a hypotonic treatment with 20 mM Tris, pH 7.4, for 2.5 min. Sertoli cells were cultured for an additional 24 hr for immunocytochemistry staining.
Immunocytochemistry Staining
Briefly, the slides were washed with PBS (10 mM sodium phosphate, 0.15 M NaCl, pH 7.4) three times and were fixed with cold methanol at -20C for 1 min. The endogenous peroxidase activity was blocked by treatment with 0.3% H2O2 in methanol for 15 min using 0.3% Triton X-100 in PBS for 15 min to increase cellular permeability. After blocking by preincubation with 10% normal goat serum in PBS at room temperature for 1 hr, the slides were then incubated with anti-Tyro 3 or anti-Axl antibody at a dilution of 1:100 at 4C in a humidified chamber overnight. The succeeding procedures are identical to the corresponding ones in immunohistochemistry staining.
Results
To test specificity of the antibodies used in this study, we performed a Western blot using the antibodies for the lysates of d35 testis. The results (Figure 1) were consistent with those provided by the manufacturer. Single specific band for each antibody was detected, suggesting the monospecificity of each antibody.
In the testes of different aged mice, all three members of Tyro 3 family receptors and their ligand Gas6 were immunolocalized in different testicular cells. The immunostaining signals for both Tyro 3 and Axl, during early development, were restricted to Sertoli cells. However, Mer and Gas6 showed a strong immunopositive staining in Leydig cells and primitive spermatogonia, although occasional lightly immunopositive Sertoli cells were seen.
