
Abstract
Angiogenesis is an essential process in the progression of malignant tumors. However, little is known of the angioarchitecture in primary oral malignant melanoma. We sought to determine this by the use of periodic acid-Schiff (PAS) stain, endothelial markers (CD34, CD105) and laminin, and by transmission electron microscopy in two cases. The results demonstrated that endothelium-lined vessels dominated the tumor microvasculature and these stained positively for PAS, laminin, and endothelial markers. Mosaic and tumorlined vessels were infrequently encountered. Most PAS-positive patterned networks and loops ultrastructurally represented intratumor microhemorrhages that probably arose secondary to tumor vessel leakiness. Vascular channels of the vasculogenic mimicry type were rare. They stained for laminin but not for endothelial markers.
Keywords
A
Recently, Maniotis et al. (1999) described a new phenomenon, vasculogenic mimicry, which refers to the ability of highly aggressive uveal and cutaneous melanoma cells to generate vascular channels de novo without endothelial cell participation and independent of angiogenesis. Accordingly, melanoma cells are able to engage in vasculogenic mimicry due to their reversion to an embryonic-like phenotype. These vascular channels consisted of patterned networks of interconnected solid or hollow loops of ECM (Maniotis et al. 1999). Vasculogenic mimicry has been described in cytotrophoblasts (Zhou et al. 1997), Drosophilia tumor models (Potter et al. 2000), and invasive ovarian cancers (Sood et al. 2001). This recent find modifies the prevailing view that angiogenesis and related mechanisms are the only means by which a tumor acquires a blood supply for growth and metastases. However, the functionally significant contribution of vasculogenic mimicry was debated because of its profound implications for metastases, drug delivery, and antiangiogenic therapy (Bissell 1999; Fausto 2000; Folberg et al. 2000; McDonald and Foss 2000; McDonald et al. 2000; Shubik and Warren 2000).
Although cutaneous and uveal melanomas represent the two most common forms of malignant melanomas (Makitie et al. 1999; Maniotis et al. 1999; Folberg et al. 2000), primary malignant melanoma of the oral cavity is very rare. In Europe (Gaze et al. 1990; Andersen et al. 1992; Gorsky and Epstein 1998; Nandapalan et al. 1998), the United States (Chiu and Weinstock 1996; Hicks and Flaitz 2000), and Australia (Larsson et al. 1999), it accounts for approximately 1–8% of all malignant melanomas. Its incidence in Japanese is about 11–24%, higher than in whites (Takagi et al. 1974; Kato et al. 1987; Tanaka et al. 1994; Umeda and Shimada 1994). A recent review found that oral malignant melanomas in Japanese show a wide age distribution (range 4 to 92 years), with a peak incidence between the sixth and ninth decades of life and a slight female preponderance (53%) (Tanaka et al. 1994). The sites of predilection are the palate (26%) and maxillary gingivae (21%). Five clinical types are categorized: pigmented nodular, pigmented macular, pigmented mixed, non-pigmented nodular, and non-pigmented mixed types. However, their prognostic value remained unknown (Tanaka et al. 1994). Surgery performed on 72.5% cases yielded only 17% cases surviving more than 3 years. This highlights the need to better understand the factors that influence the prognosis of primary oral malignant melanoma.
Although there have been many reported studies on the microcirculation patterns and their functional significance in uveal and cutaneous malignant melanoma (Makitie et al. 1999; Maniotis et al. 1999; Folberg et al. 2000; Shubik and Warren 2000; Chowers et al. 2001; Hendrix et al. 2001), a search of the existing English language literature disclosed that the microvasculature in primary oral malignant melanoma has not been previously documented. It is also not known whether a similar vasculogenic mimicry phenomenon occurs in this neoplasm. The present study was undertaken to determine the light microscopic, immunohistochemical, and ultrastructural characteristics of the microvasculature in two primary oral malignant melanomas to gain some insights into the angioarchitecture of this neoplasm. This report presents our findings.
Materials and Methods
Tissue Specimens
From the series of eight cases of primary oral malignant melanoma diagnosed at the Department of Oral Pathology and Medicine, Okayama University, two cases representing the most recently diagnosed (1998 and 1999) were selected for this preliminary study. This is to minimize the inherent problems of preservation of formalin-fixed, paraffin-embedded tissue blocks that may compromise optimal immunohistochemical staining. These slides were reviewed and confirmed as primary oral malignant melanomas. New 4–5 μm sections were cut and stained routinely with hematoxylin-eosin and periodic acid-Schiff (PAS) without hematoxylin counterstaining so as to reduce background cellular details that may mask the detection of PAS-positive patterned networks.
