
Abstract
P-Glycoprotein (P-gp) is an ATP-dependent efflux transporter that extrudes non-polar molecules, including cytotoxic substances and drugs, from the cells. It was initially found in cancer cells and then was shown to be a normal component of complex transport systems working at the blood-brain barrier (BBB). Previous studies have demonstrated that, in the brain, P-gp is localized on the luminal plasmalemma of BBB endothelial cells and that it may interact with the caveolar compartment of these cells. The aim of this study was to identify the site of cellular expression of P-gp in human brain in situ and to morphologically determine whether an association may exist between P-gp and caveolin-1, a structural and functional protein of the caveolar frame. The study was carried out on human cerebral cortex by immunoconfocal microscopy with antibodies to both P-gp and caveolin-1. The results show that P-gp marks the microvessels of the cortex and that the transporter is localized in the luminal endothelial compartment, where it co-localizes with caveolin-1. The demonstration of this co-localization of P-gp with caveolin-1 contributes a morphological backing to biochemical studies on P-gp/caveolin-1 relationships and leads us to suggest that interactions between these molecules may occur at the BBB endothelia.
Keywords
I
The precise function of P-gp in normal tissues is not completely understood. In somatic microvessels, in which P-gp was found on both luminal and abluminal membranes of the endothelial cells, the transporter could actively regulate the efflux of lipophilic substances from the endothelial cytoplasm on both blood and tissue fronts. On the contrary, in the BBB microvessels, where the highest levels of P-gp have been detected, the transporter was described only at the luminal endothelial membrane. This selective one-front localization suggests that P-gp plays a barrier protective role by extruding cytotoxic substances and drugs from the endothelial cells back into the bloodstream (Jetté et al. 1993; Stewart et al. 1996; Beaulieu et al. 1997; Demeule et al. 2001). This P-gp defense activity is likely to acquire great relevance in human brain, in which the levels of P-gp are three- to fivefold higher than in murine and bovine brain (Jetté et al. 1993). Despite the volume of data available on P-gp, some aspects, such as the cellular site of expression and regulation of its activity, have not yet been clarified. Although it is generally accepted that in the brain P-gp is expressed on the luminal membranes of the microvascular endothelial cells, studies in human brain have identified the site of expression of P-gp in perivascular astrocytes (Pardridge et al. 1997; Golden and Pardridge 1999). Moreover, biochemical studies have indicated that physical and functional interactions may take place in the caveolar microdomain of the cell plasma membranes between P-gp and caveolin-1, an integral protein of the caveolar frame (Demeule et al. 2000). With the aim of gaining further insight into P-gp localization in human brain, this in situ study was carried out on cerebral cortex by immunofluorescence laser scanning confocal microscopy using a monoclonal antibody to P-gp (the JSB-1 clone). In addition, double immunolabeling was carried out with antibodies to P-gp and caveolin-1 to ascertain whether there is any association between these molecules in the caveolar cell compartment.
Materials and Methods
This study included 10 samples of brain tissues, measuring 4–6 mm in maximal diameter, obtained from 10 patients (six men and four women; age range 50–65 years, median 58 years) undergoing tumor resection for high grade (III-IV) gliomas with concomitant mapping of tumor-free margins. The latter procedure consisted of intraoperative examination of hematoxylin-eosin (H&E)-stained frozen tissue sections obtained from half of each of the above 10 samples that confirmed absence of tumor invasion. The other halves of the tumor-free samples were immersed in a fixative mixture consisting of 2% paraformaldehyde plus 0.2% glutaraldehyde in PBS for 2 hr at 4C. Sections (25-μm) were prepared using a vibrating microtome (Leica; Milton Keynes, UK). Fluorescence single and double immunolabeling was carried out by a second-layer indirect method on free-floating sections. The primary antibodies were a mouse monoclonal antibody (MAb) raised against the cytoplasmic epitope of the plasma membrane-associated 170–180-kD glycoprotein (1:20 dilution; clone JSB-1, IgG1 subclass; Novocastra Laboratories, Newcastle upon Tyne, UK) and a rabbit polyclonal antibody raised against the N-terminus of caveolin-1 of human origin (1:100 dilution; Santa Cruz Biotechnology, Santa Cruz, CA). In addition, to further check the normal nature of the non-neoplastic brain tissue samples, the sections were subjected to double labeling for the glucose transporter isoform 1 (GLUT-1) as a marker for BBB endothelial cells and for the glial fibrillary acidic protein (GFAP) as a marker for astroglial cells, utilizing a rabbit polyclonal antibody anti-GLUT-1 (1:500 dilution; Chemicon International, Temecula, CA) and a mouse MAb anti-GFAP (1:100 dilution; Novocastra Laboratories). The immunostains were performed in parallel with those for P-gp/caveolin-1.
