
Abstract
We examined the effects of specific inhibitors, brefeldin A (BFA) and okadaic acid (OA), on the ultrastructural organization of the Golgi apparatus and distributions of amylase, Golgi-associated proteins, and cathepsin D in the rat parotid acinar cells. BFA induced a rapid regression of the Golgi stack into rudimentary Golgi clusters composed of tubulovesicules, in parallel with a redistribution of the Golgi-resident proteins and a coat protein (β-COP) into the region of the rough endoplasmic reticulum (rER) or cytosol. The rapid disruption of the Golgi stack could also be induced by the effect of OA. However, redistribution of the Golgi proteins in rER or cytosol could not be observed and β-COP was not dispersed but was retained on the rudimentary Golgi apparatus. These findings suggested that the mechanism of OA in inducing degeneration of the Golgi stack was markedly different from that of BFA. In addition, missorting of amylase, a Golgi protein, and cathepsin D into incorrect transport pathways is apparent in the course of the disruption of the Golgi stack by OA. These Golgi-disrupting effects are reversible and the reconstruction of the stacked structure of the Golgi apparatus started immediately after the removal of inhibitors. In the recovery processes, missorting was also observed until the integrated structure of the Golgi apparatus was completely reconstructed. This suggested that the integrated structure of the Golgi apparatus was quite necessary for the occurrence of normal secretory events, including proper sorting of molecules.
M
The fungal metabolite BFA has been known to induce rapid and reversible disassembly of the Golgi stack into tubules and vesicles, resulting in the redistribution of Golgi-resident enzymes into the endoplasmic reticulum in a reversible manner (Misumi et al. 1986; Yamashina et al. 1990; Alcalde et al. 1992; Hendricks et al. 1992; Hidalgo et al. 1992; Klausner et al. 1992). The target of the initial BFA action was indicated to be ADP-ribosylation factor-guanine nucleotide exchange factor (ARF-GEF) to disturb attachment of coatomers to the donor membrane (Morinaga et al. 1996). This effect causes inhibition of budding formation, redistribution of Golgi protein into the ER and, finally, disorganization of Golgi stack.
OA, a monocarboxylic polyether isolated from marine sponges, is a potent inhibitor of the protein phosphatases 1 and 2a and has been used to investigate the role of dephosphorylation in various regulated cell events (Takai et al. 1987; Yamashita et al. 1990; Takuma and Ichida 1991). It has also been shown that incubation of interphase cultured cells with OA induces disassembly of the Golgi stack into clusters of small vesicles, in a process analogous to that which occurs during the mitotic process. The effects of OA on cells are quite reversible, and the reassembly of the Golgi stack that occurs on OA removal is reminiscent of that which occurs during telophase in mitosis (Lucocq et al. 1991; Thyberg and Moskalewski 1992). However, the mechanism of how OA disrupts the structure of the Golgi stack is still unclear. Mitotic Golgi fragmentation is dependent on GM130 phosphorylation on serine 25 and dissociation of p115 from the Golgi membrane, whereas OA induces Golgi fragmentation without GM130 phosphorylation or p115 dissociation (Lowe et al. 2000). Therefore, we also investigated the dynamics of Golgi-related proteins under the effect of OA to clarify the mechanism of the Golgi-disrupting action by OA. We found marked differences between BFA and OA in the distributional change of a Golgi-resident enzyme and a coat protein.
In addition, the disassembly and reassembly processes in Golgi apparatus induced by BFA or OA were found to be a quite useful model to analyze not only the mechanism for the establishment and maintenance of the integrated stacked structure but also the relationship between the structure and function of the Golgi apparatus (Yamashina 1995). Immunogold electron microscopic findings for the Golgi disassembly and reassembly processes in parotid acinar cells indicated that the integrated structure was required for the Golgi apparatus to express normal sorting machinery on the stack.
Detailed investigations at the ultrastructural level in highly specialized and parenchymal cells have been limited, mainly because of the difficulties in preservation of antigenicity and this organelle's complicated morphology. We conducted an examination on the importance of the stacked structure and functions of the Golgi apparatus in the rat parotid acinar cell, because this cell type has the great advantage of containing highly developed and organized secretory components that are easily defined morphologically (Amsterdam et al. 1969), and its excretion can be controlled by isoproterenol, an adrenergic β-agonist (Byrt 1966; Simson et al. 1974). As the existing mature secretory granules are entirely released from the cell immediately after isoproterenol injection, granules observed in the stimulated cell can be identified as newly formed. The most abundant protein in the secretory pathway, amylase, can be detected by immunogold electron microscopy with high sensitivity (Tanaka et al. 1981). Therefore, the traffic of a newly synthesized secretory material can be recognized morphologically and immunocytochemically during the process of reconstitution of secretory granules without pulse-labeling or inhibition of protein synthesis. Because the flow of secretory materials is accelerated by secretory stimulation and is believed to be physiologically natural for the cell, this experimental system is therefore quite useful for analysis of intracellular transport in conjunction with dynamics of organelles.
Materials and Methods
Materials
The monoclonal antibody (MAb) raised from the Golgi fraction of guinea pig pancreas, GF-1 (mouse IgM), has been described previously (Tamaki et al. 1992; Tamaki and Yamashina 1994). The rabbit anti-human α-amylase anti-serum, the rabbit anti-calf brain cathepsin D antiserum, anti-β-COP MAb (mouse IgG1), and BFA were purchased from Sigma (St Louis, MO). Anti-mannosidase II MAb (mouse IgG2b) was obtained from Berkeley Antibody (Richmond, CA). OA was from Wako Pure Chemicals (Tokyo, Japan). BFA and OA were prepared as stock solutions in methanol and stored at −20C. Goat anti-mouse IgG1 labeled by FITC and goat anti-mouse IgG2b labeled by rhodamine were purchased from Southern Biotechnology Associates (Birmingham, AL). Biotinylated goat anti-mouse IgM and HRP-conjugated streptavidin were purchased from Vector Laboratories (Burlingame, CA). Goat anti-mouse IgM labeled by 15-nm colloidal gold, goat anti-mouse IgG1, and goat anti-rabbit IgG labeled by 10-nm gold particles were purchased from Zymed (San Francisco, CA) and EY Laboratories (San Mateo, CA). Dulbecco's modified Eagle's minimal essential medium (MEM) was a product of Nissui Pharmaceutical (Tokyo, Japan). DL-isoproterenol-HCl was from Nakarai Tesque (Kyoto, Japan). Medium-grade LR White resin was obtained from London Resin (Basingstoke, Hampshire, UK). All other chemicals were analytical or electron microscopic grade.
Experimental Procedure
Male Wistar rats were injected IP with isoproterenol at a dose of 2 mg/100 g body weight to induce salivary secretion. One hour later, both lobes of parotid glands were excised immediately after exsanguination through the abdominal aorta under ether anesthesia and were dissected into small pieces of lobules. These tissue pieces were incubated in MEM containing 25 mM HEPES, 0.3 mg/ml sodium bicarbonate (pH 7.2), and one of the Golgi-disrupting drugs, either BFA or OA at 37C with constant oscillation in a water bath incubator. The working concentration of the drugs added to the medium and the incubation time were as follows: 0.5 μM OA, up to 30 min or 2.5 μg/ml BFA, up to 90 min. To examine the recovery phase from the effects of BFA or OA, the tissue pieces treated with the Golgi-disrupting drugs as described above were rinsed three times with fresh medium containing no drug and chase incubation was conducted. After these treatments, tissue pieces were processed for immunocytochemistry. At least three incubations from three animals were conducted for every treatment.
