
Abstract
The human salivary mucins MG1 and MG2 are well characterized biochemically and functionally. However, there is disagreement regarding their cellular and glandular sources. The aim of this study was to define the localization and distribution of these two mucins in human salivary glands using a postembedding immunogold labeling method. Normal salivary glands obtained at surgery were fixed in 3% paraformaldehyde-0.1% glutaraldehyde and embedded in Lowicryl K4M or LR Gold resin. Thin sections were labeled with rabbit antibodies to MG1 or to an N-terminal synthetic peptide of MG2, followed by gold-labeled goat anti-rabbit IgG. The granules of all mucous cells of the submandibular and sublingual glands were intensely reactive with anti-MG1. No reaction was detected in serous cells. With anti-MG2, the granules of both mucous and serous cells showed reactivity. The labeling was variable in both cell types, with mucous cells exhibiting a stronger reaction in some glands and serous cells in others. In serous granules, the electron-lucent regions were more reactive than the dense cores. Intercalated duct cells near the acini displayed both MG1 and MG2 reactivity in their apical granules. In addition, the basal and lateral membranes of intercalated duct cells were labeled with anti-MG2. These results confirm those of earlier studies on MG1 localization in mucous cells and suggest that MG2 is produced by both mucous and serous cells. They also indicate differences in protein expression patterns among salivary serous cells.
Keywords
S
Human saliva contains two major mucin components, MG1 and MG2. The high molecular weight MG1 consists mainly of the MUC5B gene product (Troxler et al. 1997; Nielsen et al. 1997: Thornton et al. 1999), whereas the low molecular weight MG2 is a product of the MUC7 gene (Bobek et al. 1993). Although there is general agreement that MG1 is produced and localized in mucous cells of the submandibular, sublingual, and some minor salivary glands (Cohen et al. 1990; Audie et al. 1993; Nielsen et al. 1996), this is not the case for MG2. Immunohistochemical (IHC) and in situ hybridization (ISH) studies by Cohen et al. (1991) and Khan et al. (1998) suggest that MG2 also is present in and produced by mucous cells in the submandibular glands. On the other hand, similar studies by Nielsen et al. (1996, 1997) indicate that MG2 is present in a subset of the serous cells of these glands. Using the same antibody and nucleotide probes, Sharma et al. (1998) described a differential localization of MG1 and MG2 in mucous and serous cells, respectively, of human bronchial submucosal glands. No reactivity for MG1 or MG2 was seen in the parotid gland or in salivary gland intercalated, striated, or excretory duct cells in any of these studies.
The purpose of the present study was to further investigate the distribution of these two mucins in human salivary gland tissue using immunoelectron microscopy.
Materials and Methods
Samples of normal submandibular and sublingual glands were obtained from eight consenting male and female patients, aged 44–69 years, undergoing surgery at the Otorhinolaryngology Clinic, University of Cagliari, Cagliari, Italy. All procedures were approved by the Human Experimentation Committee, University of Cagliari. In addition, one sublingual gland was obtained from a 7-year-old boy undergoing sialadenectomy at the Hospital for Sick Children, Toronto, Canada. The procedure was approved by the Hospital Committee for Human Experimentation. The various samples studied are summarized in Table 1.
For light microscopic studies the glands were fixed overnight in 4% paraformaldehyde (Polysciences; Warrington, PA) in 0.1 M sodium cacodylate buffer, pH 7.2, then stored in 1% paraformaldehyde in cacodylate buffer at 4C. The samples were rinsed in 0.1 M cacodylate buffer, dehydrated in cold methanol, embedded in LR Gold resin (Polysciences), and polymerized under UV light (365 nm) at −20C. One sample was dehydrated in ethanol, embedded in LR White resin, and polymerized overnight at 50C. One-micrometer sections were collected on glass slides and incubated for 90 min at room temperature (RT) with rabbit polyclonal antibodies to MG1 (Troxler et al. 1995; Rayment et al. 2000b) diluted 1:1000–1:2000 in 1% bovine serum albumin (BSA)-5% normal goat serum (NGS) in PBS, or to an N-terminal synthetic peptide of MG2 (Liu et al. 1999) diluted 1:500–1:1000 in a mixture of fish gelatin, ovalbumin, and Tween-20 in PBS. Sections incubated with medium devoid of primary antibody or containing nonimmune rabbit serum were used as controls. After incubation the sections were rinsed with PBS and incubated for 60 min at RT with goat anti-rabbit IgG labeled with 5-nm gold particles (Amersham, Arlington Heights, IL) diluted 1:50 in 1% BSA-PBS. After rinsing with PBS and distilled water, silver enhancement (British BioCell; Cardiff, UK) was performed to visualize the bound gold particles. The sections were lightly stained with 1% methylene blue-1% azure II and were observed and photographed in a Leitz Orthoplan microscope.