The Tyro 3 immunostaining of Sertoli cells was strongly positive during the whole process of the postnatal development from d3 to d35. No positive staining was detected in spermatogenic cells, peritubular myoid cells, or interstitial cells. Distribution of Tyro 3 in Sertoli cells varied with the testicular development. At d3 and d7 (Figures 2A and 2B), the entire Sertoli cell except for the nucleus was strongly stained. At d14 (Figure 2C), immunostaining was concentrated at the adlumenal compartment of seminiferous tubules, only weakly positive or no positive staining was detected in the basal compartment. At d21 and d35 (Figures 2D and 2E), protein Tyro 3 localized uniformly on the processes of Sertoli cells surrounding spermatocytes and spermatids. Negative or very weak immunostaining was seen in the basal compartment including spermatogonia and preleptotene spermatocytes. No positive staining was seen in the perinuclear region of the Sertoli cells.
The immunostaining intensity and distributions of Tyro 3 on Sertoli cells in mature testis (d56) were stage-dependent, although immunoreactivity was detected in all the spermatogenic stages (Figures 3A and 3B). The stage-dependent immunoexpression and distribution of Tyro 3 in mature testes were evident in all animals examined. In stages IX-XII when no round spermatid were present in the seminiferous epithelium, the intensity of the staining was low and mainly appeared around zygotene spermatocytes, diplotene