
Abstract
We investigated the phenotypic features of cardiomyocytes, including the gap junctions, in the myocardial sleeve of thoracic veins. Single cardiomyocytes, isolated from the canine pulmonary veins (PV) and superior vena cava (SVC) using digestive enzymes, were examined by immunoconfocal microscopy using antisera against connexin43 (Cx43), Cx40, and other cell markers. The results showed that isolated cardiomyocytes displayed rod shapes of various sizes, ranging from <50 μm to >200 μm in length, and all the cells expressed α-actinin and vinculin. Gap junctions made of various amounts of Cx43 and Cx40 were found at the cell borders. These two connexins were extensively co-localized. Comparison between the thoracic veins showed that cells of the SVC contained more Cx43 gap junctions (total Cx43 gap junctions area per cell surface area, 4.0 ± 0.2% vs 1.5 ± 0.2%; p<0.01). In addition, for single-nucleus cells, those from the PV were longer (103.7 ± 3.6 vs 85.0 ± 3.1 μm; p<0.01) but narrower (14.4 ± 0.5 vs 16.9 ± 0.9 μm; p<0.01). In conclusion, canine thoracic veins contain cardiomyocytes with differences in shape and gap junctions, suggesting that the electrical conduction properties may be different between the thoracic veins.
E
Gap junctions, made of molecules belonging to the connexin multigene family, are clusters of cell membrane aqueous channels, which in the heart provide a low-resistance pathway for electrical coupling between the adjacent cardiomyocytes. In mammals, Cx40, Cx43, and Cx45 are present in the atrial cardiomyocytes (Gros and Jongsma 1996; Severs et al. 2001). Changes in the expression of the connexins has been shown to be associated with a variety of cardiac pathologies and contribute to the development of cardiac arrhythmia (Peters et al. 1993,1995; Lerner et al. 2000; Gutstein et al. 2001). Although information about the distribution of gap junctions and expression of connexins in the myocardial sleeve of the thoracic veins has just emerged (Yeh et al. 2001), whether or not the expression of connexins differs between the thoracic veins remains unclear.
In this study we investigated the phenotypes, including the morphology and gap junctions, of the cardiomyocytes isolated from canine PV and SVC. Our results show that a marked heterogeneity exists in the cardiomyocytes of the venous wall.
Materials and Methods
Isolation of Cardiomyocytes
Cardiomyocytes were isolated from five adult mongrel dogs (20–25 kg). Animals were anesthetized with sodium pentobarbital (30 mg/kg IV). After thoracotomy the hearts were rapidly removed and immersed in normal Tyrode solution (containing 137 mM NaCl, 4 mM KCl, 15 mM NaHCO3, 0.5 mM NaH2PO4, 0.5 mM MgCl2, 2.7 mM CaCl2, and 11 mM dextrose) equilibrated with a gas mixture (97% O2 and 3% CO2) at room temperature (RT). The left atrium (LA), PV (cut at lines about 5 mm proximal to the LA-PV junction and 20 mm distal to the ending of myocardial extension onto PV), and SVC (cut at lines about 5 mm proximal to the RA-SVC junction and 20 mm distal to the ending of myocardial extension onto SVC), were dissected out. The tubelike isolated PV and SVC were turned inside out and the proximal end and side branches were ligated, leaving the distal end connected to a Langendorff perfusion column via a polyethylene tubing. Perfusion (500 ml/hr) started with oxygenated normal Tyrode solution (37C) and was then replaced with Ca2+-free oxygenated Tyrode solution containing 300 U/ml collagenase (Type I; Sigma, St Louis, MO) and 0.5 U/ml protease (Sigma; Type XIV). After softening, the atrial end of the samples was cut off at about 5 mm. The remaining samples were cut into fine pieces and gently shaken in 5–10 ml of Ca2+-free oxygenated Tyrode solution to free single cardiomyocytes. LA myocytes were also isolated through the perfusion of collagenase and protease. The isolated cells were fixed with 2% paraformaldehyde (pH 7.2) for 10 min, washed with Ca2+-free Tyrode solution, spun (250 rpm, 5 min) onto silanized slides (DAKO; Glostrup, Denmark) using a cytospin (Life Science International; Cheshire, UK), and stored at −30C in PBS containing 0.02% NaCN3. The work conforms with the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health (NIH Publication No. 85–23, revised 1996) and with the ROC Animal Protection Law (Scientific Application of Animals), 1998.