SDS-PAGE and Immunoblotting
To evaluate the dynamics of the GF-1 antigen molecule under the effect of OA, immunoblotting for GF-1 was conducted. Tissue pieces were incubated in MEM with or without 0.5 μM OA up to 30 min as above. Then the tissues and culture medium were separated. The medium was concentrated by ultrafiltration to prepare for immunoblotting. The tissue slices were washed with ice-cold PBS and homogenized in a glass homogenizer with a Teflon pestle. Soluble and membrane-bound proteins were separated into aqueous and detergent phase by phase separation with Triton X-114 solution (Bordier 1981). Samples were separated by SDS-PAGE on 10% gels and transferred to nitrocellulose membrane. The membranes were blocked in 1% BSA-PBS before incubation with GF-1 solution. Biotinylated goat anti-mouse IgM and HRP-conjugated streptavidin were used to detect GF-1 antigen.
Immunofluorescent Cytochemistry
Parotid tissue pieces were fixed with PLP solution at 4C for 2 hr. After cryoprotection with gum sucrose, cryostat sections about 6 μM thick were cut and nonspecific staining was blocked with 1% BSA-PBS for 1 hr. The sections were incubated with a mixture of anti-β-COP and anti-mannosidase II in 1% BSA-PBS at RT for 1 hr. After washing in PBS, the sections were incubated with a mixture of FITC-conjugated anti-mouse IgG1 and rhodamine-conjugated anti-mouse IgG2b for 1 hr. Negative control staining was conducted by replacing the primary antibodies with 1% BSA-PBS. Cross-reaction of secondary antibodies against IgG subclass was also examined by swapping the combination with primary antibodies. They were then observed using a confocal scanning laser microscope (MRC-1000; Bio-Rad, Hercules, CA). Serial optical sections (collected at 1-μm intervals) in the z-axis were collected and overlaid for the final images shown.
Immunogold Electron Microscopy
Tissue pieces were fixed with 0.05% glutaraldehyde-4% paraformaldehyde in 0.1 M cacodylate buffer (pH 7.2) for 1 hr. After cryoprotection with 2.3 M sucrose in PBS (Tokuyasu 1980), fixed tissue pieces were quickly frozen in liquid nitrogen. Then ultrathin cryosections were cut on a cryomicrotome (Ultracut S, Reichert FCS; Leica, Vienna, Austria) equipped with a diamond knife. The sections were incubated with 1% BSA-PBS for 1 hr, followed by incubation with anti-β-COP in 1% BSA-PBS at RT for 2 hr. After washing with PBS, they were incubated with anti-mouse IgG1 labeled by gold particles in 1% BSA-PBS for 1 hr. Then the sections were washed, postfixed with 1% osmium tetroxide in 0.1 M cacodylate buffer (pH 7.2) for 10 min, and counterstained with uranyl acetate and lead citrate solutions. Then they were dehydrated through a graded series of ethanol and contrasted by section-embedding in an ultrathin layer of LR White resin (Keller et al. 1984).
High-resolution immunoelectron microscopy using post-fixation with ferocyanide-reduced osmium and embedding in LR white resin was conducted as previously described (Tamaki and Yamashina 1994). Multiple staining with GF-1 and anti-amylase on ultrathin sections was conducted by detection with optimal combinations of secondary antibodies and sizes of colloidal gold particles. Indirect immunoreaction for anti-cathepsin D was carried out separately. Control staining was performed by replacing the primary antibodies with 1% BSA-PBS. All sections were counterstained with uranyl acetate and lead citrate and then examined in a JEM-1200EX electron microscope (JEOL; Tokyo, Japan) operated at 80 kV.
Quantitative Evaluation of Immunogold Labeling
To confirm missorting of GF-1 antigen and cathepsin D, the density of labeling expressed as number of gold particles per μm2 of the different cell compartments (secretory granules, Golgi apparatus, lysosomes, vesicles/ER/cytosol) were evaluated on at least five electron micrographs of the drug-treated cells and untreated cells. The outlines of each cell compartment were traced on a digitizing tablet with a stylus pen and were fed into the TRI image analyzer (Ratoc System Engineering; Tokyo, Japan), and then area measurement was performed. Mean labeling density (±SD) for each cell compartment was determined.
Results
Dynamics of the Golgi-resident Enzyme and Coat Protein
Figure 1 illustrates the distributional change in a Golgi-resident enzyme, mannosidase II (Man II), and a COP-I coat protein (β-COP), in rat parotid acinar cells under the effects of Golgi-disrupting agents. In untreated acinar cells, β-COP was mainly associated with the region of the Golgi apparatus in a reticular form on which Man II was co-localized (Figures 1a-1c). Rapid redistribution of Man II into the region of the endoplasmic reticulum (ER) was obvious in the cells 90 min after BFA treatment (Figure 1f). β-COP also lost its association with the Golgi region and dispersed throughout the cytoplasm (Figures 1d and 1e). OA also disrupts the reticular structure of the Golgi apparatus into many fragments but, in contrast to BFA, redistribution of Man II into the ER could not be observed. Man II was closely associated with the fragmented Golgi apparatus. β-COP was markedly accumulated in the region of the disrupted Golgi on which Man II was retained (Figures 1g-1i).
Immunogold electron microscopy on ultrathin cryosections clarified the distributional change in the β-COP-accompanied structural changes in the Golgi apparatus induced by BFA or OA. Figure 2a shows the immunogold reaction of anti-β-COP on an ultrathin cryosection of normal rat parotid acinar cells. Gold labeling is abundant mostly on the Golgi stack. Figure 2b shows the rapid redistribution of β-COP to the entire cytoplasm in BFA-treated cells. It is apparent that gold labeling is decreased on the degenerating Golgi apparatus and is dispersed throughout the cytoplasm. However, β-COP is closely restricted on the degenerating Golgi apparatus without dispersion to the cytoplasm in the cells 30 min after OA treatment (Figure 2c). In the high-power view, the Golgi cluster was found to be composed of β-COP-positive and -negative vesicles. Both vesicles show somewhat regional distributions in the cluster (Figure 2d).