Tissue processing for electron microscopy employed fixation of small pieces (1 mm3) of the same samples in a mixture of 3% paraformaldehyde and 0.1% glutaraldehyde in 0.1 M cacodylate buffer for 2 hr, after which the glands were stored in 1% paraformaldehyde in cacodylate buffer. After rinsing in buffer, the tissues were dehydrated in cold methanol, embedded in either LR Gold or Lowicryl K4M resin (Polysciences), and polymerized as described above. One gland was fixed in 1% glutaraldehyde and embedded in LR White resin. Ultrathin sections were collected on Formvar-coated nickel grids and treated with either 1% BSA-1% instant milk in PBS or fish gelatin, ovalbumin, and Tween-20 in PBS to block nonspecific binding. The sections were incubated with anti-MG1 diluted 1:100–1:500 in 1% BSA-5% NGS in PBS for 60 min at RT or with anti-MG2 diluted 1:50–1:200 in fish gelatin, ovalbumin, and Tween-20 in PBS overnight at 4C. Omission of the primary antibody or incubation with nonimmune rabbit serum was used as control. After rinsing with PBS, the grids were incubated for 60 min at RT with goat anti-rabbit IgG labeled with 10-nm gold particles (Amersham). The grids were washed with PBS and distilled water, stained with uranyl acetate and lead citrate, and observed and photographed in a JEOL 100CX TEM.
Summary of anti-MG1 and anti-MG2 labeling of human submandibular (SM) and sublingual (SL) glands a
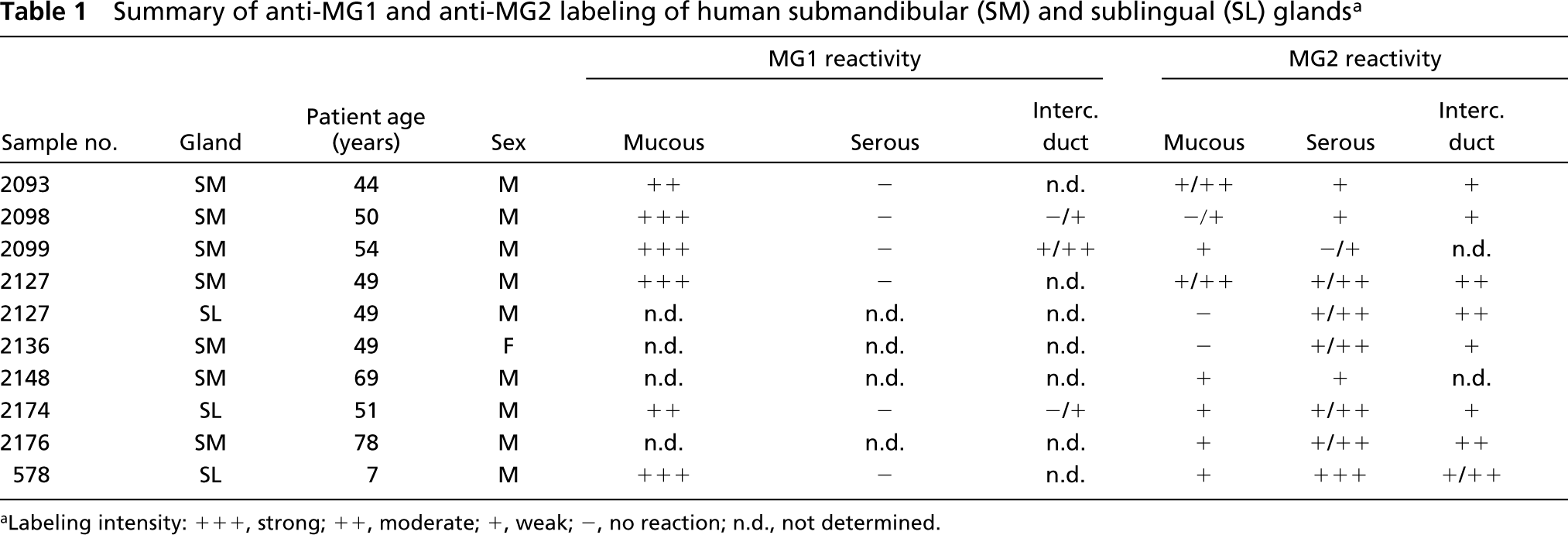
aLabeling intensity: +++, strong; ++, moderate; +, weak; -, no reaction; n.d., not determined.