Immunocytochemistry
Antibodies. The polyclonal antisera against Cx43 and Cx40 were produced in rabbits [designated Cx43 (R530) and Cx40 (R2), respectively], a chicken [Cx43(Chk)], and a guinea pig [Cx40 (GP8)] against the synthetic peptides corresponding to residues 314–322 [for Cx43] or 254–268 (for Cx40) of the cytoplasmic C-terminal tail of rat connexins. The polyclonal antiserum against Cx45 was produced in a rabbit [Cx45 (R1)] and a rat [Cx45 (Rat9)] against a synthetic peptide corresponding to residues 354–367 of the cytoplasmic C-terminal tail of human Cx45. Affinity-purification and confirmation of the specificity by Western blotting and immunoconfocal microscopy were conducted as previously reported (Yeh et al. 1998,2000), using connexin-expressing HeLa cells, including HeLa cells expressing Cx37 (HeLa-37), HeLa-40, HeLa-43, HeLa-45, and the wild-type (HeLa-W). All the cells were kindly donated by Professor Klaus Willecke [University of Bonn, Germany (Elfgang et al. 1995)]. Cardiomyocytes were identified using mouse monoclonal antiserum against α-actinin and vinculin (Sigma). Smooth muscle cells were identified using smooth muscle α-actin (Dako). For secondary antibodies, donkey anti-rabbit, anti-chicken, or anti-guinea pig immunoglobulin conjugated to either CY3 or CY5, and anti-mouse to CY5 (Chemicon; Temecula, CA) were used. For single labeling, CY3-conjugated antibodies were used.
Immunolabeling of Connexins and Cell Markers. For single labeling, cells were blocked in 0.5% BSA (15 min) and incubated with the anti-Cx43 [1:100 for Cx43 (R530); 1:50 for Cx43 (Chk)], anti-Cx40 [1:200 for both Cx40 (R2) and Cx40 (GP8)], anti-Cx45 (1:50 for both Cx45 (R1) and Cx45 (Rat9)], anti-α-actinin (1:200), anti-vinculin (1:200), or anti-smooth muscle α-actin (1:50) at 37C for 2 hr. The samples were then treated with CY3-conjugated secondary antibody (1:500, RT, 1 hr). In experiments in which each of the connexins and α-actinin were simultaneously localized in the same samples, incubation was with a mixture of one of the polyclonal antisera (the same dilution as in the single-labeling experiment) plus anti-α-actinin (1:200), followed by incubation with a mixture of a CY3-conjugated antibody (for labeling connexin) and anti-mouse CY5 (for α-actinin). Double labeling of Cx40 and Cx43 was conducted using Cx40 (GP8) and Cx43 (R530), followed by incubation with a mixture of anti-guinea pig CY3 and anti-rabbit CY5. Between each step, the slides were thoroughly washed with PBS. Finally the cells were mounted. All experiments included cryosections of dog left atrium, fixed by methanol at −20C, and the isolated LA cells as positive controls and omission of primary antibody as negative controls. In parallel, cells were counterstained with bisbenzamide (1 μg/ml; Sigma) for 15 min to visualize their nuclei.
Confocal Laser Scanning Microscopy and Image Analysis
Stained samples were examined by confocal laser scanning microscopy using a Leica TCS SP equipped with argon/krypton and UV laser with the appropriate filter spectra adjusted for the detection of CY3, CY5, and bisbenzamide fluorescence. Consecutive optical sections taken at 0.5-μm intervals through the full thickness of cells were collected and transformed into projection views. The images from cells of double labeling were taken using sequential dual-channel scanning. Quantification of the cell size and Cx43-labeled spots was conducted using QWIN image analysis software (Leica; Heidelberg, Germany). The number of Cx43 gap junctions in the cardiomyocytes, as detected by the Cx43 (R530), is expressed as percentage of the cell surface area. Data, expressed as mean values (± SE), were compared statistically by t-test. A p value less than 0.05 was considered significant.
Results
Characterization of Anti-connexin Antisera
Western blotting analysis showed that both anti-Cx43 antibodies recognized closely situated bands of about 43 kD in a cell membrane preparation of HeLa-43. Similarly, both anti-Cx40 antibodies recognized a single band of about 40 kD in the HeLa-40 cells, and both anti-Cx45 antibodies recognized a single band of about 45 kD in the HeLa-45 cells (Figure 1). For each group of experiments, the bands of Cx43, Cx40, and Cx45, each of which was absent in the other connexin-expressing HeLa cells and the wild-type cells (HeLa-W), were markedly inhibited by preincubation of the antibody with the peptide to which it was raised.
Immunoconfocal microscopy of the HeLa cells incubated with each of the anti-connexin antisera demonstrated that, in each corresponding connexin-expressing cell, distinct punctate labeling existed at cell-cell borders, typical of gap junctions. The labeling was abolished by peptide inhibition (Figure 2). No positive signal was apparent when the antibody was applied on the other HeLa transfectants and HeLa-W.
Immunoconfocal Examination of Single Cardiomyocytes
Microscopic examination showed that cells with various morphological features were freed from the venous wall after treatment with the digestive enzymes. Among the isolated cells, those of rod shapes were generally larger compared to cells of other shapes. Immunolabeling showed that the rod-shaped cells isolated from the PV and SVC bore cross-striations positive for α-actinin and vinculin (Figure 3). In general, the striation signals of α-actinin were much more conspicuous and better organized compared to those of vinculin, which also appeared at portions of cell borders. Such staining patterns of α-actinin and vinculin in the rod-shaped cells isolated from the venous wall were identical to the patterns seen in the LA working cardiomyocytes (Figure 3). In addition, no signal of smooth muscle α-actin was detected in any rod-shaped cells, thus confirming the cells with such features to be cardiomyocytes. On the other hand, all the striated cells were typically rod-shaped. Regarding the size, although cells as long as more than 200 μm and as short as less than 50 μm were observed, cells of extreme sizes were less common. Determinations of cell size should consider the possibility that cells in close attachment can be mistaken for single cells, especially for those possessing more than one nucleus, which appeared in about one third of the cells (Figure 3). Therefore, measurements were conducted on cells containing a single nucleus as unequivocally visualized in cells stained with bisbenzamide. Analysis of the cell size showed that the cardiomyocytes isolated from the PV were slightly but significantly longer and narrower than those from the SVC (p<0.01; Figure 4).