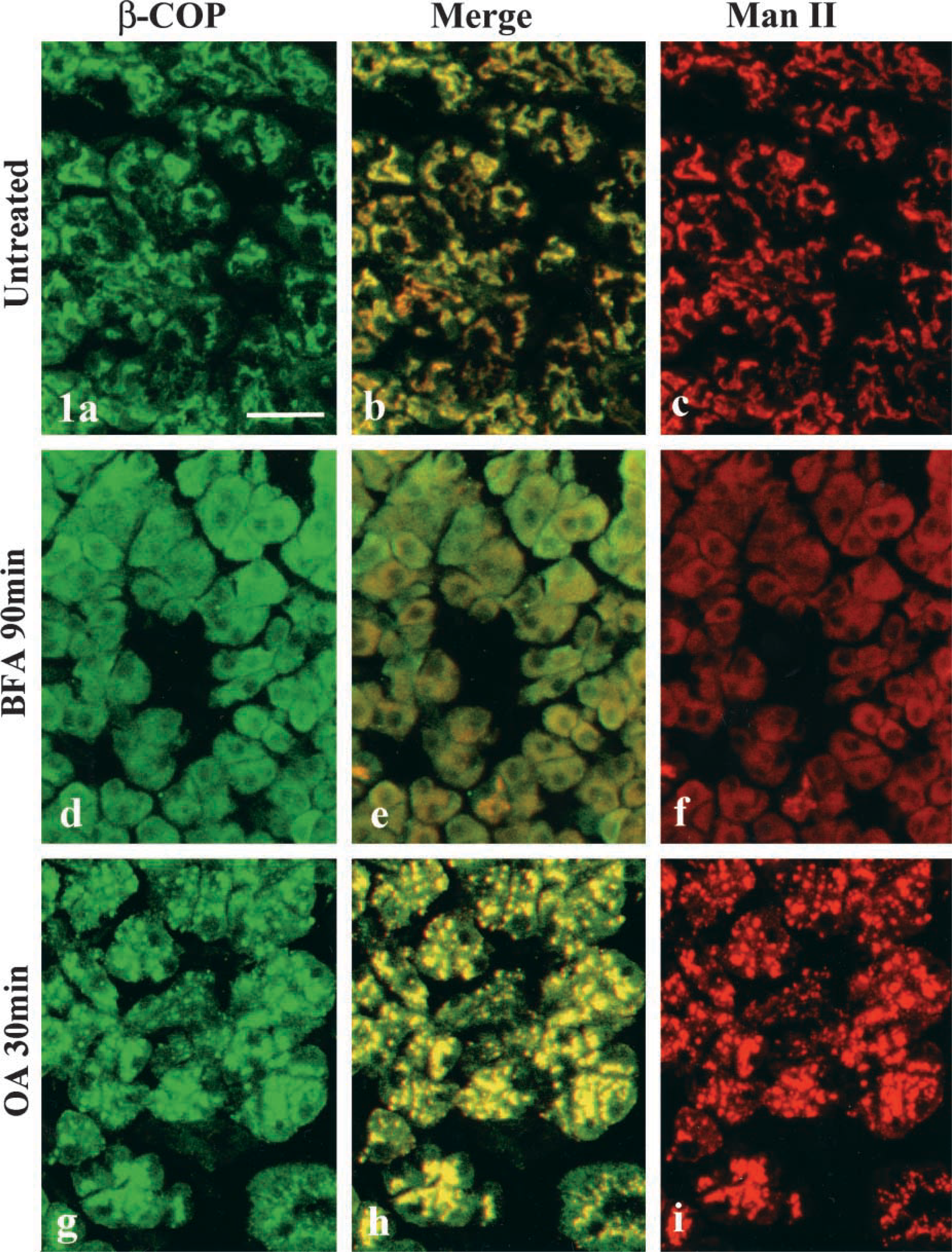
Double immunofluorescent detection for anti-β-COP and anti-mannosidase II. Left column, β-COP (green); right column, mannosidase II (red); center column, merged images. (
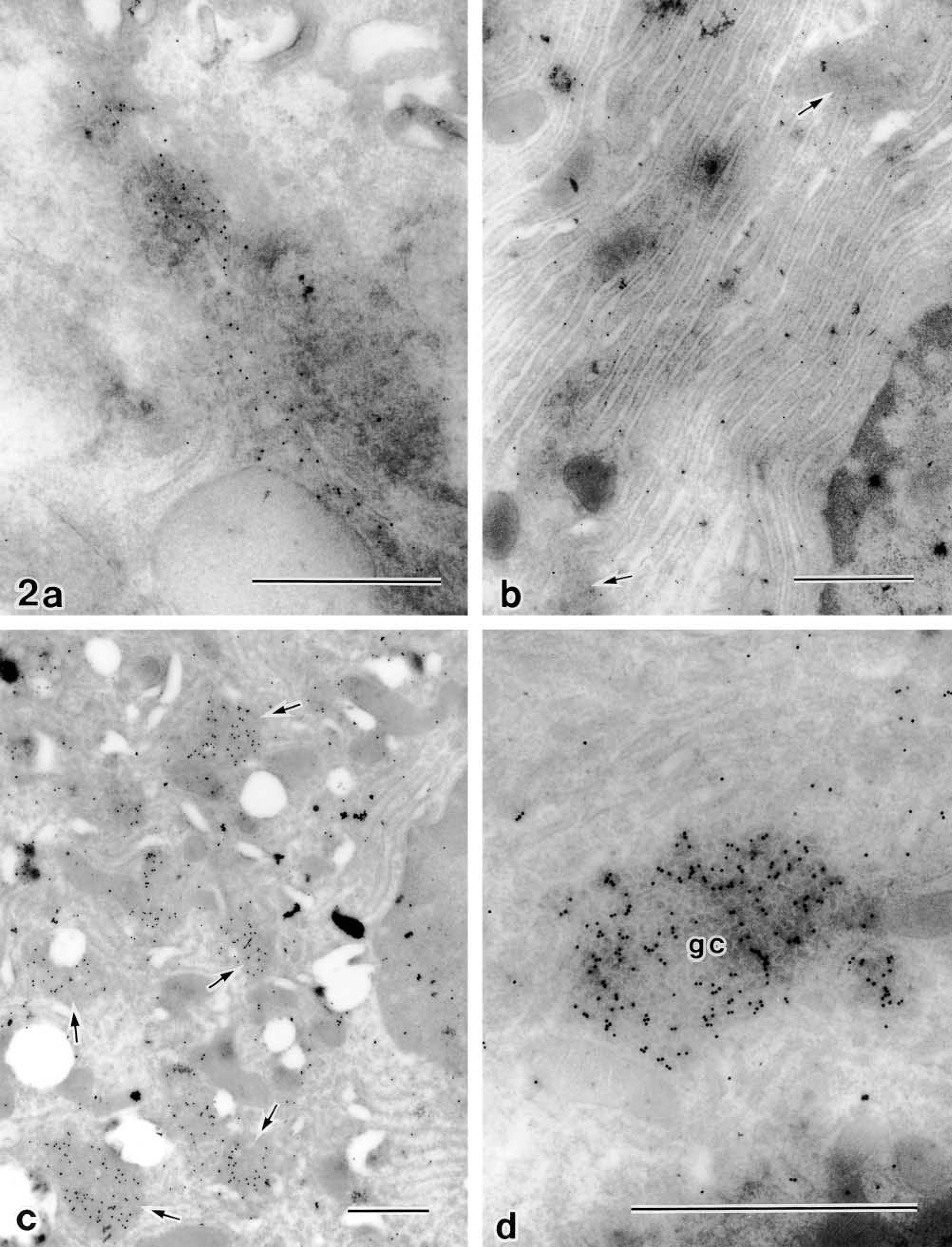
Immunogold staining for β-COP on ultrathin cryosections of parotid acinar cells. (
Distributional Changes of Exportable Proteins and GF-1 Antigenicity
Next, ultrastructural change was collated with the distributional dynamics of exportable proteins and GF-1 antigen, and the effect of BFA and OA was compared by the double-labeling method at the electron microscopic level.