The sections were immersed and gently agitated at room temperature as follows: (a) 30 min in blocking buffer (BB: PBS, 1% BSA, 2% FCS); (b) 30 min in 0.5% Triton X-100 in PBS; (c) 1 hr in either one or a mixture of the two primary antibodies (double labeling) suitably diluted in BB; (d) 1 hr in the appropriate secondary antibody/ies, Alexa Fluor 488 goat anti-mouse IgG (Fab'2-conjugated fragment; Molecular Probes Europe, Leiden, The Netherlands; for P-gp and GFAP), Alexa Fluor 568 goat anti-rabbit IgG conjugate (Molecular Probes; for caveolin-1 and GLUT-1) both diluted 1:200 in BB. The sections were washed three times for 10 min in PBS between each step. Nuclear counterstaining was performed by incubations either in RNase diluted 5 μl/ml in PBS (30 min at 37C) and then propidium iodide (Molecular Probes) diluted 1 μl/ml in PBS, or by TO-PRO-3 iodide (Molecular Probes) diluted 1:10 in PBS and added to the third wash after the conjugate antibodies. The sections were then transferred onto subbed slides (1% gelatin plus 1% formalin in distilled water), carefully drained, coverslipped in Vectashield anti-fade mounting medium (Vector Laboratories; Burlingame, CA), and sealed with nail polish. To fully preserve the fluorescence signal, no pretreatments were applied to avoid autofluorescence of both lipofuscin pigments and lipofuscin-like substances in neurons and perivascular cells (Schnell et al. 1999).
The control sections were prepared (a) by substituting the primary antibodies with BB, (b) by substituting anti-P-gp with an inappropriate MAb of identical subclass and anti-caveolin-1 with normal rabbit serum, both at the same working dilution, (c) preadsorbing the primary antibody anti-caveolin-1 with an excess of the pure antigen available from the supplier, or (d) mismatching the secondary antibodies. The results of these negative controls were consistent with the expected results.
The sections were viewed under the Leica TCS SP confocal laser scanning microscope using X40 and X63 oil-immersion objective lenses with either X1 or X2 zoom factors. On the double-immunolabeled sections, a sequential scan procedure was applied during image acquisition of the two fluorophores, Alexa Fluor 488 (excitation at 488 nm and detection range 500–535 nm; green fluorescence) and Alexa Fluor 568 (excitation with 568 nm and detection range 580–620 nm; red fluorescence). Confocal images were taken at 0.5-μm intervals through the z-axis of the section covering in total 15–20 μm in depth. Images from individual optical sections and multiple serial optical sections were analyzed, recorded digitally, and stored as TIFF files in Adobe Photoshop software (Adobe Systems; San Jose, CA).
All the investigation procedures on human tissues were carried out in accordance with the local institutional ethical committee policies.