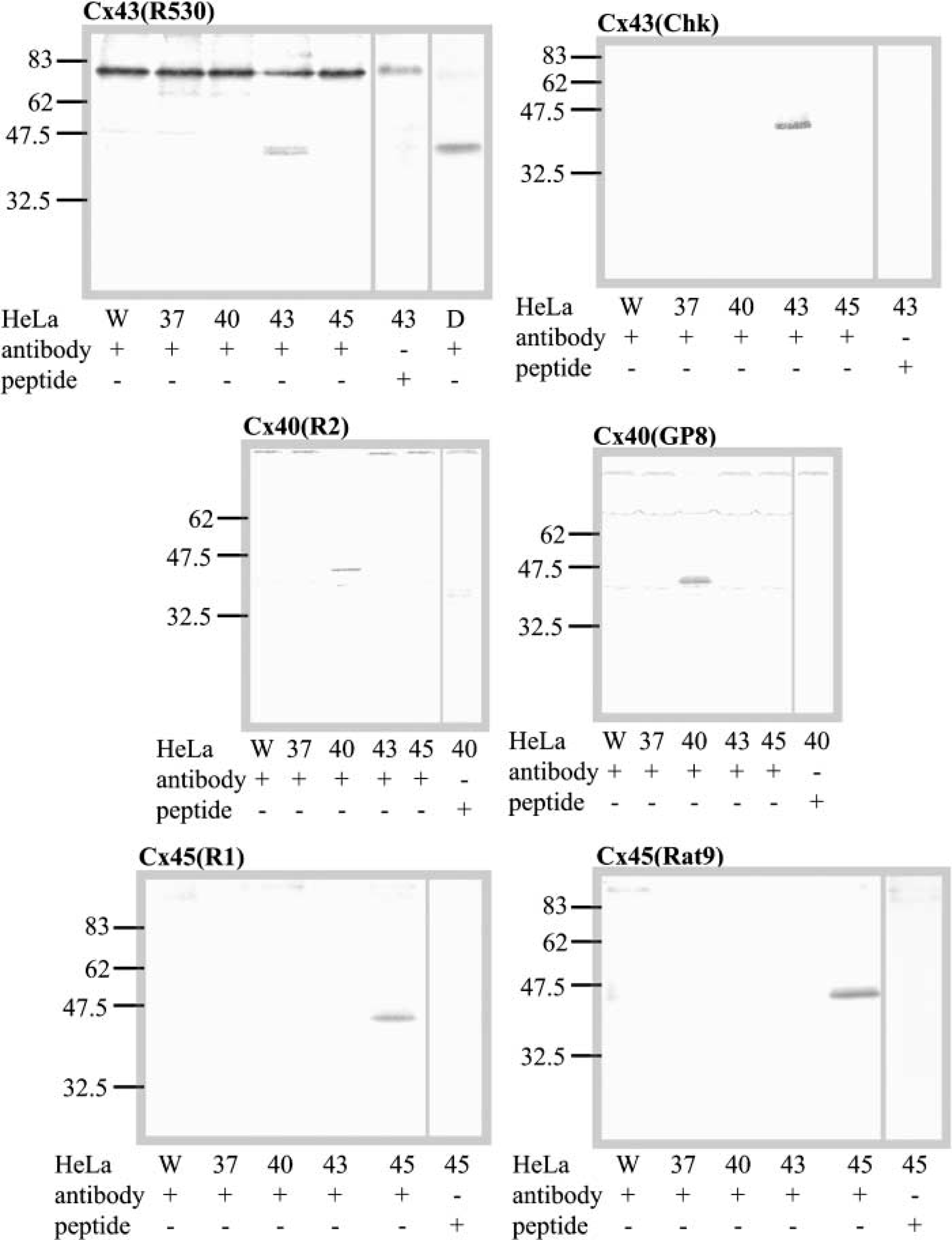
Characterization by Western blotting of anti-peptide antibodies against Cx43 (
Cx43-labeled spots were found clustered at domains of the cell borders (Figure 5), typical for the location of gap junctions. For the majority of cells, both ends of the long axis contained Cx43-labeled spots, although the amount varied widely among individual cells. Apart from the ends, Cx43 labeled spots were also detected at the lateral borders. Both Cx43 (R530) and Cx43 (Chk) antisera gave the same findings. For Cx40, detection with either Cx40 (R2) or Cx40 (GP8) showed a distribution pattern similar to that of Cx43 (Figure 5). Double labeling for both Cx40 and Cx43 demonstrated that more than 90% of the labeled spots were positive for both connexins.