Untreated Cells. Figure 3a shows an electron micrograph of an untreated rat parotid acinar cell doubly stained with anti-amylase and GF-1 immunocytochemistry. The cytoplasmic organelles are well developed for the exocrine function in this cell. The rough endoplasmic reticulum (rER) is tightly stacked in the basal region of the cell. The Golgi apparatus consists of four or five layers of flattened cisternae. Condensing vacuoles of various shapes and sizes are present at the concave face of the Golgi stack. A number of secretory granules are accumulated in the apical cytoplasm. The immunoreactivity of amylase, a major secretory enzyme in this cell, as detected by smaller gold particles (10 nm in diameter) is distributed through the rER, Golgi apparatus, condensing vacuoles, and secretory granules. It is apparent that amylase is gradually condensed in the manner of a regulatory secretion pathway, whereas larger (15 nm) gold particles labeling GF-1 antigenicity are localized on the membranes of the concave trans-region of the Golgi stack, the trans-most discontinuous membranes identified as the trans-Golgi network (TGN), and condensing vacuoles. However, secretory granules, rER, lysosomes, cytosol, and the cell surfaces are completely negative in the reaction. The GF-1 antigen was retained on the membranes of the trans-Golgi area and was never sorted into the post-Golgi transport pathway in normal acinar cells. Immunoreactivity for lysosomal enzyme, cathepsin D, was detectable only on the contents of lysosomes, and the secretory granules are completely negative (Figure 3b). Quantitative analysis of immunoelectron micrographs further confirmed these observations. GF-1 antigenicity was mostly detectable on the Golgi apparatus (Table 1, untreated), and cathepsin D was on lysosomes (Table 2, untreated).
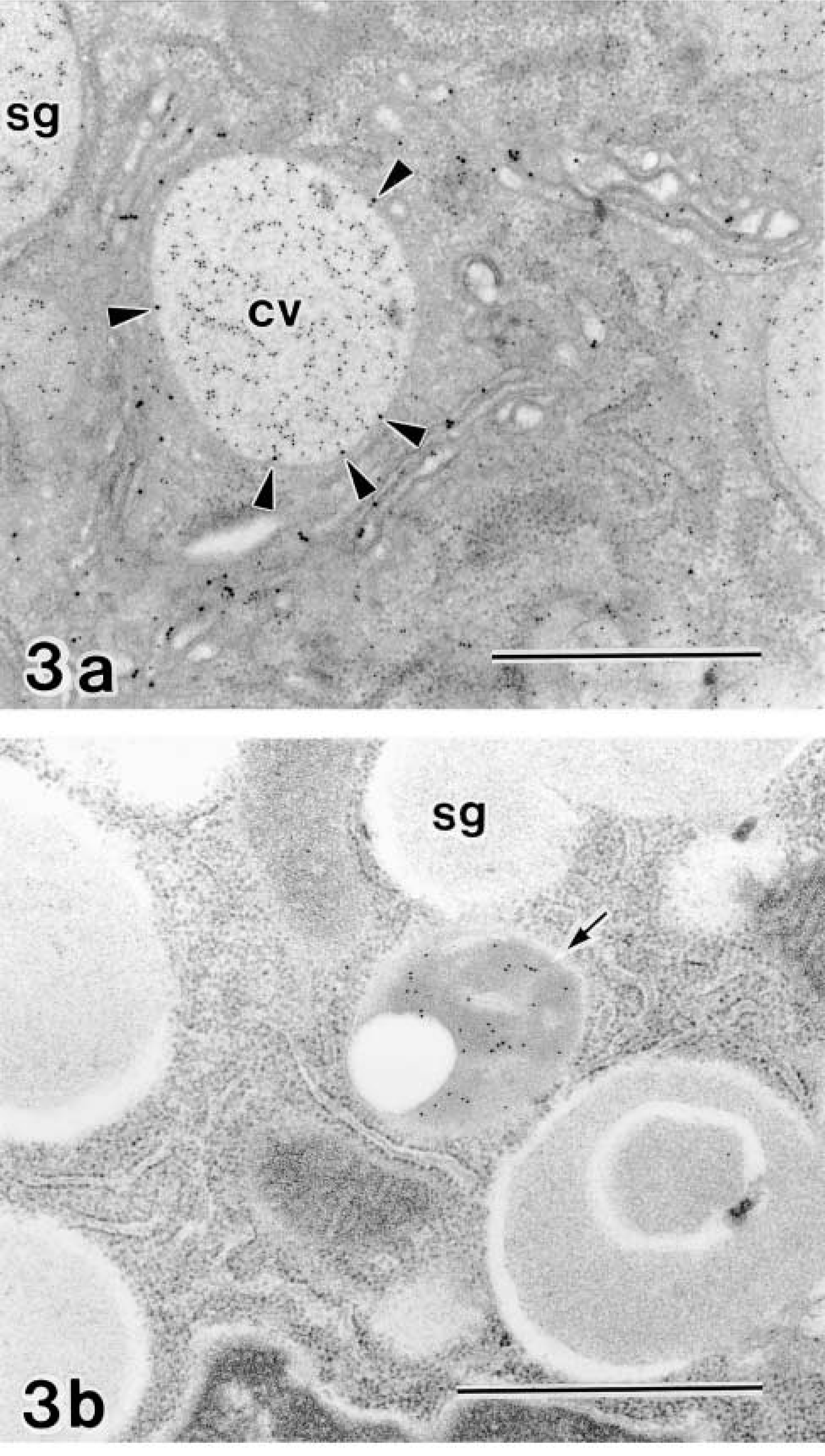
Immunogold staining on untreated rat parotid acinar cells. (
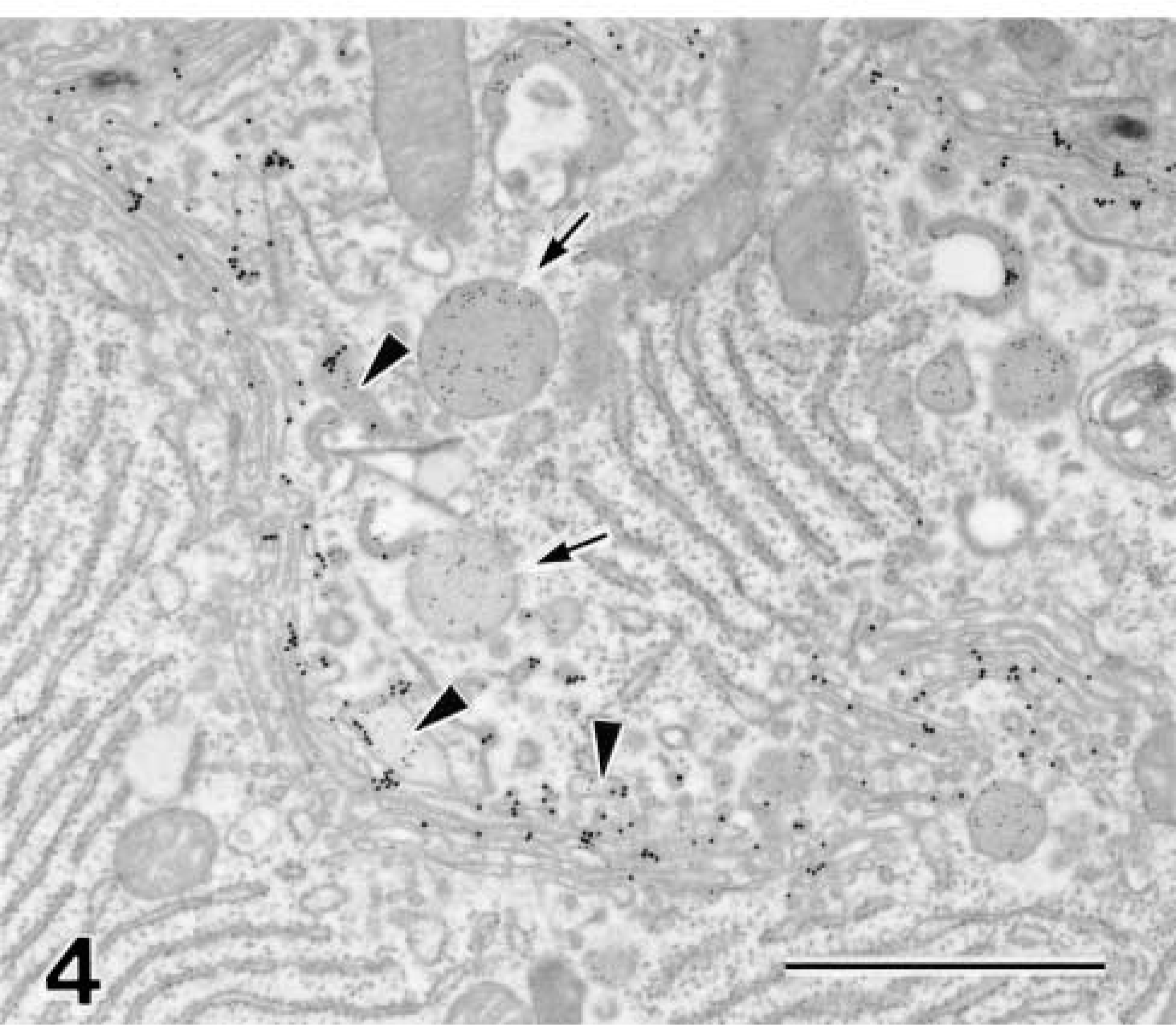
Isoproterenol-stimulated parotid acinar cell double stained by anti-amylase and GF-1. The accumulated secretory granules have been released from the cell by the stimulation. Newly formed secretory granules (arrows) containing amylase (10 nm gold) are observed in the trans-face of the Golgi apparatus. The GF-1 reactivity (15 nm gold) is increased on the trans-region of the Golgi apparatus. The TGN (arrowheads), in which amylase is condensed, is markedly developed. GF-1 antigenicity is strongly detectable on the membranes of trans-cisternae of the Golgi stack and the TGN, whereas secretory granules are completely negative in the reaction. Bar = 1 μm.