Immunohistochemistry
For immunohistochemistry, three primary antibodies were selected, two known to react with the cell membrane of endothelial cells and one with the basal lamina. CD34, a panendothelial marker, is a transmembrane glycoprotein of 115 kD. It is present on immature hematopoietic precursor cells and endothelial cells, but its function awaits clarification (Fina et al. 1990). Endoglin (CD105) is an Mr 180,000 homodimeric membrane glycoprotein expressed on endothelial cells, which acts as a receptor for transforming growth factor-β1 and TGF-B3 (Fonsatti et al. 2001; Tanaka et al. 2001). It has been shown to be important for developmental angiogenesis and plays an important role in capillary-like tube formation by endothelial cells in three-dimensional collagen matrix cultures. CD105 exhibits a greater affinity for activated endothelial cells in angiogenic tissues, including tumors, but weakly or not at all with those of most normal tissues. Its superiority as a marker in angiogenic studies lies in the fact that it is used as an activated endothelial cell-specific marker and hence detects new blood vessel formation (Fonsatti et al. 2001). Laminin, a major component of the basal lamina of blood vessels, is a critical glycoprotein in tube formation by endothelial cells in three-dimensional collagen gels (Ponce et al. 1999). All monoclonal antibodies (MAbs) were from Dako (Carpinteria, CA).
For immunohistochemical staining, deparaffinized sections were pretreated for antigen retrieval by autoclave heating (132C, 3 min) in 10 nM citrate buffer (pH 3.3, 5 min). These sections were then immersed in 0.3% methanol containing 1% H2O2 for 30 min to block endogenous peroxidase and rinsed in 0.05 M TBS (5 min, three times) before immersion in blocking solution (Funakoshi; Tokyo, Japan) for 10 min at room temperature. Then the sections were covered with the primary antibody and incubated overnight at 4C. The immunoreaction was performed using an EnVision/AP kit (Dako; Kyoto, Japan). The antigenic sites were demonstrated by reacting sections with New Fuchsin kit according to manufacturer's instructions (Dako). The nuclei were counterstained with hematoxylin. For negative control, sections were reacted with normal rat serum or with the secondary antibody alone. All the control sections were negative. Positive staining controls were included for each antibody and, where present in the specimens, internal staining controls were also checked for appropriate reactions with each antibody.
Transmission Electron Microscopy
Tissue samples containing small regions of tumor with vascular loops localized from light microscopic sections were microdissected out, postfixed in 2.5% glutaraldehyde, and postfixed further in a solution of 1% OsO4. These were dehydrated and embedded according to standard procedure, and thin sections were cut and stained with uranyl acetatelead citrate and examined with a EF LEO912 transmission electron microscope.
Results
Light Microscopic and Immunohistochemical Observations