Comparison between the thoracic veins showed that in the PV cardiomyocytes the number of Cx43-labeled spots was less (10.1 ± 0.9 per cell) and the size of the spots was smaller (0.5 ± 0.1 μm2), compared to the SVC cells (23.7 ± 1.4 per cell and 0.9 ± 0.1 μm2, respectively; both p<0.01). Considering the variation in cell size and that in some cases, as mentioned above, closely attached cells could not be distinguished from single cells, to overcome such interferences with interpretation, we present the data as the value of the total area of Cx43-labeled spots divided by the cell surface area for each cell or each group of closely attached cells. The result showed that a significant difference exists between the PV and the SVC (p<0.01; Figure 6).
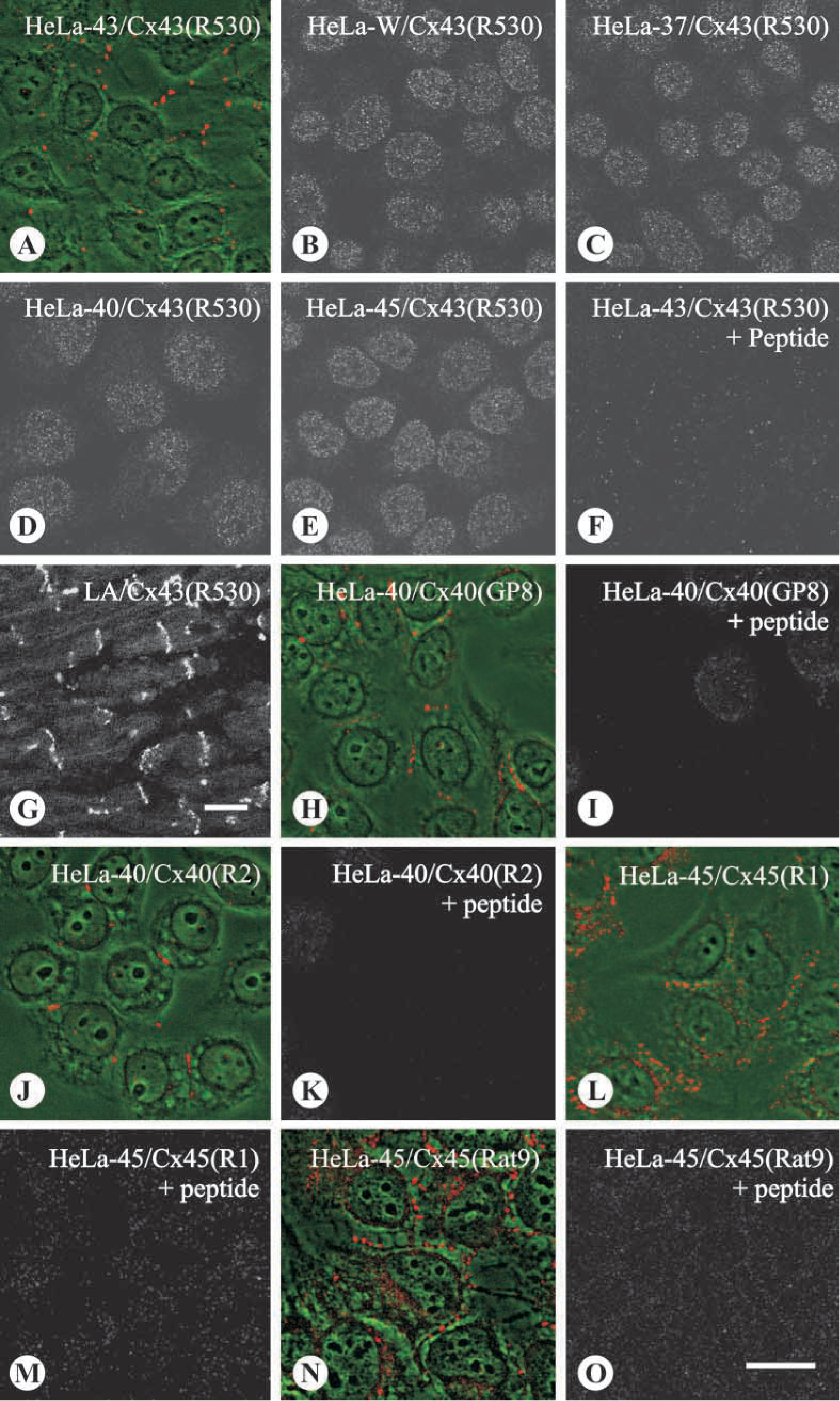
Characterization by immunocytochemistry of anti-peptide antibodies against Cx43 (
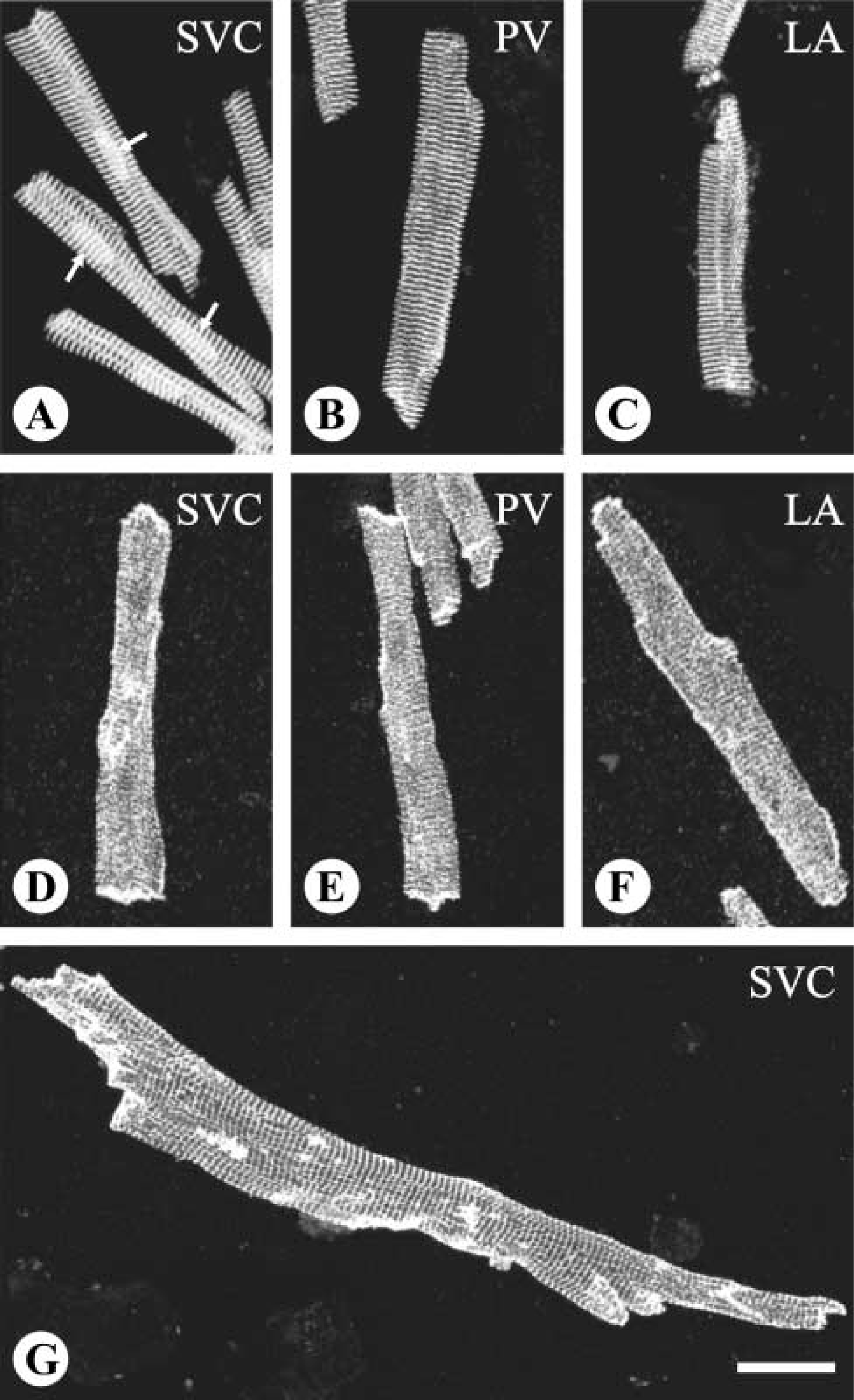
Phenotype identification of isolated cardiomyocytes with antisera against α-actinin (
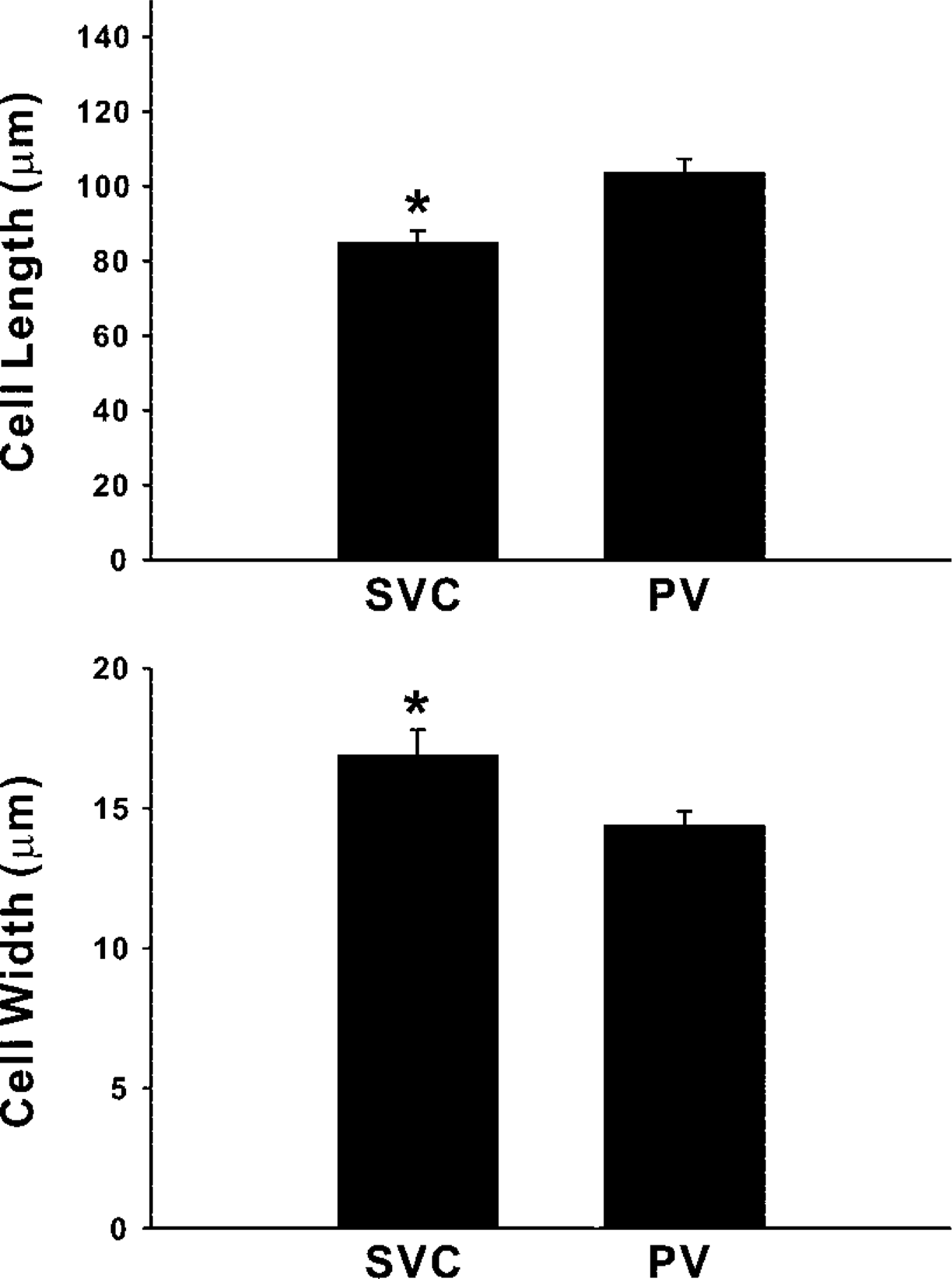
Comparison of the size of the cardiomyocytes between the thoracic veins. Upper histogram is for the length and lower histogram for the width. Data come from 52 (SVC) and 82 (PV) randomly selected single-nucleus cells. ∗ p<0.01 between the veins.
For Cx45, neither antisera Cx45 (R1) nor Cx45 (Rat9) gave signals in the isolated cardiomyocytes.
Discussion