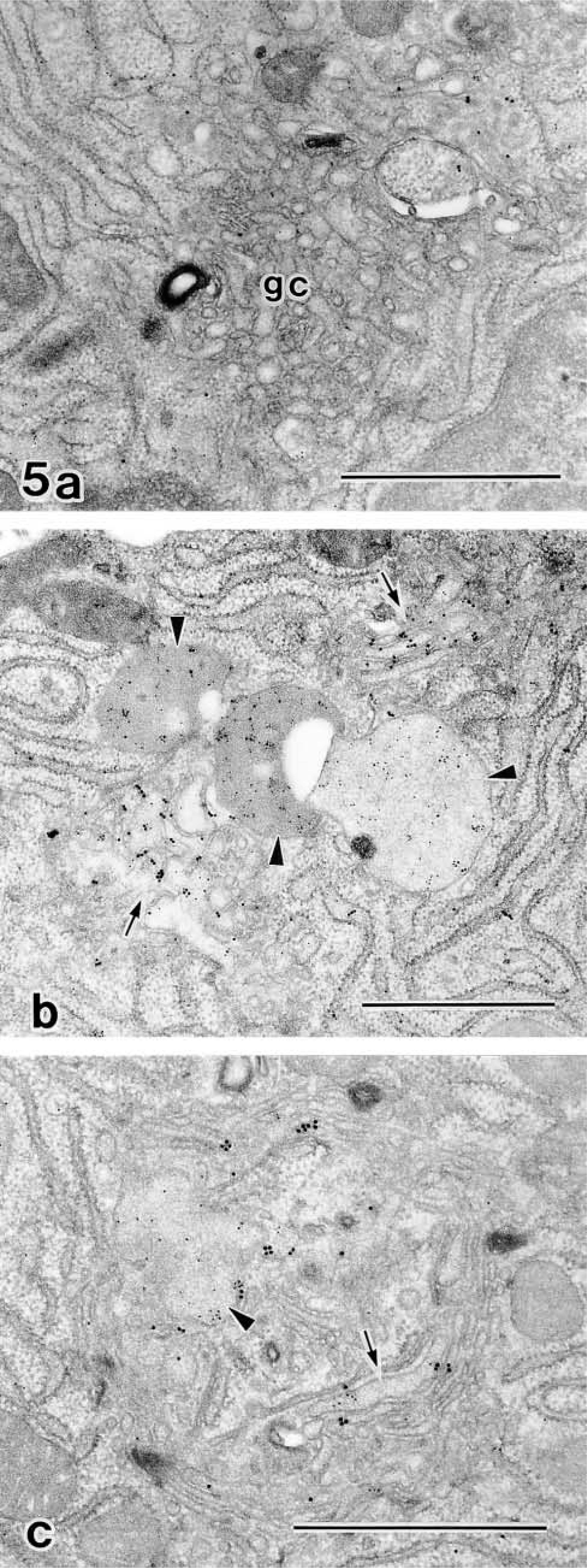
Labeling density for GF-1 a
aAverage number of gold particles/μm2 ± SD.
bParticles on vesicles, endoplasmic reticulum, and cytoplasm.
cCells 60 min after removal of BFA.
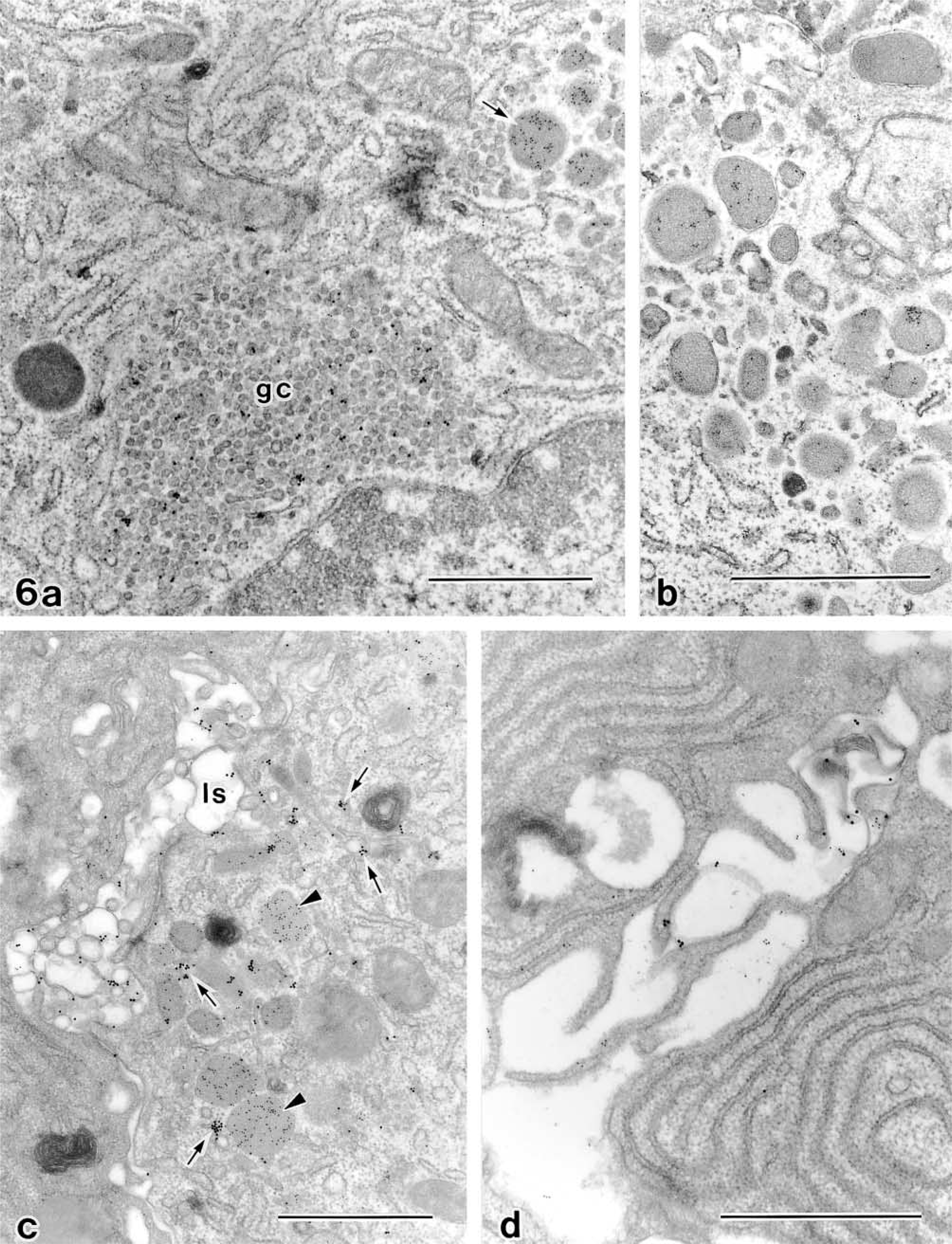
Labeling density for anti-cathepsin D a
aAverage number of gold particles/μm2 ± SD.