spermatocytes, and elongating spermatids. In stages I-VI, the staining was higher around spermatocytes and round spermatids. In contrast, in stages VII-VIII, when more round spermatids and elongated spermatids were formed, the strong staining appeared in the adlumenal compartment, particularly around the heads of elongated spermatids.
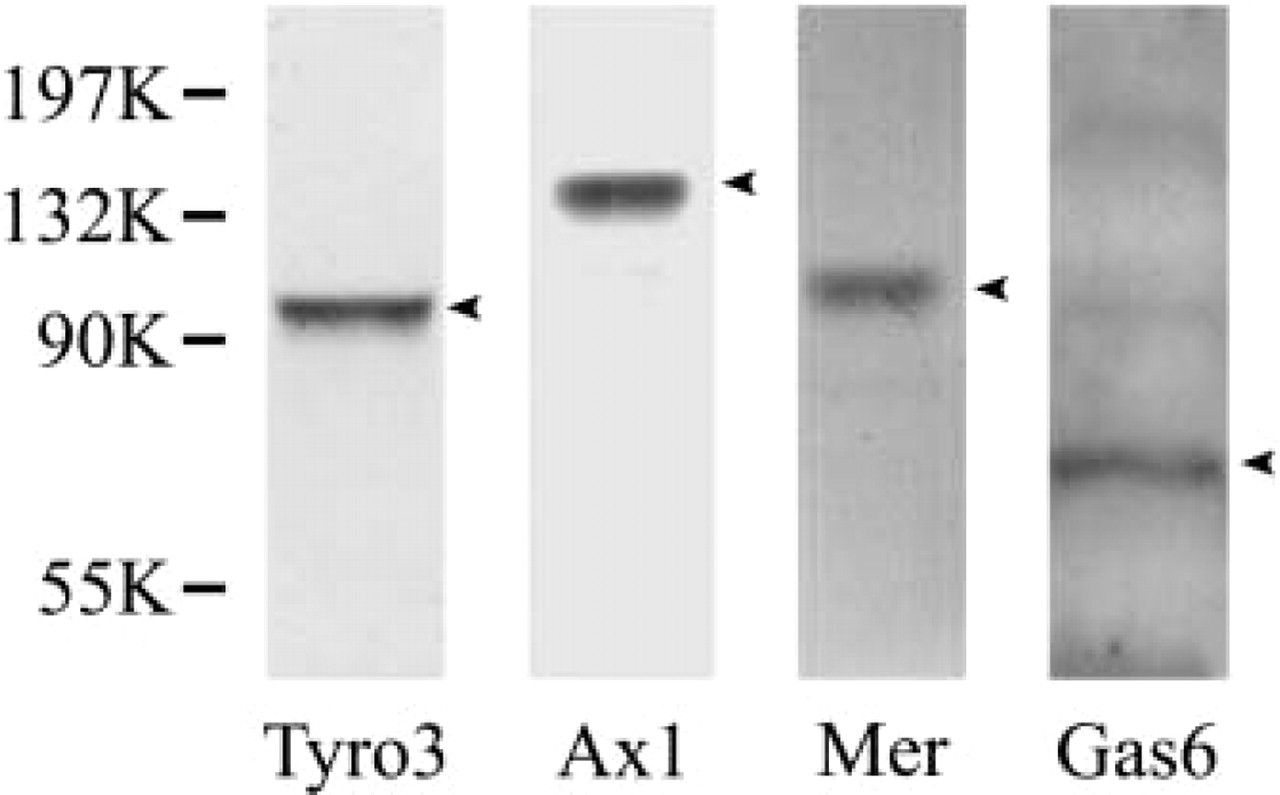
Specificity of antibodies against Tyro 3, Axl, Mer, and Gas6. Day 35 mouse testis was lysed with Triton X-100 and SDS. Western blotting was performed with goat anti-Tyro 3, Axl, Mer, and rabbit anti-Gas6 antibodies. A single band for each antibody was developed.
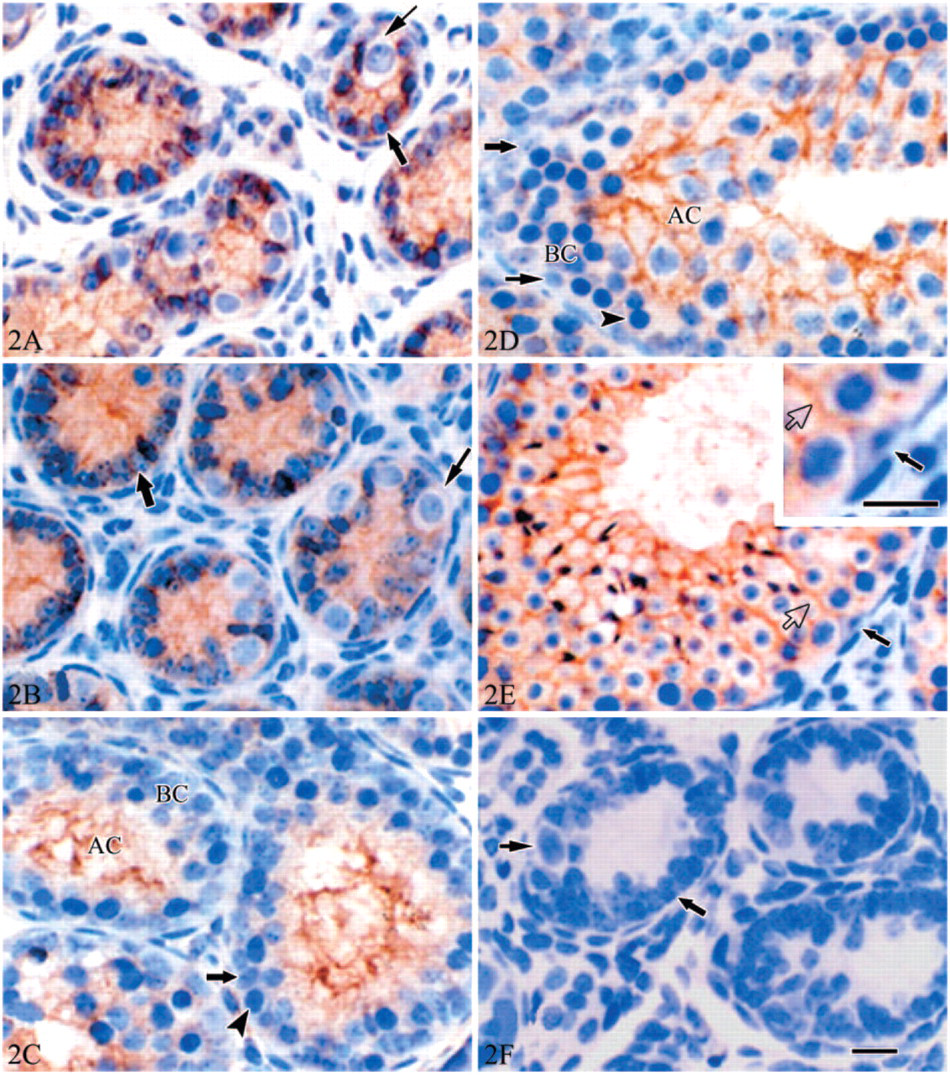
Tyro 3 immunoexpression in the developing mouse testis. At postnatal day 3 (
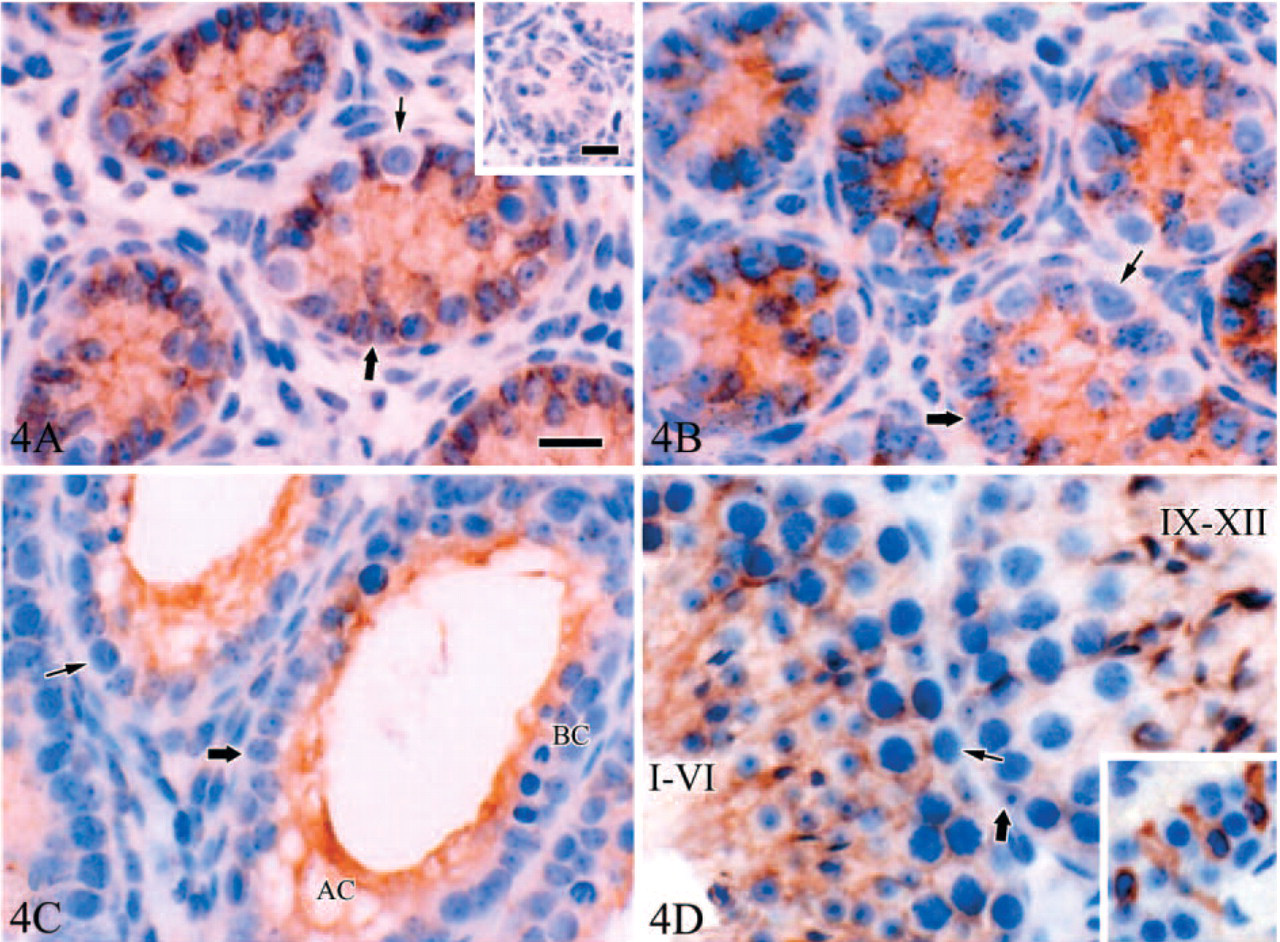
Similarly to the Tyro 3 pattern, Axl immunoexpression was evident only in Sertoli cells in early development mice, from d3 to d35 (Figures 4A–4C). All other cell types were negative. In mature testis (d56), Axl also showed a stage-dependent expression in Sertoli cells (Figure 4D). However, Axl immunostaining in mature testis was seen in some Leydig cells (islet in Figure 4D). Two types of Leydig cells were recognized in mature testis (Browne et al. 1990; Ariyaratne et al. 2000; Sipahutar et al. 2003): spindle-shaped Leydig cells with spindle-shaped nuclei and polygonal Leydig cells with round nuclei. Interestingly, intense Axl immunostaining was evident in some spindle-shaped Leydig cells, but not in the polygonal Leydig cells.