Results
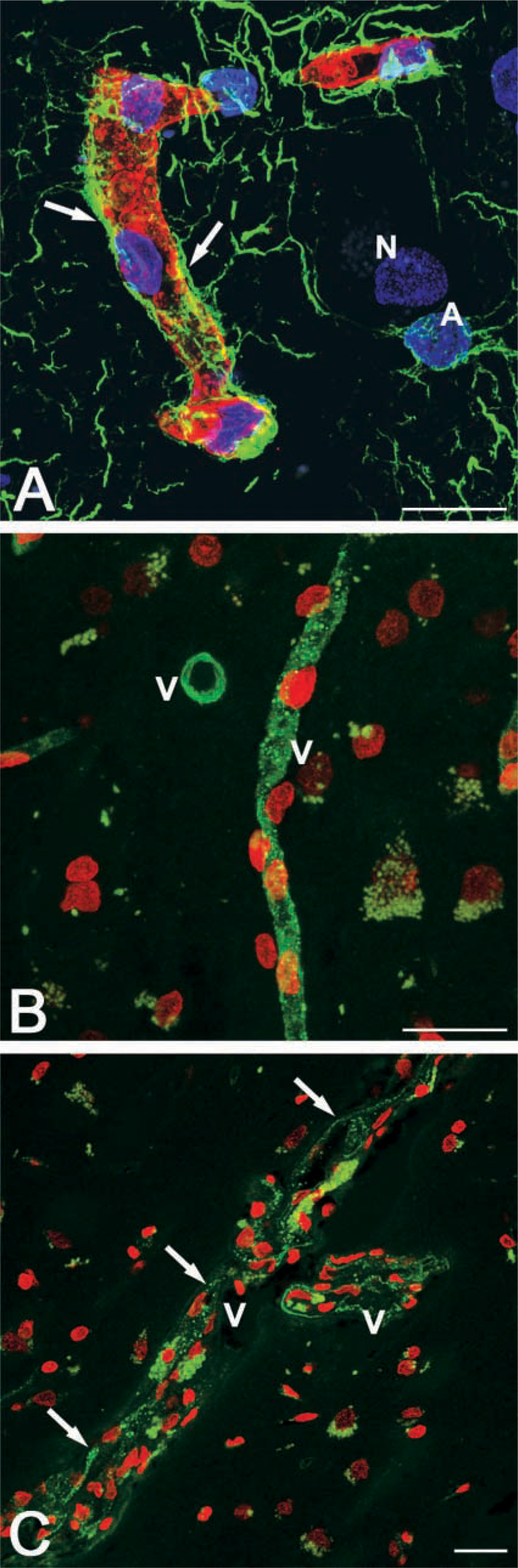
Examination of the sections double immunolabeled for GLUT-1 and GFAP confirms the normal morphological features of the tissue. GLUT-1 is expressed by the endothelial cells of the cortex microvessels and GFAP marks astrocyte processes scattered in the neuropil and regularly distributed on the microvessel wall (Figure 1A).
The cortex microvessels are heavily stained by P-gp immunolabeling, showing a distribution that appears to be independent of the lamination of the cortex. P-gp preferentially stains the cortex microvessels, whereas it appears weaker or undetectable on the larger vessels. It does not stain neurons and glial cells in the neuropil. In neurons, only the autofluorescence of lipofuscin is detectable and is always easily distinguishable from the bright-green P-gp-specific staining (Figure 1B). On reconstructed microvessels obtained with multiple optical planes, the P-gp labeling appears finely punctate and evenly distributed all along the vascular walls, with many scattered, more evident fluorescent puncta (Figure 1B). On longitudinal- and cross-sectioned microvessels, the P-gp labeling reveals the entire vessel profile and is seen to correspond to the thin endothelial cells that line the vessel walls (Figure 1C). Perivascular phagocytic cells containing autofluorescent lipofuscin-like granules are a constant feature of these vascular fields (Figure 1C). On the double P-gp/caveolin-1-immunolabeled sections (Figures 2A-2L), observation at higher magnification on serial optical planes of cross-sectioned microvessels confirms that the expression of P-gp corresponds to the endothelial cells and also reveals that the transporter is localized in their luminal compartment (Figures 2A and 2F). In the vascular wall, weaker reactivity is also detectable in cells in close contact with the endothelium and recognizable as pericytes owing to their position and nuclear shape (Figure 2F). Unlike P-gp, caveolin-1 stains the entire thickness of the endothelium from the luminal to the abluminal side, with a finely punctate pattern in the endothelial luminal compartment and larger fluorescent puncta in the abluminal one (Figures 2B and 2G). Comparing the single channel signal of P-gp and caveolin-1, green and red fluorescence, respectively, it is evident that both proteins are present in the luminal compartment of the endothelial cells (Figures 2A, 2B, 2F, and 2G). Nevertheless, only on the merged images, obtained by superimposing the two fluorescent signals, is the exact extent of P-gp/caveolin-1 co-localization revealed (Figures 2C-2E and 2H-2L; yellow fluorescence). More frequently, P-gp and caveolin-1 appear to largely co-localize in the luminal compartment of the endothelial cells (Figures 2C-2E), although elsewhere the fluorescent signals do not appear to overlap completely and co-localization is detectable only at the boundary between the luminal and abluminal endothelial cell compartments (Figures 2H-2L).