Disassembly and reassembly of the Golgi stack under the effects of BFA and its removal in the isoproterenol-stimulated acinar cells. Immunogold reactions for anti-amylase (10 nm gold) and GF-1 (15 nm gold) are doubly detected. (
Isoproterenol-stimulated Cells. Isoproterenol induced complete exocrine release of secretory granules from rat parotid acinar cells, followed by the formation of new granules. Figure 4 shows the Golgi area of the acinar cell after 60 min of secretory stimulation. The apical cytoplasm contained no secretory granules, but newly formed granules were observed in the concaved trans-face of the Golgi apparatus. The area of the trans-Golgi component was markedly extended, and GF-1 reactivity on the trans-Golgi membranes was also apparently increased. These morphological findings strongly suggested that biosynthesis and intracellular transport were enhanced and accelerated by the secretory stimulation. However, GF-1 antigen was still retained on the trans-Golgi membranes and was never transported into newly formed secretory granules, as well as other post-Golgi traffic in such stimulated cells.
Effect of BFA on Stimulated Cells
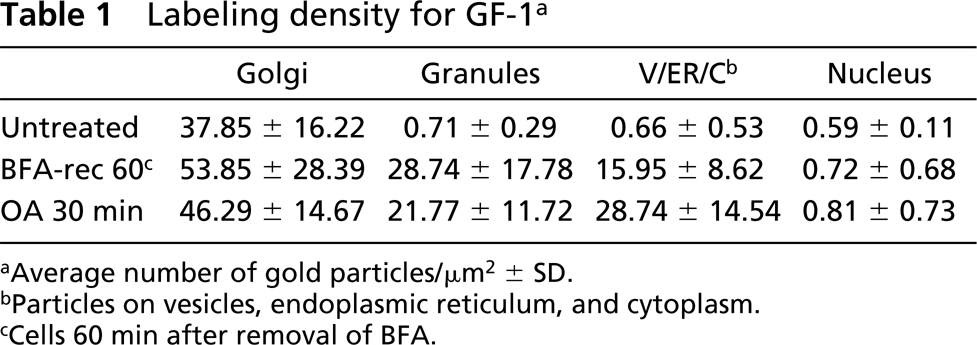
Next we examined effect of BFA on the morphological and functional alterations of the Golgi apparatus during the reconstitution phase of secretory granules in the isoproterenol-treated cells. In parotid acinar cells at 90 min after treatment with BFA, the cisternae of the rER were markedly dilated and filled with electron-lucent material, and the Golgi apparatus completely lost its stacked structure and degenerated into Golgi clusters composed of various shapes and sizes of tubulovesicles. Because of the inhibition of protein transport, neither secretory granules nor condensing vacuoles were formed during the BFA treatment. Both GF-1 and anti-amylase immunoreactivities were decreased on the tubulovesicles of the disrupted Golgi cluster and were markedly redistributed into the dilated cisternae of the rER (Figure 5a).
Such effects of BFA have been known to be fully reversible, and gradual reconstruction of the stacked structure of the Golgi apparatus could also be observed after withdrawal of the drug in the parotid acinar cells (Yamashina et al. 1990). Figure 5b shows a cell at 60 min after removal of BFA. The dilation of the rER has recovered and the re-formation of the Golgi stack was initiated from the tubulovesicles. During this early phase of recovery, the re-forming Golgi saccules showed very short and swollen profiles. In addition, the random distribution of GF-1 antigen indicated that the integration of the cis-trans direction of the Golgi stack has not yet been established in this stage. GF-1 antigen labeled by larger gold particles is distributed in the entire saccules of the developing Golgi apparatus in this stage. Secretory granules containing amylase are newly formed from such an incomplete Golgi stack. A part of the GF-1 antigen is translocated from the Golgi membranes into the contents of newly formed secretory granules, resulting in co-distribution of amylase and GF-1 antigen, indicating that sorting machinery on the Golgi apparatus was out of order when the stacked structure was incomplete en route to recovery from the effect of BFA (Table 1). At 120 min after removal of BFA (Figure 5c), the re-formation of the stacked structure and curved profile of the Golgi apparatus has almost been completed, and the GF-1 antigen became localized to the membranes of the trans-Golgi region and condensing vacuoles. It is apparent that the normal sorting and retention mechanisms on the Golgi apparatus should be reconditioned as soon as the recovery of stacked structure and the internal polarity of Golgi apparatus have been completed.
Effect of OA on Stimulated Cells
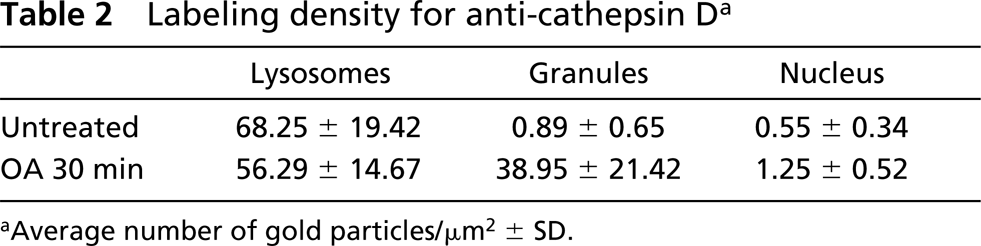
OA also completely disrupted the Golgi stack of acinar cells, within 30 min, into clusters of small vesicles about 70 nm in diameter. However, the manner of the disruption of the Golgi stack differed considerably from that by BFA. The tubular profile of the membrane was very rare and the vesicles in the Golgi clusters were more uniform in shape and size than those obtained by BFA. Neither dilation of the rER nor redistribution of the GF-1 antigen to the rER could be observed during OA treatment. In addition, the formation of the secretory granules was not stalled during the degeneration of the stack (Figure 6a). A small number of the granules, which were smaller than those in normal cells or the cells in the recovery phase from BFA, were observed in the cytoplasm. The granules contained not only amylase but also GF-1 antigenicity. The antigenicity of a lysosomal enzyme, cathepsin D was also abnormally detectable in these granules (Figure 6b; Table 2). In addition, amylase and GF-1 antigen were sorted onto the constitutive pathway. Both antigenicities were detectable on very small vesicle-like structures (Table 2) that apparently differed from secretory granules in morphology and were released into not only the luminal space (Figure 6c) but also into the basolateral space between acinar cells (Figure 6d) without secretory stimulation.