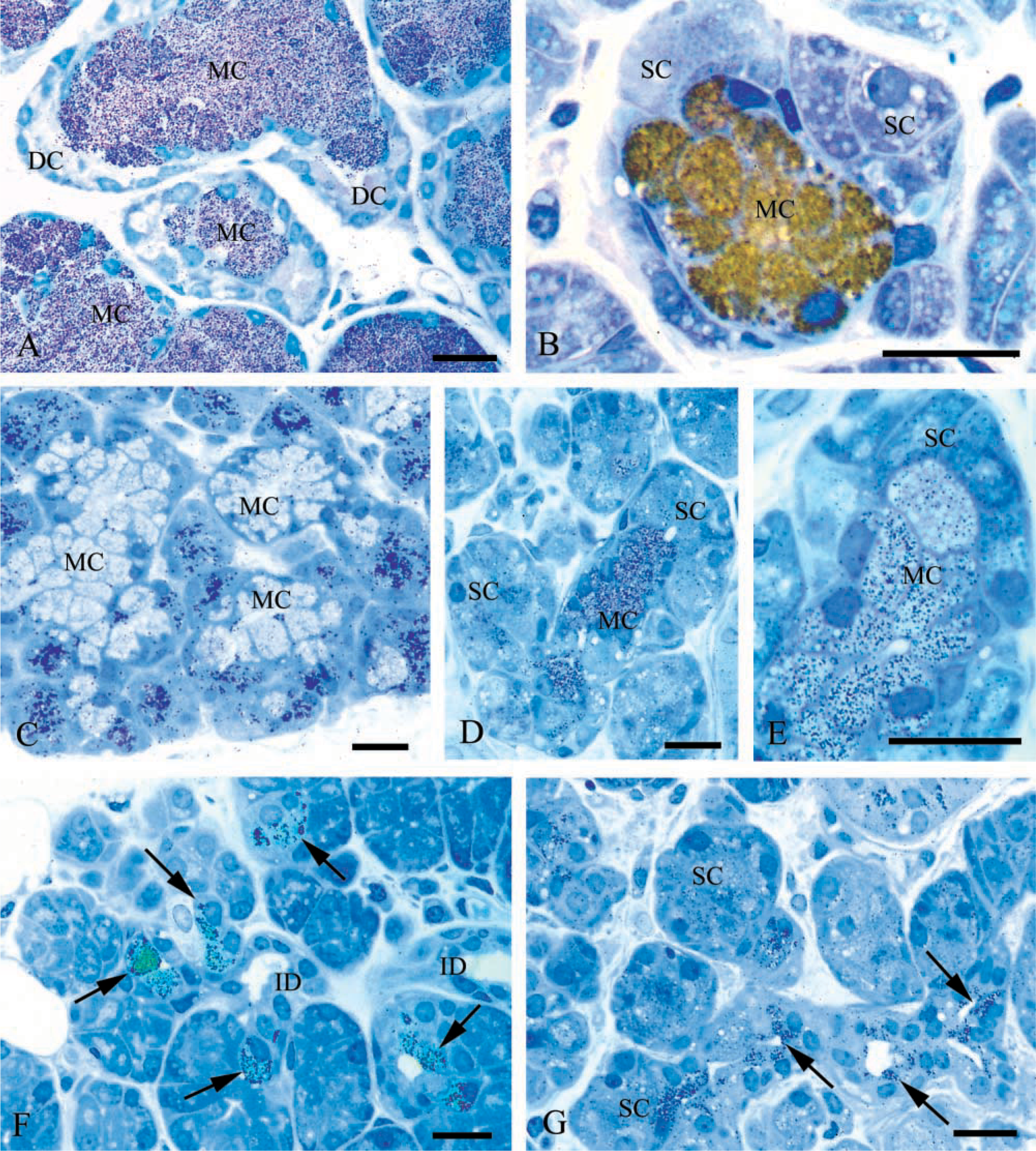
Human salivary glands: immunogold silver staining of 1-μm LR Gold sections with anti-MG1 and anti-MG2 antibodies. (
Results
Specific reactivity for MG1 was detected at the light microscopic level (Figures 1A and 1B) on the granules of all mucous cells of the submandibular and sublingual glands examined at dilutions of the primary antibody of 1:1000–1:2000. The granules of some intercalated duct cells located adjacent to the acini also showed labeling (Figure 1F). No reactivity was found in serous acinar or demilune cells. Striated duct cells also were devoid of labeling.
In the same glands, MG2 labeling exhibited a different distribution. Both mucous and serous components of the tissues showed reactivity at dilutions of the primary antibody of 1:500–1:1000. In most glands examined, the intensity of labeling of serous cells was moderate, whereas that of mucous cells was somewhat weaker. The labeling of both cell types with the anti-MG2 antibody, however, exhibited significant variability. In some glands, mucous cells, especially those of smaller size, labeled strongly (Figures 1D and 1E). In the sublingual gland from the young individual, the serous demilune cells were strongly reactive, whereas the mucous acinar cells exhibited relatively weak labeling (Figure 1C). Intercalated duct cells located near acini exhibited labeling in their apical cytoplasm (Figure 1G). These cells were more consistently reactive than the mucous and serous cells. No labeling was seen in striated duct cells.
In the absence of the primary antibody, or when nonimmune IgG was substituted for the primary antibody, no specific labeling was observed.