The present study demonstrates that canine cardiomyocytes in the myocardial sleeve of the PV and SVC and those in the LA share a similar shape and immunocytochemically identified cross-striations. In addition, although situated in the venous wall, the cardiomyocytes differ phenotypically from the adjacent vascular smooth muscle cells. Furthermore, regarding the size of cardiomyocytes and the expression of gap junctions in the cells, a marked variation exists between the SVC and the PV. All these findings are based on the examination of cardiomyocytes freed from the tissues by enzyme digestion and by the application of a panel of affinity-purified anti-connexin antisera.
It should be noted that, in the present study, the two anti-Cx45 antisera, Cx45 (R1) and Cx45 (Rat9), give no signals in the isolated cells. Because the two anti-Cx45 are well characterized by Western blotting and immunocytochemistry using the connexin-expressing HeLa cells, one possible explanation for the absence of Cx45 signal in the isolated cells is the fixation method. The use of paraformaldehyde for the single cardiomyocytes in the present study may reduce the signal of Cx45, the expression level of which is known to be low in cardiac muscle (Coppen et al. 1998). In addition, data obtained from the isolated cells may not faithfully reflect the in situ environment. For example, the cell membrane may be injured during the course of isolation. Therefore, the structure of certain membrane-located molecules in the isolated cell, such as Cx45, may be changed and thus may affect the detectability of anti-Cx45.