Effects of OA on the parotid acinar cell. Single immunogold reaction for anti-cathepsin D (
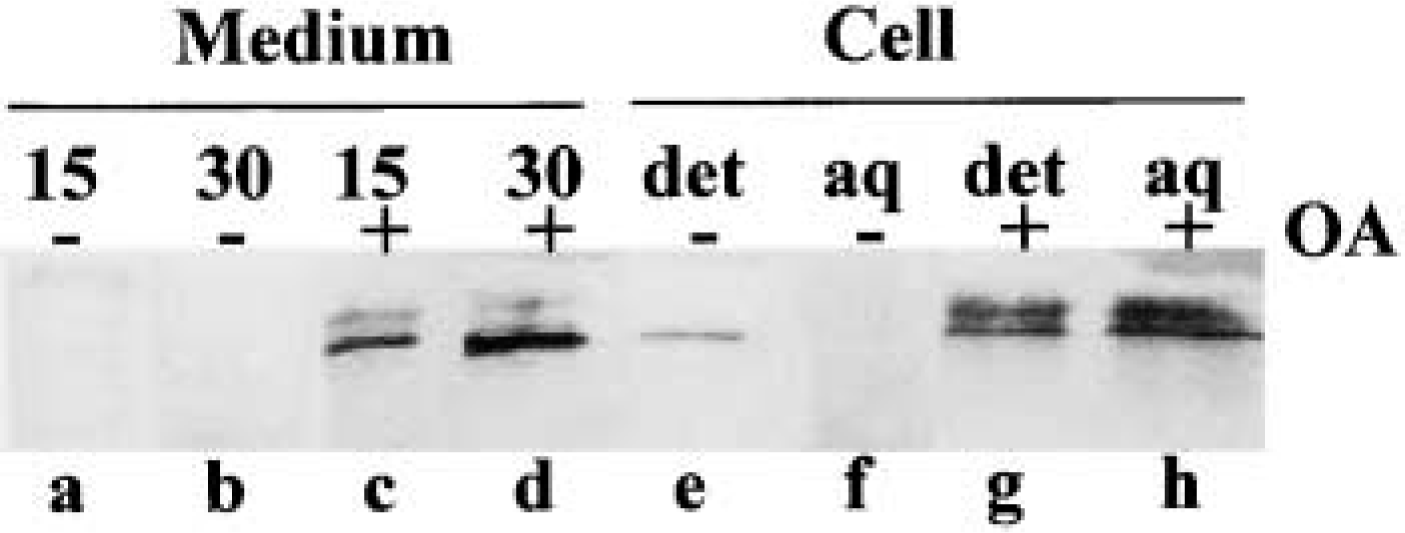
Evaluation of constitutive secretion of GF-1 antigen and sorting into soluble fraction under the effect of OA. Lanes a-f, culture medium; Lanes e-h, cell homogenate. (Lanes a,b) Parotid tissue pieces were incubated at the absence of OA for 15 min and 30 min, respectively. (Lanes c,d), Incubation in the presence of 0.5 μM OA. Note the spontaneous secretion of GF-1 antigen into the medium in OA-treated cells. (Lanes e,f) Fractions from untreated cell. (Lanes g,h) Fractions from cells treated with OA for 30 min. det, detergent phase; aq, aqueous phase. GF-1 antigen appeared not only as the membranous but also as the soluble form in OA-treated cells. Additional GF-1-positive bands of larger molecular weight were detected in OA treatment (Lanes c,d,g,h).
Such missorting and constitutive secretion were confirmed by biochemical assay using immunoblotting for GF-1 antigen. GF-1 antigen is a Golgi membrane-bounded protein and is separated in the detergent phase in untreated cells. In OA-treated cells, however, the protein is transferred to the soluble aqueous phase and is released into the culture medium without secretory stimulation (Figure 7). As above, marked abnormality of sorting and retention mechanisms also took place in the degenerating Golgi apparatus by OA. The Golgi stack-disrupting effect of OA was reversible, similar to that of BFA. Figure 8a shows a cell at 1 hr after the removal of OA. In the clusters of small vesicles, some tubular profiles have started to appear on which GF-1 antigenicity is condensed. These tubular structures should be considered as a primitive form of the reforming Golgi stack. At 2 hr after removal of OA (Figure 8b), re-formation of the Golgi stack continued, from which condensing vacuoles arose. However, GF-1 antigenicity was still distributed through the re-forming stack rich in vesicular membranes and was partially separated from the limiting membranes, resulting in co-localization with amylase in the vacuole contents. At this stage, it was apparent that recovery of sorting machinery had not been completed. At 4 hr after removal (Figure 8c), the re-formation of the Golgi stack was nearly completed. GF-1 antigenicity was restricted to the trans-region of the Golgi apparatus. The condensing vacuole containing only amylase was formed from a well-defined trans-Golgi network. In OA treatment, the normal sorting or retention mechanism was also recovered on the completion of stack formation and establishment of the cis-trans direction of the Golgi apparatus.