Electron microscopic immunogold labeling confirmed and extended the light microscopic observations. The granules of all mucous cells in the submandibular and sublingual glands examined were intensely labeled after incubation with anti-MG1 antibody at dilutions of 1:100–1:500 (Figures 2A–2D). Nonspecific labeling of nuclei, mitochondria, and extracellular spaces was minimal. No labeling was observed in serous cells, and no substantial differences in the distribution and intensity of MG1 reactivity were observed between submandibular and sublingual glands.
The granules of some intercalated duct cells, particularly cells located near the acini, were labeled with the anti-MG1 antibody (Figure 3). The gold particles were present mainly over the electron-lucent portion of these granules. The labeling density of the intercalated duct cell granules appeared to be less than that of granules in the mucous acinar cells. In a few cases, gold particles were present along the luminal surface of the duct cells (Figure 3, inset), suggesting that this surface may be coated with MG1.
Ultrathin sections of the submandibular and sublingual glands incubated with the anti-MG2 antibody (1:50–1:200) exhibited specific reactivity in both mucous and serous secretory granules (Figures 4 and 5). As observed by light microscopy, the distribution and intensity of labeling with the anti-MG2 antibody reactivity showed greater variability than that observed for MG1. In the samples examined, the granules of most mucous cells had low levels of labeling, and many serous cells had unreactive secretory granules (Figure 5B). Only a few serous acinar cells displayed labeling in the lucent halo of their granules. In one of the submandibular glands and one of the sublingual glands (Figure 4) studied, a more intense and uniform labeling was found in the electron-lucent halo of the serous granules, and mucous acinar cells in these glands were either unreactive or only weakly labeled. The dense cores of the serous granules were almost unreactive. Occasionally they were labeled to a similar extent as in sections incubated with appropriate dilutions of normal rabbit serum, suggesting nonspecific binding of the primary antibody to the electron-dense cores of the serous granules. The endoplasmic reticulum and Golgi apparatus of the mucous and serous cells consistently were unlabeled.
Intercalated duct cells located near acini also displayed MG2 reactivity in secretory granules and vesicles located in their apical cytoplasm (Figures 6A–6C). As noted for intercalated duct cells labeled with anti-MG1 antibody, gold particles representing MG2 reactivity were restricted mainly to the electron-lucent portion of the granules. The labeling density of the intercalated duct cell granules was similar to that seen in granules of serous cells. In contrast, the labeling density of the small vesicles appeared to be somewhat greater. In addition, the Golgi apparatus of these cells was labeled (Figure 6E) and reactivity was present along the basal and lateral plasma membranes, especially in regions with prominent membrane folding (Figure 6D).
Discussion
MG1 was present in all mucous cells of the submandibular and sublingual glands. No reactivity for MG1 was observed in serous cells. This pattern of reactivity is similar to the distribution of MG1 described in a previous IHC study (Nielsen et al. 1996). Cohen et al. (1990) also found that MG1 was restricted to mucous cells, but noted that some mucous cells were unreactive. A possible explanation for this difference is that the antibody used by Cohen et al. (1990) was to a carbohydrate epitope, whereas the antibody used by Nielsen et al. (1996) was a monoclonal antibody directed against an epitope on the apomucin. The antibody used in the present study is a polyclonal antibody that recognizes both polypeptide and carbohydrate epitopes (R.F. Troxler and G.D. Offner, unpublished observations). Heterogeneity among mucous cells with regard to the carbohydrate structures present on mucins has been described in other studies (Laden et al. 1984). In addition, ISH with an MUC5B-specific probe revealed MG1 transcripts in all mucous cells of the submandibular and sublingual glands (Nielsen et al. 1997).