Microscopically, both tumors consisted of solid sheets of melanoma cells disposed in a predominant lobular growth pattern (Figure 1A). Melanin pigmentation was patchy and of a moderate amount. Except for one case in which the surface oral mucosa was ulcerated and covered with a fibrinopurulent membrane, necrosis was not observed in the multiple tumor samples examined.
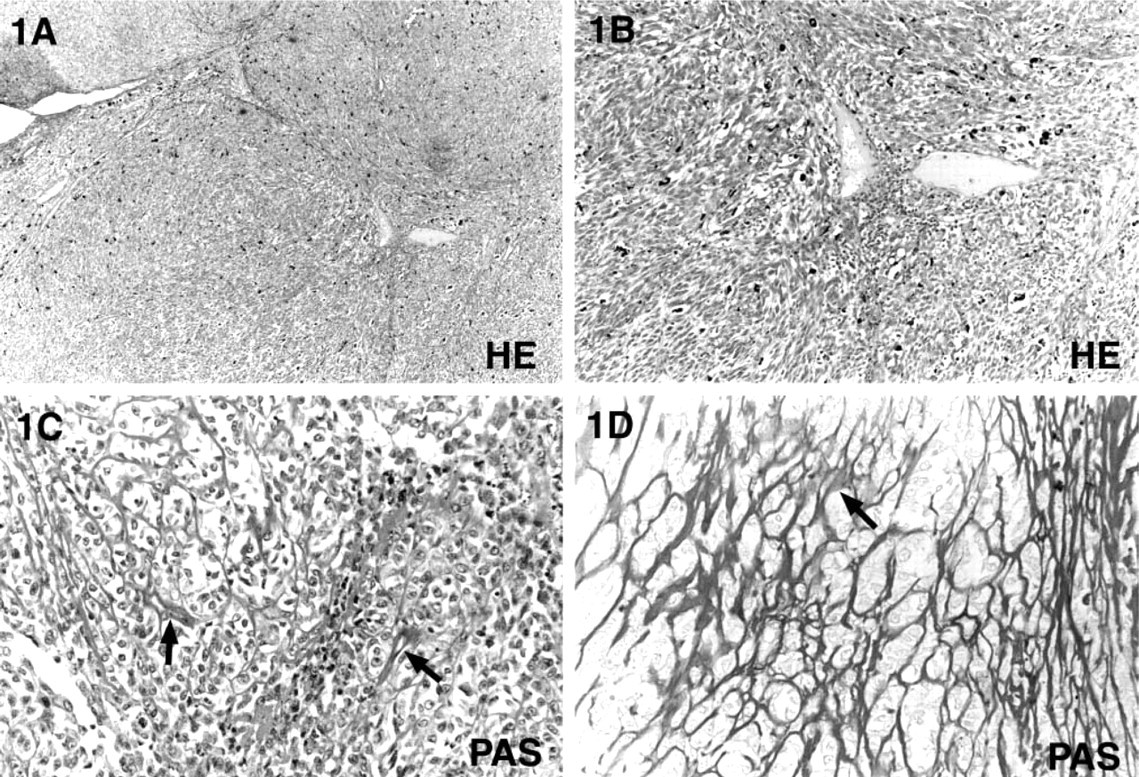
Light microscopic observations of primary oral malignant melanoma showing a predominant lobular growth pattern (
Hematoxylin-eosin-stained sections showed tumor microvasculature consisting of scattered irregularly shaped blood vessels and many slit-like channels also containing blood or fibrinous material (Figures 1A and 1B). PAS staining demonstrated the formation of complex networks and closed loops within the vertical growth phase of these tumors, each loop surrounding smaller lobules or nests of melanoma tumor cells (Figures 1C and 1D). Blood vessels were embedded in the PAS-positive pattern (Figures 1C and 1D).
Immunohistochemical examination showed that some intratumor vessels were positively stained with pan-endothelial marker anti-CD34 antibodies (Figure 2A) but were not positively stained with anti-CD105 antibodies, indicating that these represented normal vessels entrapped within tumor tissues. Other intratumor blood vessels stained positively for both anti-CD34 and anti-CD105 antibodies but with strong expression of the latter marker (Figure 2B), thus indicating that these represented areas of activated endothelial cells and tumor-associated neoangiogenesis (Figure 2B). Moreover, CD105 stained melanoma cells, suggesting that activated melanoma cells may potentially show endothelial cell-like characteristics, which results in vasculogenic mimicry (Figure 2B). However, in areas of PAS-positive vascular loops and patterned networks, a weak to negative reaction for these endothelial markers was observed. Laminin regularly stained the basal lamina of the normal vessels, intratumor blood vessels, and areas containing PAS-positive networks and loops (Figure 2C). In the latter, laminin positivity produced a reticular meshwork pattern (Figure 2C).
Transmission Electron Microscopic Observations
Tumor-associated vessels, consisting of capillary-like structures lined by normal endothelial cells (Figure 3), were identified within the tumor mass. These probably represented entrapped normal vessels and/or new tumor vessels co-opted from the existing host vasculature. In the PAS-positive networks and loops, many of these loops were hollow irregular channels, containing red blood cells and fibrinous material and lined with melanoma tumor cells (Figure 4). Most were intratumor microhemorrhages that probably occurred secondary to tumor vessel leakiness. Residual collagen fibrils were identified within them, confirming that these were stromal compartments and not vascular lumens (data not shown). True tumor-lined vessels and mosaic vessels in which endothelial and melanoma tumor cells formed the luminal surfaces were infrequently encountered. Vascular channels composed of solid or hollow ECM columns were also rare. In these channels, red blood cells and an internal basal lamina lining were identified (Figures 5A and 5B). Melanoma cells lined these channels externally. Endothelium was absent.