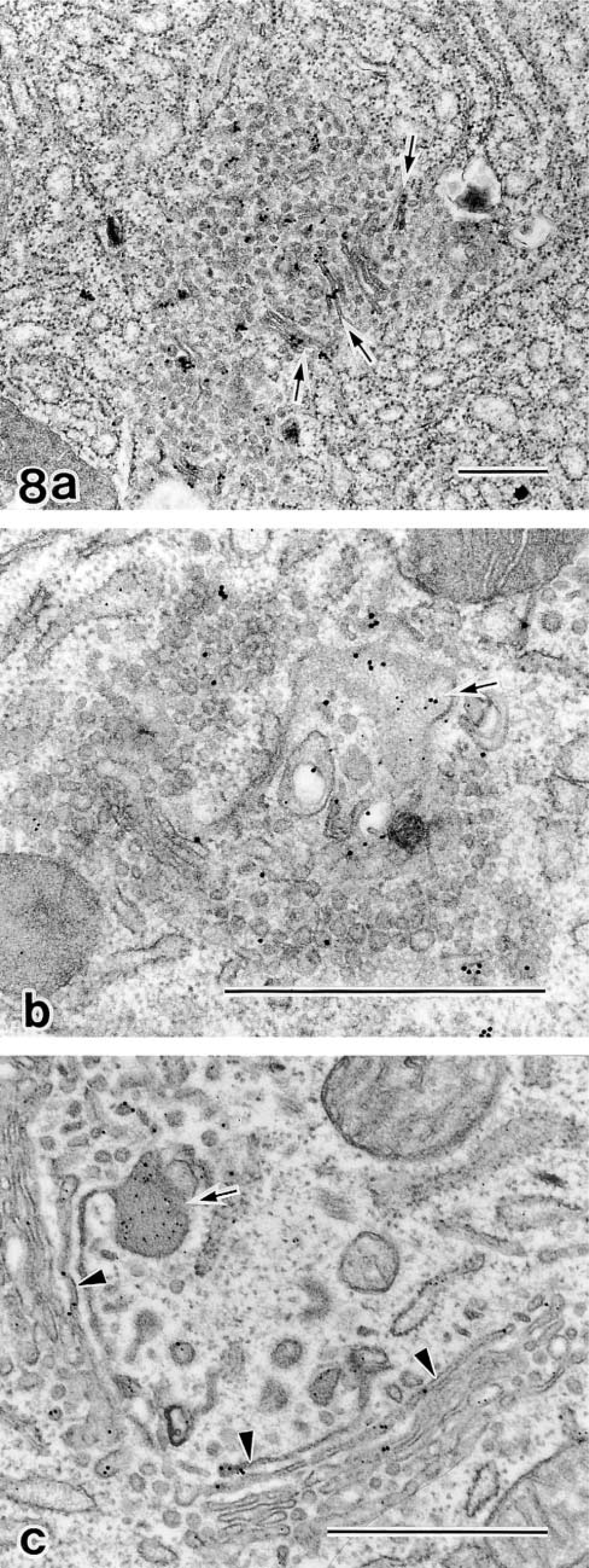
Double staining by GF-1 (15 nm gold) and anti-amylase (10 nm gold) immunocytochemistry in acinar cells en route to recovery from OA. (
Discussion
Disruption of the Golgi Stack by BFA and OA
We demonstrated that the Golgi apparatus of parotid acinar cells shows complete and reversible disorganization of the stacked structure into rudimentary Golgi clusters composed of vesicles or tubules under the effects of both BFA and OA. BFA induced redistribution of Man II and β-COP from the Golgi apparatus into the entire cytoplasm. BFA may inhibit attachment of β-COP to membranes, resulting in the dispersion of β-COP and disturbance of COP-I transport vesicle formation. The interruption of β-COP-associated transport induced by BFA should cause a redistribution of a Golgi membrane protein into the ER, followed by disorganization of the stacked structure.
It is widely known that BFA rapidly and reversibly blocks intracellular transport by preventing the association of β-COP with the donor membranes to cause a concomitant rapid redistribution of Golgi enzymes into the rER and vesiculation of the Golgi stack in many cell types. These phenomena were explained to be a discrete event and, once the transport from rER to the Golgi was inhibited by BFA, the microtubule-dependent tubular structure emanated from the Golgi stack and then the tubule fused to the rER (Lippincott-Schwartz et al. 1989,1990,1995; Sciaky et al. 1997). After fusion, redistribution could occur by diffusive mixing of Golgi and rER membranes or by unidirectional absorption of the Golgi membrane into the rER, resulting in degeneration of the Golgi stack.
OA also induced disorganization of the stack, but the mechanism seemed to be different from that of BFA. OA is a potent inhibitor of protein phosphatase, and has various effects on cellular events that related to intracellular traffic, such as (a) impairment of protein synthesis, (b) inhibition of fluid-phase endocytosis, (c) inhibition of regulated exocytosis, and (d) arrest of transport from the rER to the Golgi apparatus (Takuma and Ichida 1991; Waschulewski et al. 1996). These phenomena were suggested to have great similarities to those normally occurring during the mitotic events (Lucocq et al. 1991). The exact mechanism of how OA induces disruption of the Golgi stack is not fully understood. However, the present study pointed out that the mechanism differed considerably from that of BFA for the following reasons. First, β-COP was not dispersed in the cytoplasm but was retained in the fragmented Golgi apparatus. Second, redistribution of Golgi protein into the rER could not be observed in OA treatment, and most of the GF-1 antigen was retained in the Golgi cluster. Third, the formation of secretory granules was not completely arrested during OA treatment, indicating that post-Golgi transport was not inhibited. Finally, the vesicles composing the Golgi cluster were more uniform in size than those obtained by BFA and showed profiles very similar to transport vesicles. Marked accumulation of β-COP-coated vesicles in the Golgi region induced by OA strongly indicated that imbalance of phosphorylation and dephosphorylation did not prevent β-COP-coated vesicle formation but inhibited their integration to the target Golgi membranes, resulting in inhibition of intra-Golgi transport.
Missorting by a Disorganized Golgi Stack
The most interesting finding in this study was the abnormality of sorting and retention machinery in both the degenerating and regenerating Golgi apparatus. The GF-1 antigen molecule, which is normally attached to the membranes of the trans-Golgi and condensing vacuoles, became detached from membranes and was transported via regulative as well as constitutive pathways. Amylase and cathepsin D were also abnormally sorted into incorrect pathways. Wagner et al. (1994) described that the morphological alterations of the TGN caused by BFA were paralleled by abnormalities in the sorting capacity on the TGN. It is quite likely that the normal sorting and retention machinery on the TGN is broken down as a result of structural disruption of the Golgi apparatus by BFA and OA. The alterations of related molecules could be another explanation for the missorting (Havinga et al. 1985). Unknown post-Golgi sorting or retention signals on molecules could be modified by the disorganized Golgi stack on which processing enzymes are strictly arranged under normal conditions. In immunoblotting of GF-1, another band with larger molecular weight became detectable in OA-treated cells. However, because detailed assay of GF-1 antigen at the molecular level was not conducted in the present study, we cannot fully regard or disregard this possibility. Nevertheless, missorting events could be commonly observed in three different proteins (amylase, GF-1 antigen, and cathepsin D) after administration of two different Golgi disruptive agents, BFA and OA.
Another possible explanation of the expression of the GF-1 antigen into the luminal and the basolateral space is the inhibition of the recycling pathway between the Golgi apparatus and the cell surface by increased phosphorylation induced by OA. Horn and Banting (1994) described that OA induced fragmentation of TGN, in parallel with increased expression of a TGN38 at the cell surface, has been known to recycle between the TGN and the cell surface with a steady-state distribution where the majority is in the TGN. Because the GF-1 antigen is a Golgi-resident protein not localized on the cell surface in normal cells and is not considered to be a recycling protein (Tamaki et al. 1992), this possibility could also be eliminated.
Granule Formation
The distributional changes in the GF-1 antigenicity on the Golgi clusters is another interesting finding in the present study. The Golgi cluster induced by BFA showed faint reactivity for GF-1 and could not produce new secretory granules. Granule formation was re-initiated as soon as the GF-1 antigen was condensed on the reconstructing Golgi stack, whereas the Golgi cluster induced by OA, on which strong GF-1 antigenicity could be detected, had the capability of producing small amounts of secretory granules. A high-resolution immunoelectron microscopic study during the treatments with BFA or OA and during the recovery phase revealed that both degenerating and regenerating Golgi stacks retained the capability of forming secretory granules if the GF-1-positive trans-Golgi elements were maintained but that the sorting machinery within an incomplete stack was considerably disturbed. Normal secretory function was concluded to be fully active only when recovery of the structural integrity of the Golgi apparatus was completed.
Footnotes
Acknowledgements
Supported in part by a grant-in-aid for General Scientific Research (no. 11470008) from the Ministry of Education, Science and Culture of Japan and by the All Kitasato Project Study Grant from the Kitasato Gakuen Foundation.
We thank Mr Osamu Katsumata for technical assistance. The electron microscopy was performed at the Electron Microscopic Laboratory Center of the Kitasato University School of Medicine.