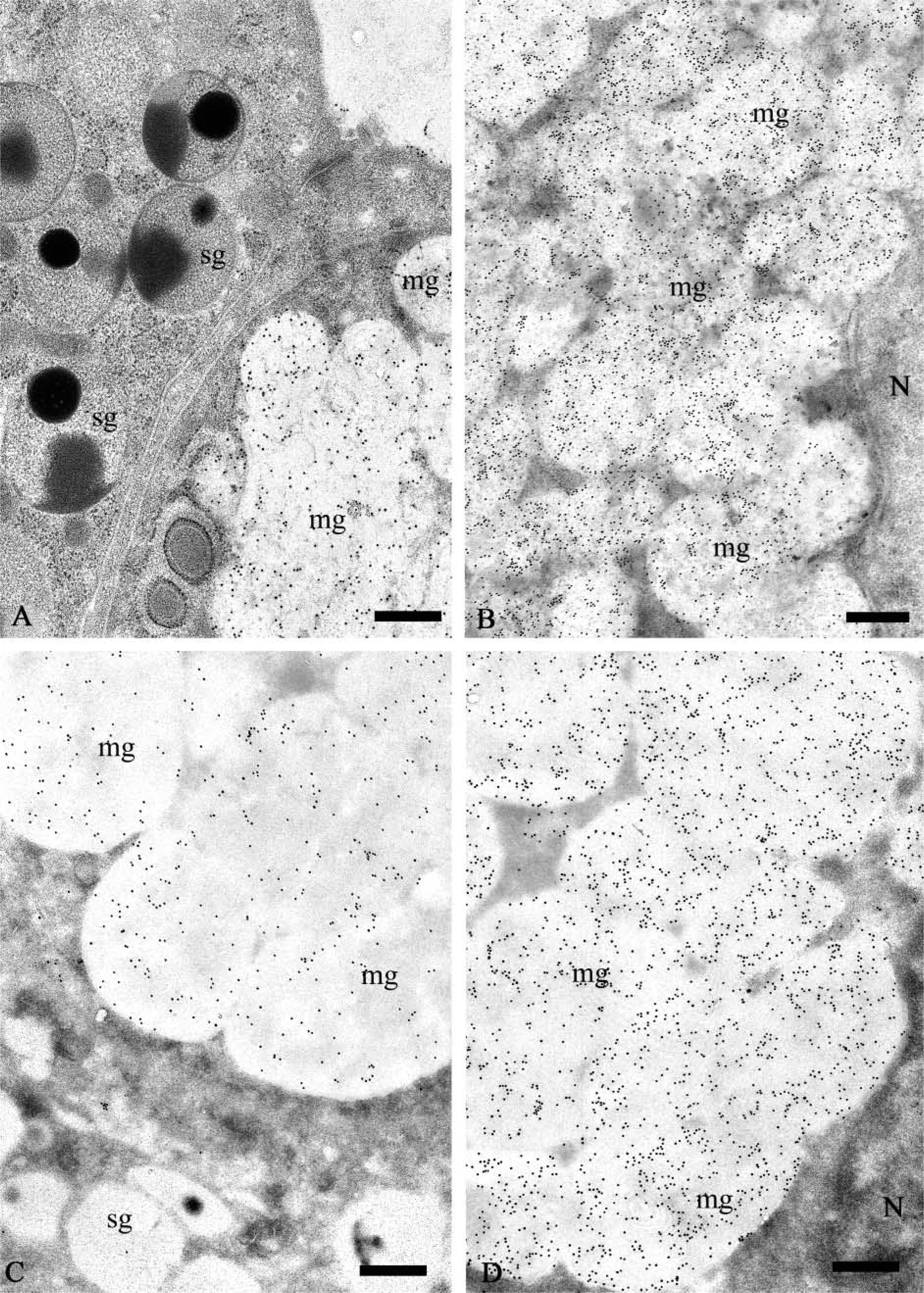
Human salivary glands: EM immunogold labeling with anti-MG1 antibody. (
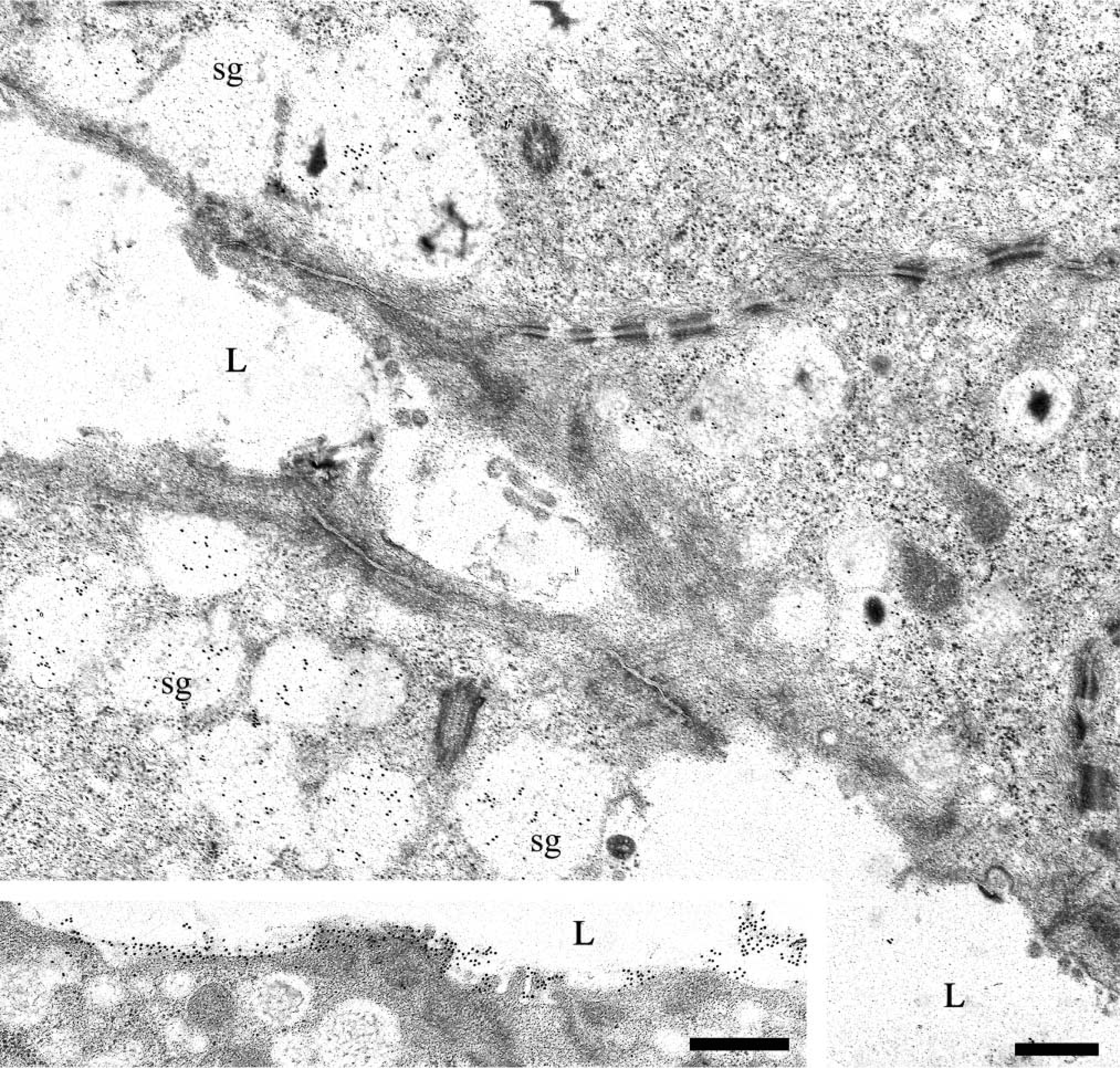
Human submandibular gland: EM immunogold labeling with anti-MG1 antibody. Secretory granules (sg) of intercalated duct cells show reactivity. (
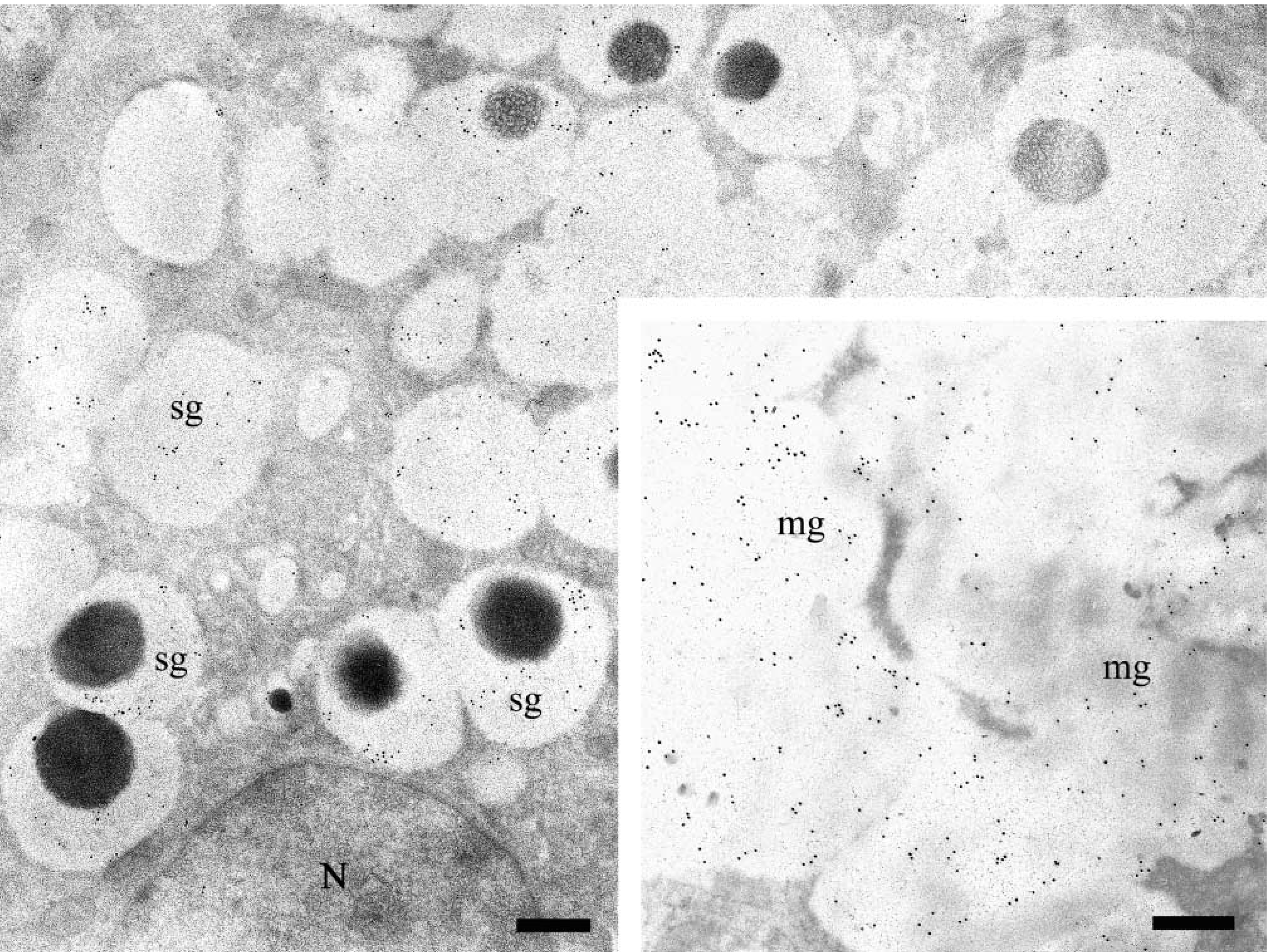
Human sublingual gland: EM immunogold labeling with anti-MG2 antibody. Serous demilune cells show more uniform reactivity in their secretory granules (sg) than that detected on submandibular serous cells. (
Although labeling of the mucous granules with anti-MG1 was intense, little or no labeling of other mucous cell compartments was present. There are several possible explanations for this observation. The concentration of MG1 in the lumina of the endoplasmic reticulum and Golgi saccules may be below the limit of detection allowed by the tissue preparation and labeling methods employed. The association of the apomucin with other molecules in these compartments, e.g., molecular chaperones in the endoplasmic reticulum or glycosyltransferases in the endoplasmic reticulum and Golgi apparatus, might interfere with antibody labeling. Possible differences in conformation of the mucin polypeptide related to the extent of glycosylation could alter the reactivity of various epitopes with the antibody.