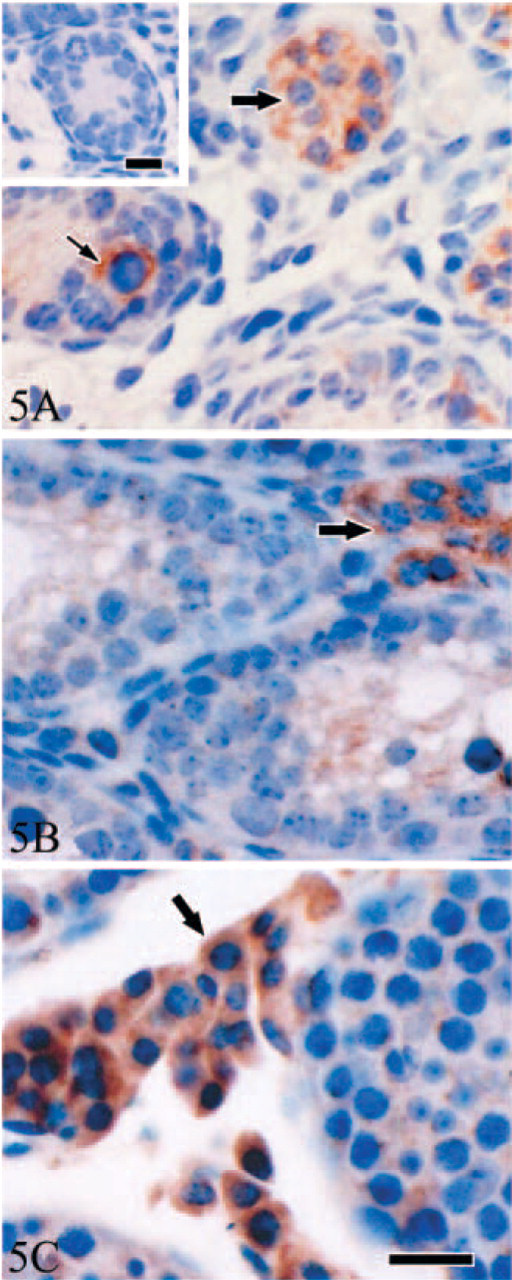
Unlike Tyro 3 and Axl, at d3, protein Mer immunostaining was prominently observed in the cytoplasm of primitive spermatogonia and of some interstitial cells and was only weakly detectable in Sertoli cells (Figure 5A). Based on morphology, immunoreactive positive interstitial cells were Leydig cells. In d7 testis, the Mer expression pattern was identical (not shown). However, from d14, spermatogonia became negative, whereas a strong cytoplasmic staining persisted through d35 in the Leydig cells (Figures 5B and 5C). A relatively weak immunostaining was also observed in the processes of Sertoli cells between spermatocytes throughout postnatal testis development.
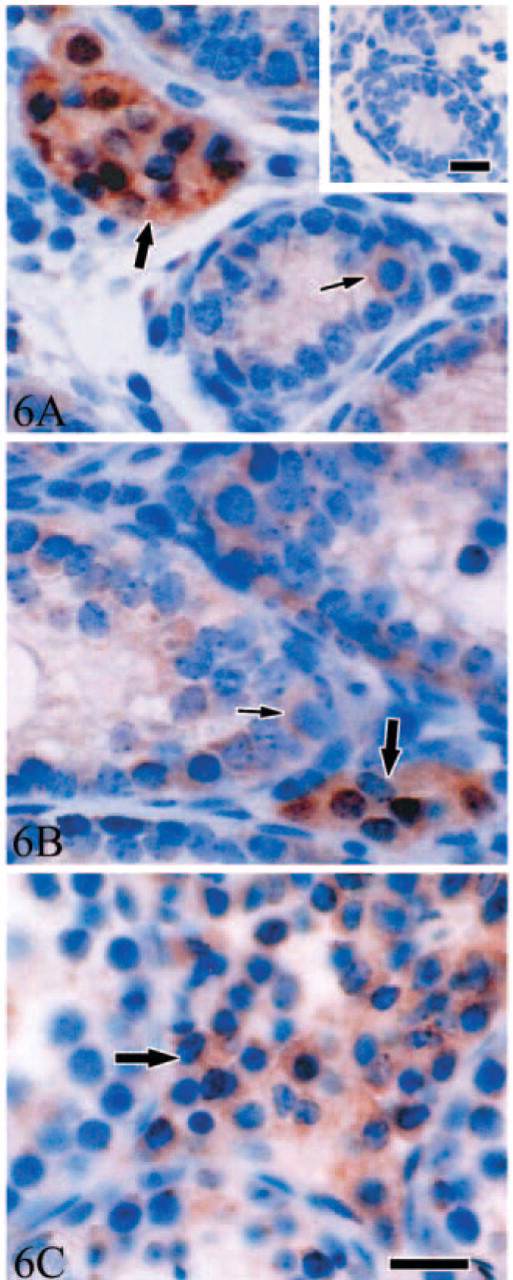
A distinct immunoexpression pattern of Gas6 was also demonstrated by immunohistochemistry (Figure 6). An intense Gas6 immunostaining was evident only in Leydig cells from postnatal d3 to d35. However, a relatively weak positive signal was also detected in cytoplasm of Sertoli cells and spermatogonia at d3, d7, and d14 (Figures 6A and 6B). At d35 and d56, intense Gas6 immunostaining was still evident in Leydig cells, whereas staining signal disappeared in spermatogonia by this time (Figure 6C).