The finding in the present study that cardiomyocytes isolated from the venous wall have similar expression patterns of Cx43 and Cx40 is consistent with our recent report that the majority of cardiomyocyte gap junctions in canine SVC contain both Cx40 and Cx43 (Yeh et al. 2001). In that study we also reported that, in the proximal potion of the SVC there are small groups of clustered cells mainly expressing either Cx40 or Cx43 (Yeh et al. 2001). However, cells with such an atypical expression pattern of connexins were not seen in the present study. Because the number of single cells freed from the tissues is far less than the actual number of the cells in the tissues, cells of a small population in the tissue, such as those with atypical connexin expression, may not be observed after the isolation procedure. On the other hand, our recent reports demonstrated that a substantial portion of cardiomyocytes isolated from the SVC and PV of the same animal possess membrane activity of spontaneous depolarization (Chen et al. 2001,2002). Considering the reported pacemaker activity in the SVC cardiomyocytes and that, anatomically, SVC is continuous with the SA node, one can suspect that the cardiomyocytes capable of spontaneous depolarization in the SVC possess features similar to those of the nodal cells. However, in the present study the strong striation signals of α-actinin (indicative of an abundant amount of contractile elements in the cytoplasm) in all rod-shaped cells provides evidence that the cardiomyocytes in the venous wall are structurally different from the nodal cells, in which contractile elements are few (Kwong et al. 1998).