Double (GLUT-1/GFAP;
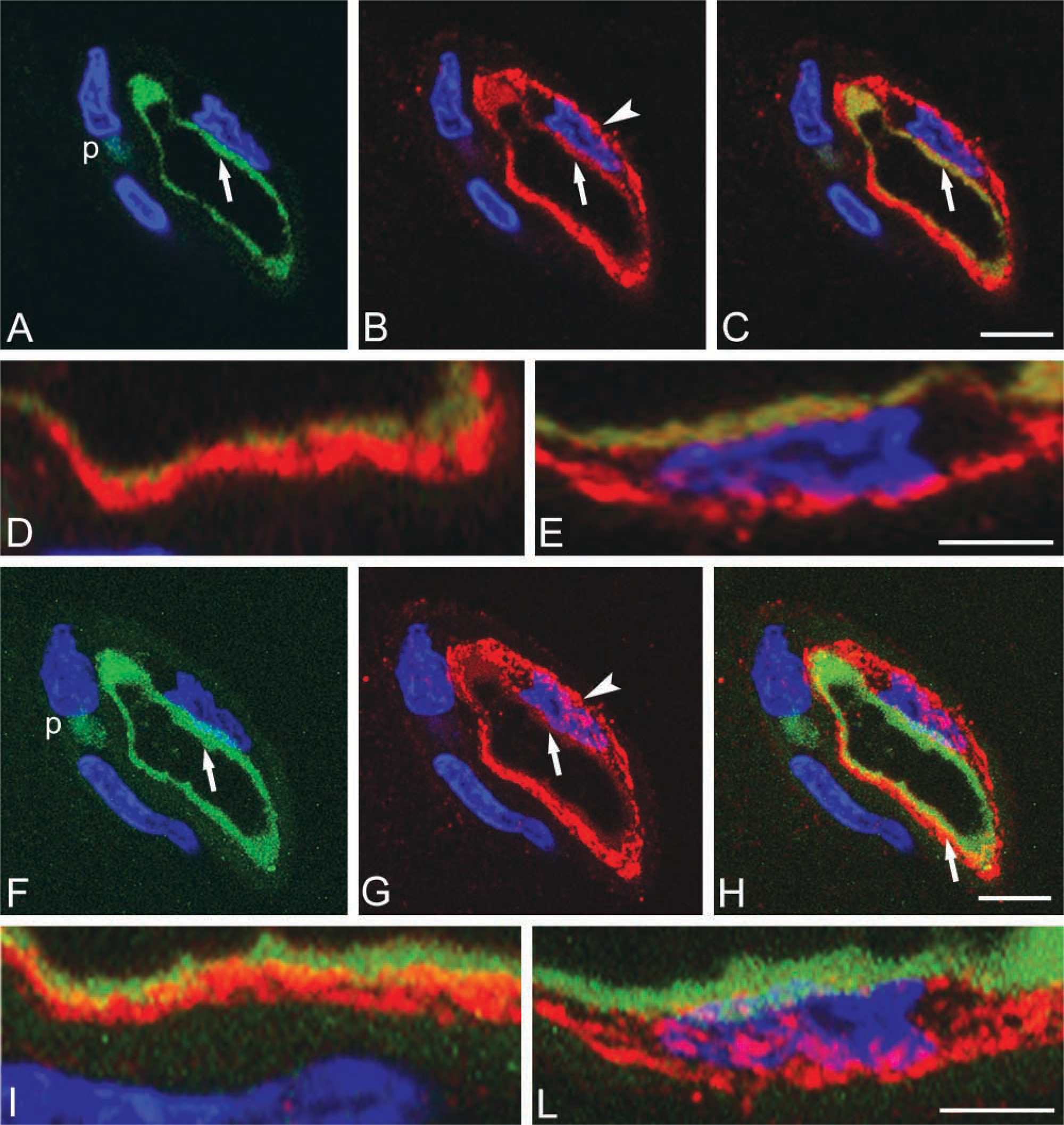
Double immunolabeling for P-gp (green) and caveolin-1 (red) of human cerebral cortex. (
Discussion
A large body of work has analyzed P-gp expression, substrates, and activities in the CNS, obtaining significant information thanks to many different experimental approaches, such as cultures of cerebral endothelial cells, isolated brain microvessels, and the P-gp knockout mouse (Greenwood 1992; Schinkel et al. 1994; Begley et al. 1996; Miller et al. 2000; Megard et al. 2002). In the specific field of the human BBB, our study contributes to the knowledge on P-gp cellular localization and molecular interactions in human cerebral cortex in situ. The results show that the primary site of P-gp expression is the endothelial cells lining the small vessels of the cortex, in which the transporter appears concentrated within the luminal cellular compartment. Weaker but clearly detectable P-gp reactivity is also revealed in the microvascular pericytes that closely envelop the endothelial cells remaining enclosed within their basal lamina. These data confirm an effective role of endothelial P-gp as a “gatekeeper” in the BBB that limits the brain influx of drugs (Schinkel 1999) and indicate the pericytes as a possible second line of defense at BBB sites. The discrepancy with previous results obtained on unfixed human brain, showing the presence of P-gp immunoreactivity in the end-feet of perivascular astrocytes (Golden and Pardridge 1999), could be explained by differences in histological and immunocytochemical procedures, and/or by the mouse MAb clone. In this study, the JSB-1 clone used on lightly fixed free-floating sections, without tissue embedding or freezing, together with the high power of confocal microscopy, yielded good immunoreactivity and preservation of both antigenicity and cell structure, as well as enhanced resolution of morphological details.