To make comparative evaluation of the expression level in different testicular cells, the testes from 7-day-old mice were used for a semiquantitative evaluation of immunoexpression of Tyro 3, Axl, Mer, and Gas6. The testes from three animals and five sections from each testis were examined for each protein. The relative intensity of the immunostaining (brown staining) was evaluated as negative (-), weak positive (+), or strong positive (+ + +). The similar immunostaining phenotype in all sections of different animals for each protein appeared. The results are shown in Table 1.
To further exclude Tyro 3 and Axl immunoexpression in germ cells, d35 testis was used to isolate germ cells and Sertoli cells. The immunocytochemistry staining was performed in the isolated cells. The results (Figure 7) showed that Tyro 3 and Axl were expressed only in Sertoli cells; no staining signal was detected in germ cells.
Discussion
Tyro 3, Axl, and Mer belong to a newly identified family of cell adhesion molecule-related receptor tyrosine kinase, and they are widely expressed in various tissues. A recent study has shown that the Tyro 3 receptor family plays essential roles in regulating mammalian spermatogenesis, although its mechanism is unknown (Lu et al. 1999). The purpose of the present studies was to follow the expression pattern of Tyro 3 receptor family and its ligand in postnatal developing testis with an effort to provide clues for the further investigation of the receptors functions in regulating spermatogenesis.
The results demonstrated that Tyro 3 and Axl were expressed in Sertoli cells and their expression intensities did not show obvious changes during postnatal testicular development. We have also shown that the expression of Tyro 3 and Axl showed a stage-dependent pattern in mature testis. Expression of Mer was mainly observed in Leydig cells and primitive spermatogonia, whereas intense immunostaining of Gas6 was only evident in Leydig cells. In contrast with Tyro 3 and Axl, both Mer and Gas6 showed relative weak expression in Sertoli cells. This is the first time the details of the immunoexpression patterns of Tyro 3 subfamily receptors and their ligand Gas6 during development of mouse testis after birth have been examined.