Labeling for MG2 was more variable than for MG1. In five of six submandibular glands and two of three sublingual glands examined, mucous cells exhibited weak to moderate labeling. Labeling of serous granules in the submandibular gland was highly variable, even within the same serous cell. Serous cells of the sublingual gland were more consistently labeled for MG2. In one submandibular and one sublingual gland the mucous cells exhibited very little reactivity, whereas the serous cells exhibited the most prominent labeling. Therefore, the previous discrepancies reported for MG2 localization may reflect variability in the level of expression of MG2 in the mucous and serous cells of different individuals. It is also possible that the heterogeneous labeling for MG2 observed in the present study may be due to restricted access of the antibody as a result of the presence of different carbohydrate moieties on the mucin molecules.
Recently, the presence of other mucins has been described in the salivary glands. MUC1 (Newland et al. 1997; Sengupta et al. 2001; Liu et al. 2002) and MUC4 (Liu et al. 2002), membrane-associated mucins, have been localized to the ducts and some serous cells of human major and minor salivary glands. Moreover, the transmembrane component of rMuc4 is reported to be present in serous cells and duct cells of the major rat salivary glands (Li et al. 2001). The presence of MG1 and MG2 in the intercalated duct cells, as described in this study, suggests that these cells express several types of mucins. The granules of the duct cells were labeled for both MG1 and MG2. The presence of MG2 reactivity in apical vesicles could indicate an alternative route for secretion or could represent endocytosis of MG2 from the duct lumen. Labeling for MG1 was