One important finding from the present study is that differences exist between the PV and the SVC regarding the size of the cardiomyocytes and their expression of Cx43 gap junctions. Previous reports have shown that expression of connexins in the contractile myocardium is chamber-specific (Vozzi et al. 1999), and a recent report further demonstrates that formation of gap junctions in atrial and ventricular cardiac tissues is regulated primarily by intrinsic factors rather than by external hemodynamics (Kostin and Schaper 2001), although in other types of cells, e.g., endothelial cells, the expression of Cx43 is affected by the hemodynamics (Cowan et al. 1998; Gabriels and Paul 1998; DePaola et al. 1999). It remains unclear whether, in the present study, variations in cell size and in Cx43 gap junctions between the thoracic veins are attributed to environmental factors, such as oxygen tension in the lumen, which is higher in the PV than the SVC, or to intrinsic factors similar to those specific for cardiac chambers. However, the presence of distinct features in the cardiomyocytes of the thoracic veins suggests that the PV and the SVC may vary in electrical properties, which is supported by the clinical observation that, in atrial fibrillation in which the initiating foci are located in the venous wall, there is a marked difference in the number of patients between the PV and SVC (Haissaguerre et al. 1998; Tsai et al. 2000). On the basis of previous reports that in cardiac tissues the cell size and the distribution of gap junctions play a key role in determination of the anisotropy (Saffitz et al. 1995; Spach and Starmer 1995), further studies comparing the conduction properties between the thoracic veins are required to clarify the functional significance of the findings from the present study.
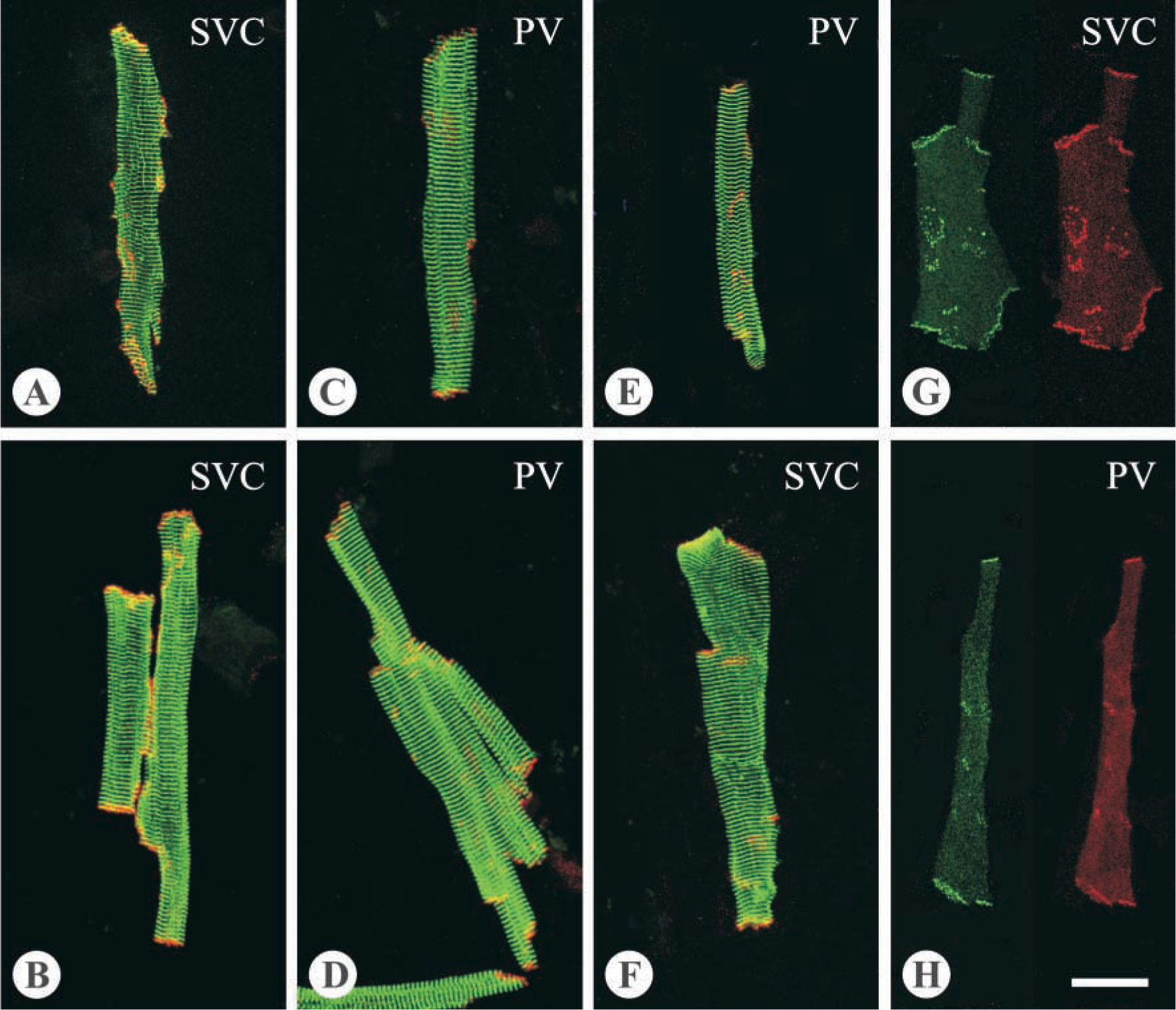
Double-labeling images of cardiomyocytes. (
In conclusion, the canine PV and SVC contain cardiomyocytes with various features. The differential expression of Cx43 gap junctions and differences in cell size in the myocardial sleeve suggest that individual thoracic veins possess distinct electrical conduction properties.