Although a few previous studies have examined the expression of Tyro 3, Axl, Mer, and Gas6 mRNA by RT-PCR and Northern blot (Chan et al. 2000; Wong and Lee 2002), the results were not consistent. Wong and Lee (2002) reported that the expression of Tyro 3 and Axl decreased during postnatal testicular development, and Chan et al. (2000) concluded that Axl was not expressed in Sertoli cells. However, it must be emphasized that Wong's study was performed by using total RNA extracted from testes of different age mice. In neonatal testis, the seminiferous tubules are mainly composed of Sertoli cells; the percentage of Sertoli cells in developing and mature testes are obviously decreased because of the increasing number of spermatogenic cells. Therefore, Wong's results probably reflect the reduced ratio of Sertoli cells versus germ cells during testicular development, instead of the actual expression levels of Tyro 3 and Axl in Sertoli cells. As for Chan et al.'s results, they were acquired from testicular clonal cell lines in which the expression of Tyro 3, Axl, and Mer might not really reflect the expression in vivo. Our study represents the first analysis of Tyro 3, Axl, and Mer proteins by immunostaining of developing testes, whereas previous analyses were performed at the mRNA level.
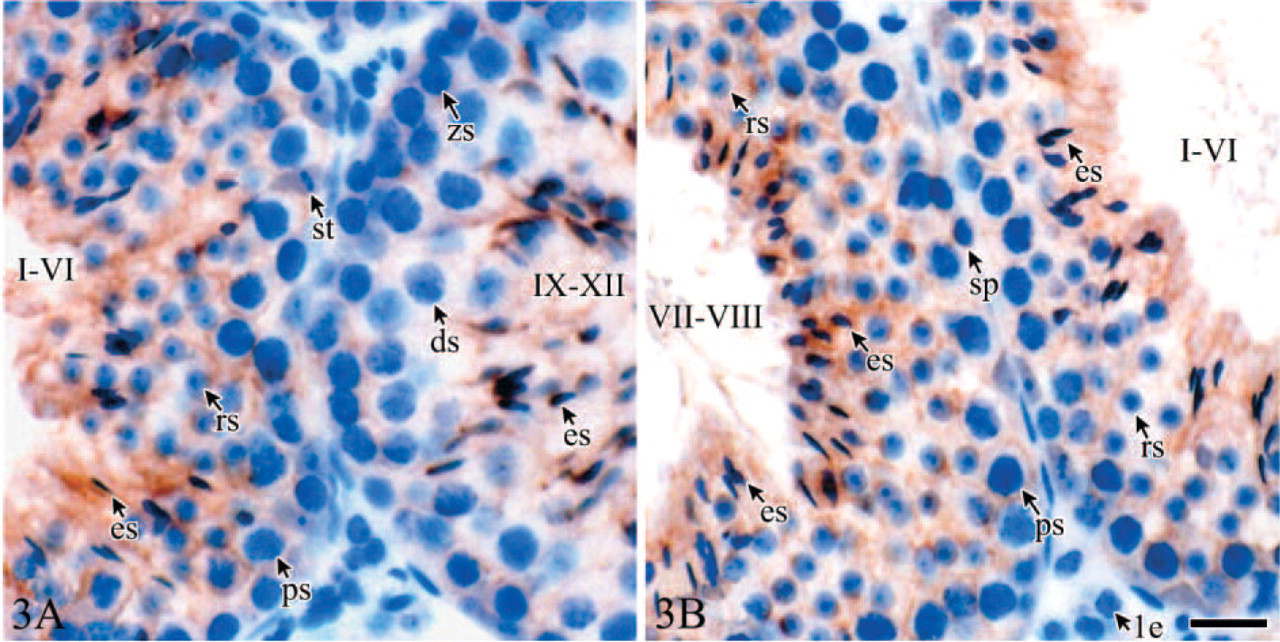
Stage-dependent immunostaining for the Tyro 3 in day 56 testis (
Distributions and relative staining intensities of Tyro 3 family receptor Tyro 3, Axl, Mer, and their ligand Gas 6 in mice testis at postnatal day 7
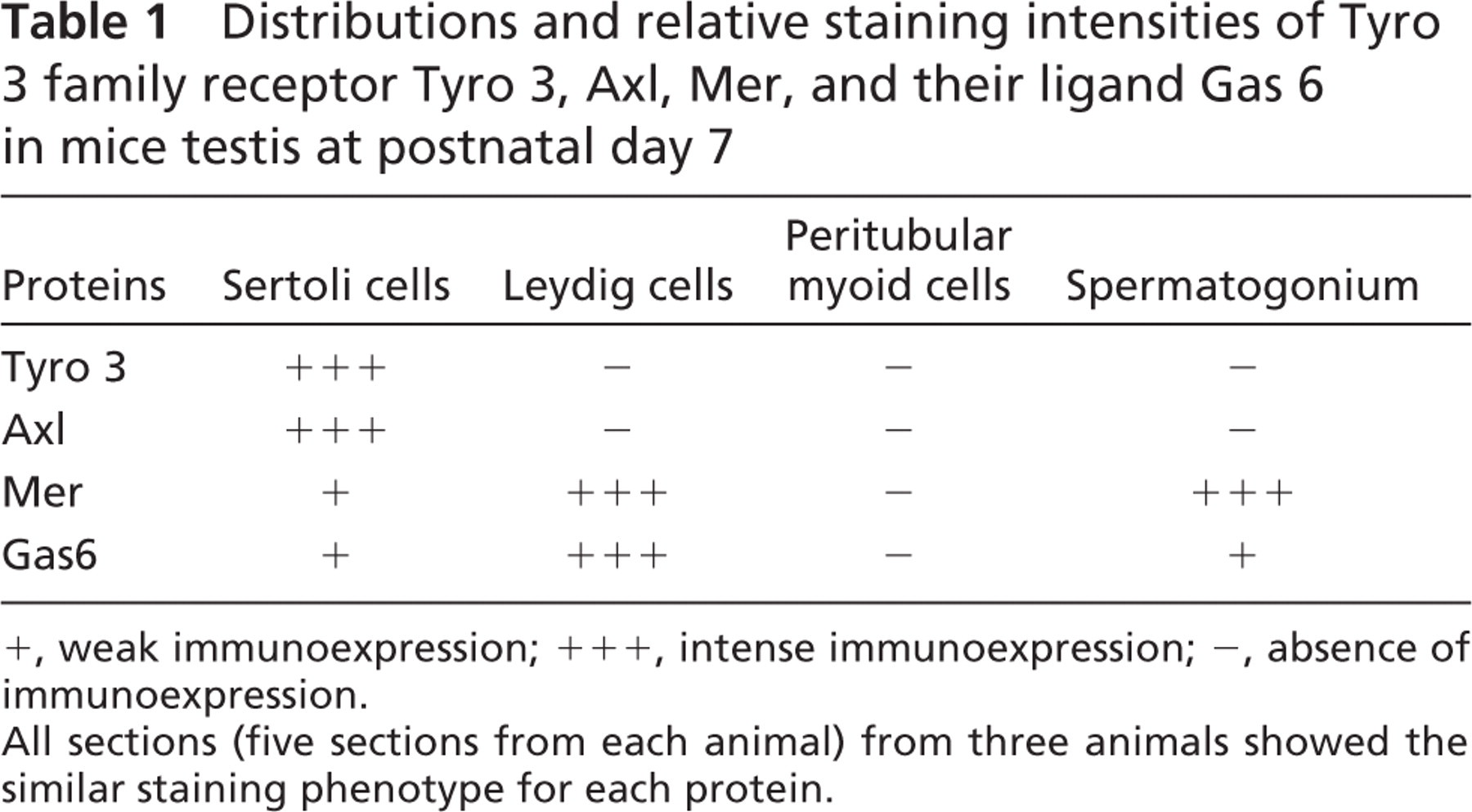
+, weak immunoexpression; + + +, intense immunoexpression; -, absence of immunoexpression.
All sections (five sections from each animal) from three animals showed the similar staining phenotype for each protein.