occasionally present along the apical membrane of the duct cells, presumably due to binding of secreted mucin. Membrane-associated and adherent mucins at the luminal surface of the duct cells may function to maintain duct patency by preventing adherence of the luminal membranes of opposing cells (Hilkens et al. 1992; Sengupta et al. 2001). Liu et al. (2002) have suggested that the cytoplasmic domain of membrane-associated mucins may be involved in signal transduction. Interestingly, labeling for MG2 also was consistently observed on the basolateral membranes of these cells. The significance of this localization remains to be determined.
Secretory granules of serous cells of human salivary glands typically exhibit a bi- or tripartite ultrastructure, with an electron-dense core and/or electron-dense crescents in an electron-lucent matrix (Tandler and Erlandson 1972; Riva et al. 1974). Granules with a similar substructure are present in intercalated duct cells located adjacent to acinar cells (Lantini et al. 1988). It is generally believed that these substructural patterns reflect the differential distribution of various granule components. Previous studies of the distribution of amylase and agglutinin (Takano et al. 1991) and the H and Lewis-b blood group antigens (Cossu et al. 1990,1994) in human salivary glands support this hypothesis. The localization of MG2 to the electron-lucent portion of the serous granules and MG1 and MG2 to the electron-lucent portion of intercalated duct cell granules is consistent with these earlier observations.