As to the mechanisms of action and regulation of the P-gp efflux pump in resistant and normal cells, previous biochemical studies demonstrated that drugs are sequestered in P-gp-containing vesicles and that the caveolar number and caveolin-1 levels parallel the expression of P-gp (Shapiro et al. 1998; Lavie et al. 1999). Furthermore, by immunoprecipitation experiments caveolin-1 has been demonstrated to physically interact with P-gp in normal brain endothelia (Demeule et al. 2000). It is noteworthy that far from being merely a structural protein of the caveolar frame, caveolin-1 is a multifunctional protein able to compartmentalize and to modulate and integrate the activities of a variety of molecules involved in vascular functions (Parton 1996; Feng et al. 1999; Schlegel and Lisanti 2001; Xu et al. 2001). The data reported in this study on the co-localization of P-gp with caveolin-1 provide the first morphological evidence of the association between P-gp and caveolin-1 in normal brain endothelia and highlight the dynamic nature of this interaction. With regard to P-gp and caveolin-1 distribution and colocalization, two major points have to be considered. Immunolabeling for caveolin-1 reveals fine fluorescent puncta in the luminal endothelial compartment and intensely fluorescent larger puncta and micropatches concentrated in the abluminal cell compartment. This pattern confirms the finding that a different subcellular distribution and a distinct state of aggregation of caveolae and caveolae-associated vesicles characterize the opposite endothelial compartments (Tani et al. 1977). A study carried out on microvessels isolated from rat brain and submitted to subcellular fractionation (Demeule et al. 2000) demonstrated that only a population of P-gp molecules is associated with caveolin-1, and has suggested that different cellular events may modify the localization of P-gp and caveolin-1 and lead to a redistribution of both the proteins. The view of highly dynamic interactions between the caveolar microdomain and P-gp (Demeule et al. 2000; Ferté 2000) is consistent with the uneven distribution of P-gp/caveolin-1 co-localization observed in human brain endothelial cells, and this further corroborates the evidence for the existence of functional relations between P-gp and caveolin-1.
Footnotes
Acknowledgements
Supported by grants from the Consiglio Nazionale delle Ricerche (to L.R.) and the Ministero dell'Istruzione, dell'Università e della Ricerca (to D.V.).
We thank Ms M.V.C. Pragnell, BA, for linguistic help and Ms M. Ambrosi for excellent technical assistance.