Although Tyro 3, Axl, and Mer belong to the same family and share high-sequence homology, they have distinct localization in testicular cells. Tyro 3 was expressed only in Sertoli cells, Axl was expressed in Sertoli cells and some Leydig cells, and Mer was expressed mainly in Leydig cells and in primitive spermatogonia. Sertoli cells and Leydig cells are the main somatic cells in the testis and play important roles in testicular development and spermatogenesis. As a nurturer, Sertoli cells provide essentially physical and trophic support for developing spermatogenic cells. Sertoli cells can also secrete an androgen-binding protein, which is necessary for spermatogenesis, under the control of follicle-stimulating hormone and testosterone. Leydig cells in interstitial tissue of testis can produce the male hormone testosterone under the control of luteinizing hormone. It has been reported (Lu et al. 1999) that Tyro 3, Axl, and Mer genes must all be disrupted for maximal degeneration of spermatogenesis. Considering our present results, we postulate that Tyro 3, Axl, and Mer may each play different roles and collaboratively control spermatogenesis.
Mammalian spermatogenesis is a highly synchronized, regular, long, and extremely complex process of cellular differentiation by which a spermatogonial “stem cell” is gradually transformed into highly differentiated haploid spermatozoa. In adult mammals, spermatogenesis is a continuous process that can be divided into three distinct phases (mitosis, meiosis, and spermiogenesis), each characterized by specific morphological and biochemical changes of nuclear and cytoplasmic components. Interestingly, our results showed that distribution of Tyro 3 or Axl in Sertoli cells varied with the testicular development and spermatogenesis. At postnatal d3 and d7, when Sertoli cells and spermatogonia proliferate, Tyro 3 and Axl were distributed uniformly in the plasma membrane and cytoplasm of Sertoli cells, suggesting that they may play roles in regulation the proliferation of spermatogonia and Sertoli cells in early stage of testis. At postnatal d14, when mature Sertoli cell function began to develop (e.g., the formation of the blood-testis barrier, the expression of androgen-binding protein, the production of seminiferous fluid) (Gondos and Berndtson 1993), Tyro 3 and Axl mainly concentrated on the adlumenal compartment of seminiferous tubules. At postnatal d21, with the first wave of spermatogenesis, Tyro 3 and Axl were localized in the processes of Sertoli cells that surrounded the spermatocytes and round spermatids. At d35 and after, the expression of Tyro 3 and Axl became stage-dependent in Sertoli cells. Immunostaining was stronger in stages I-VIII when many round spermatids and elongating spermatids are developing. At stages VII and VIII, positive staining mainly focused on the regions near the lumen of seminiferous tubules where elongating spermatids contacted the processes of Sertoli cells by apical ectoplasmic specializations. This periodic cycling with tubule stage is a property exhibited by many other Sertoli cell gene products, including the transcription factor GATA-1, which is expressed only at stages VII-VIII (Yomogida et al. 1994) and androgen receptor, which has a highest expression at stages VII-VIII (Bremner et al. 1994). The stage-dependent expression patterns of Tyro 3 and Axl suggest that they may involved in the differentiation of Sertoli cells to regulate spermatogenesis.
A few previous studies have shown Mer-mediated phagocytosis and clearance of apoptotic cells by macrophage (Scott et al. 2001; Cohen et al. 2002). In testis, Sertoli cells are special cells and function as macrophage by maintaining the integrity of the seminiferous epithelium. During spermatogenesis, Sertoli cells phagocytose apoptotic germ cells and residual bodies; therefore, it is worthwhile to investigate whether the receptors play roles in regulating the phagocytosis of Sertoli cells.
Expression of Axl in mouse testis. At day 3 and day 7 (
Expression of Mer in mouse testis. At day 3 (