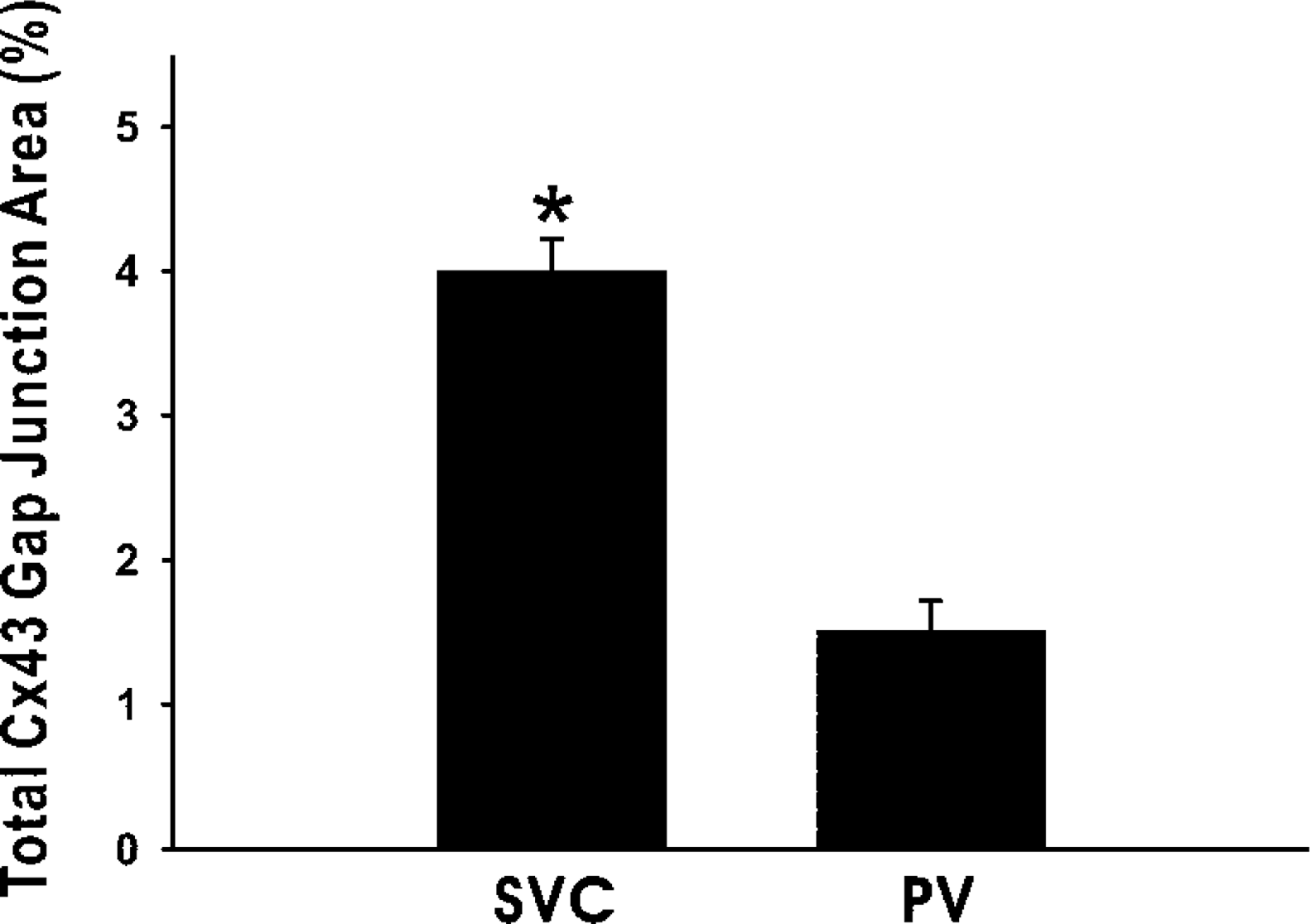
Immunoconfocal analysis of gap junction spots detected by the Cx43 (R530) antibody. Data come from 77 (SVC) and 56 (PV) randomly selected cells. ∗ p<0.01 between the veins.
Footnotes
Acknowledgements
Supported by grants NSC-90–2314-B-195–018 from the National Science Council, Taiwan, and MMH-9112 from the Medical Research Department of the Mackay Memorial Hospital, Taiwan.
We thank Dr Fung J. Lin for assistance with statistics.