The presence of MG2 in submandibular and sublingual serous cells, as described in this study and by Nielsen et al. (1996, 1997), but not in parotid serous cells, indicates that protein expression patterns differ among the serous cells of human salivary glands. A similar situation exists for the salivary glands of other species (Hand et al. 1999). The expression of MG2 and possibly other mucins by salivary gland serous cells contributes to a blurring of the classical histological distinction between serous and mucous cells. It also bears on the debate, based largely on histochemical staining for glycoconjugates, over the use of the terms “serous” or “seromucous” to describe these cells (Pinkstaff 1993). Additional studies that further define the molecular phenotypes of the secretory cells of human salivary glands may help to settle these issues.

Human submandibular gland: EM immunogold labeling with anti-MG2 antibody. (
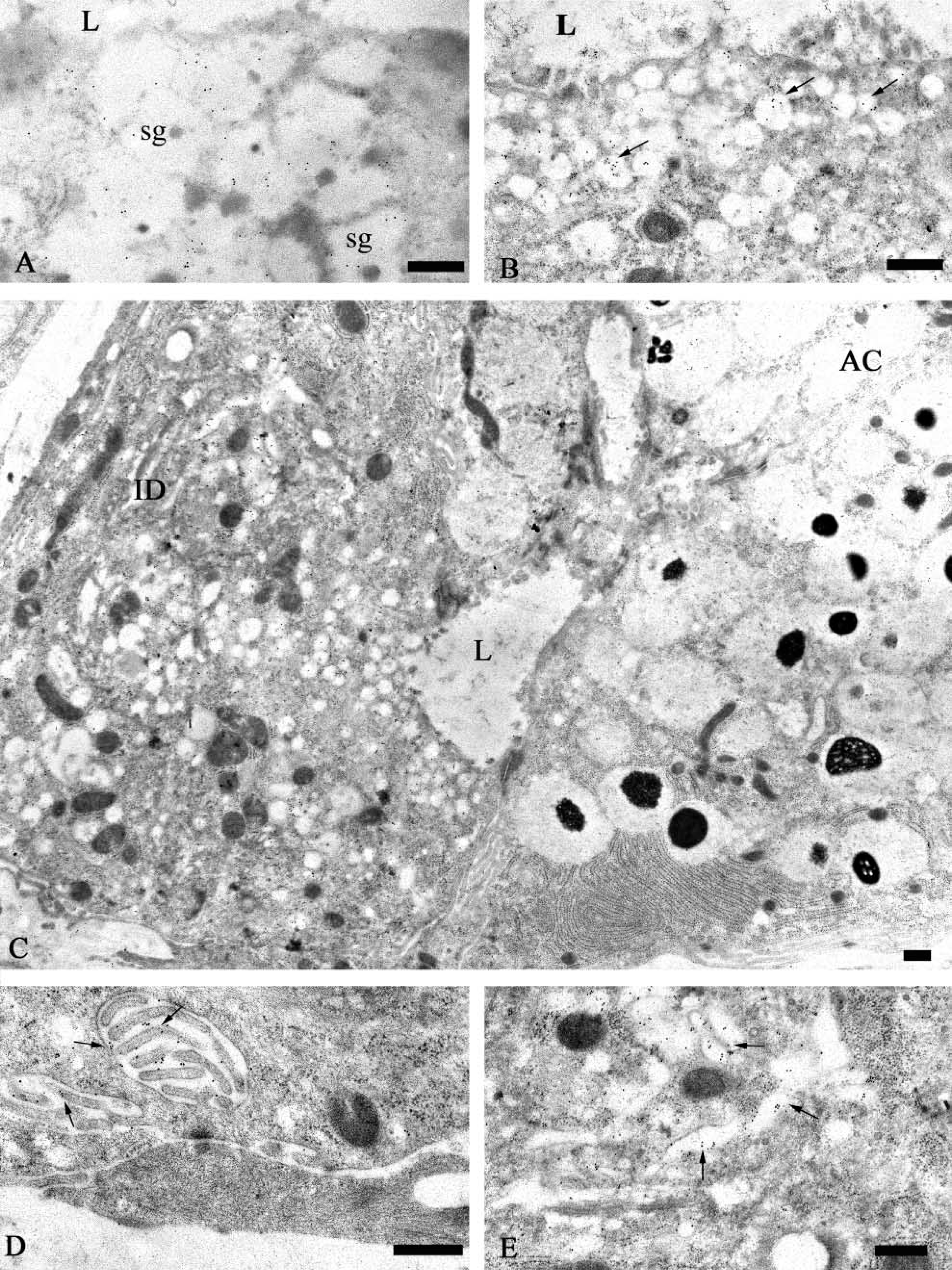
Human submandibular gland: EM immunogold labeling with anti-MG2 antibody. (
Footnotes
Acknowledgements
Supported by a fellowship from the Universita Degli Studi di Cagliari (MP), the University of Connecticut Health Center, and by NIH grants DE11691, DE07652, and DK44619.