Expression of Gas6 in mouse testis. At day 3 (
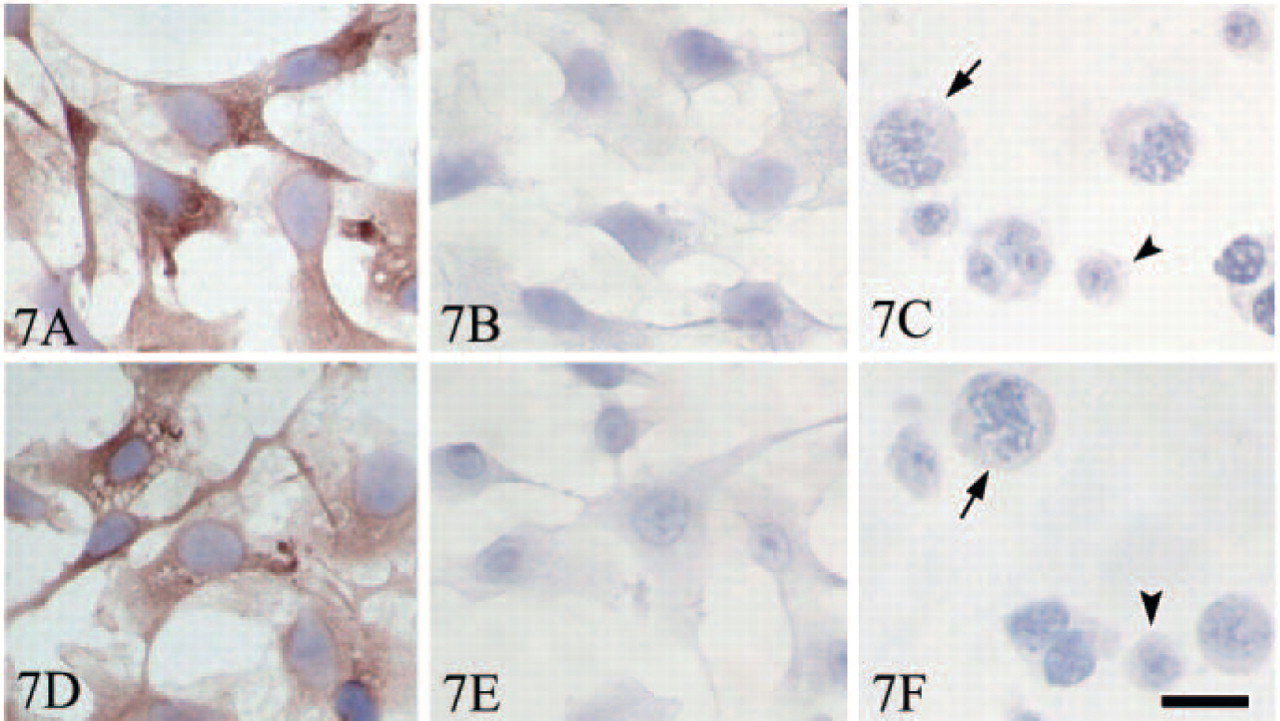
Tyro 3 and Axl immunoexpression in Sertoli cells and germ cells isolated from day 35 testis. Tyro 3 is expressed in isolated Sertoli cells (
As the common ligand for Tyro 3, Axl, and Mer, the immunoexpression of Gas6 in testis was also examined. The expression pattern of Gas6 in mouse testis was previously reported by Lu et al. (1999). In agreement with their results, we observed that Gas6 is mainly expressed in Leydig cells. However, we did not observe the periodic cycling of Gas6 in Sertoli cells as they observed by in situ hybridization. Actually, only a relatively weak immunostaining signal was detected in Sertoli cells in our present study. The controversy with the results of Lu et al. may depend on the fact that we studied the protein expression by immunohistochemistry, whereas they analyzed mRNA by in situ hybridization. It has been reported that Gas6 was a potential growth factor that stimulated human Schwann cell proliferation through the Axl/Tyro 3 tyrosine kinase receptors (Li et al. 1996). Gas6 expression by Sertoli cells in vitro was stimulated by forskolin, and this forskolin-stimulated Gas6 expression was accompanied by an increase in Tyro 3 phosphorylation (Chan et al. 2000). These previous studies suggest that Gas6 may exert its biological effects through autocrine or paracrine patterns. Gas6 expression pattern in testis suggests that it may play roles in spermatogenesis by regulating behaviors and functions of Sertoli cells.
Based on the structure motif, Tyro 3 family receptors have two potential functions: cell proliferation and adhesion. Previous studies have showed that Tyro 3 family is related to cell proliferation. In some malignant tumors, such as leukemia (O'Bryan et al. 1991), gastric cancer (Wu et al. 2002), uterine leiomyoma (Sun et al. 2003a), uterine endometrial cancer (Sun et al. 2003b), and ovarian cancer (Sun et al. 2004), the members of tyro3 receptor family and their ligand Gas6 were overexpressed. That Mer and Gas6 express in spermatogonia suggests that they may relate to proliferation of spermatogonia. A few studies also confirmed that the Tyro 3 family could induce homophilic binding independent of ligand in fibroblasts (Bellosta et al. 1995) or mediated by soluble Gas6 in the 32D myeloid cell line (McCloskey et al. 1997). Adherens junctions between Sertoli cells and spermatogenic cells are crucial not only for mechanical adhesion, but also for cell morphogenesis and differentiation during spermatogenesis. In the testis of triple mutant (Axl−/-Tyro3−/-Mer−/-) mice that completely lose spermiogenesis, no obvious changes in amounts of Sertoli cells in which Axl and Tyro 3 were expressed and of Leydig cells in which Mer was expressed were observed, whereas apoptotic spermatogenic cells apparently increased (Lu et al. 1999), suggesting that proliferation of Sertoli cells and Leydig cells was not affected apparently by Tyro 3 family. The present study shows that Axl and Rse are mainly distributed in the processes of Sertoli cells that contact spermatogenic cells, particularly with round and elongating spermatids. Based on these observations, it would be interesting to ask if the Tyro 3 family receptors are involved in adhesion between Sertoli cells and germ cells. We are investigating this possibility.
In summary, in the present study, we have clearly shown that Tyro 3 family receptors and their ligand Gas6 are differentially expressed at the protein level in different testicular cells. Based on previous studies, these observations further suggest that Tyro 3 family receptors and their ligand Gas6 may participate in testicular development and spermatogenesis by regulating the function of Sertoli cells and Leydig cells. Our results provide clues to further study the mechanism of Tyro 3 family in regulating spermatogenesis.
Footnotes
Acknowledgements
This work was supported by Special Funds for Major State Basic Research Project of China (Grant No. G1999055901).