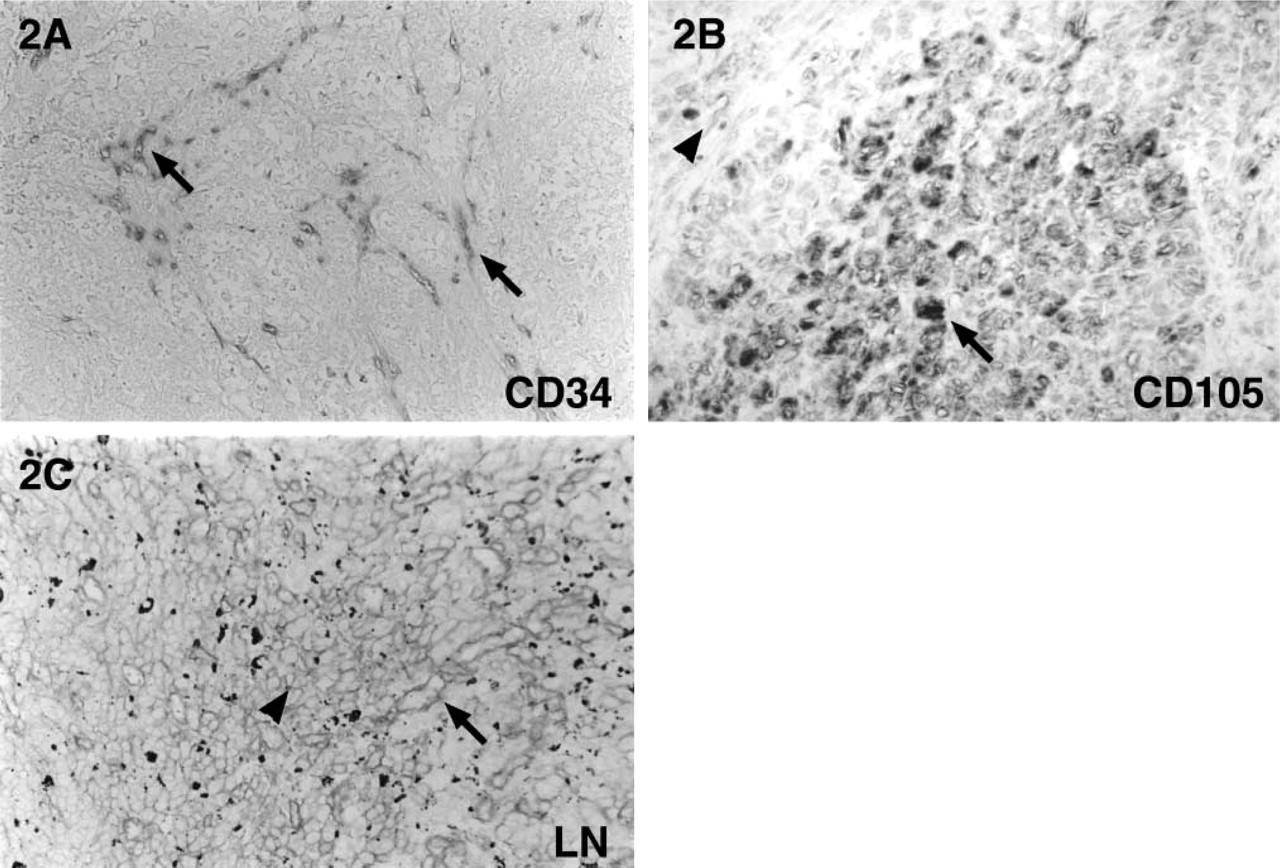
Immunohistochemical staining shows positive expression for CD34 in endothelium-lined normal and tumor vessels (
Discussion
Angiogenesis is an essential process in the progression of malignant tumors during growth and metastasis. This study investigated for the first time the light microscopic, transmission electron microscopic, and immunohistochemical characteristics of the microvasculature in two primary oral malignant melanomas. In these cases, the tumors presented clinically as large pigmented nodular masses arising from the mucoperiosteum of the hard palate and maxillary gingival tissues. Extensive bilateral cervical nodal metastases were present in one case but these tissues were not included for evaluation here.
The present study confirmed earlier reports that the tumor vasculature in malignant melanoma is structurally heterogeneous (Maniotis et al. 1999; Folberg et al. 2000). Within the substance of the tumor mass, normal existing blood vessels were entrapped amongst the endothelium-lined tumor vessels, tumor-lined vessels, and the rare non-endothelium-lined vascular channels. It is well recognized that tumor vessels are highly disorganized structurally and functionally and that they differ from their normal counterparts in many respects. An irregular diameter, infrequent smooth muscle coat, incomplete basement membrane, an abnormal pericyte coat, and an abnormal branching pattern are some of their unusual characteristics (Papadimitrou and Woods 1975; Steinberg et al. 1990; Vermeulen et al. 1995; Hewitt et al. 1997). Consequently, these vessels do not fit well into the usual classification of arterioles, capillaries, or venules.
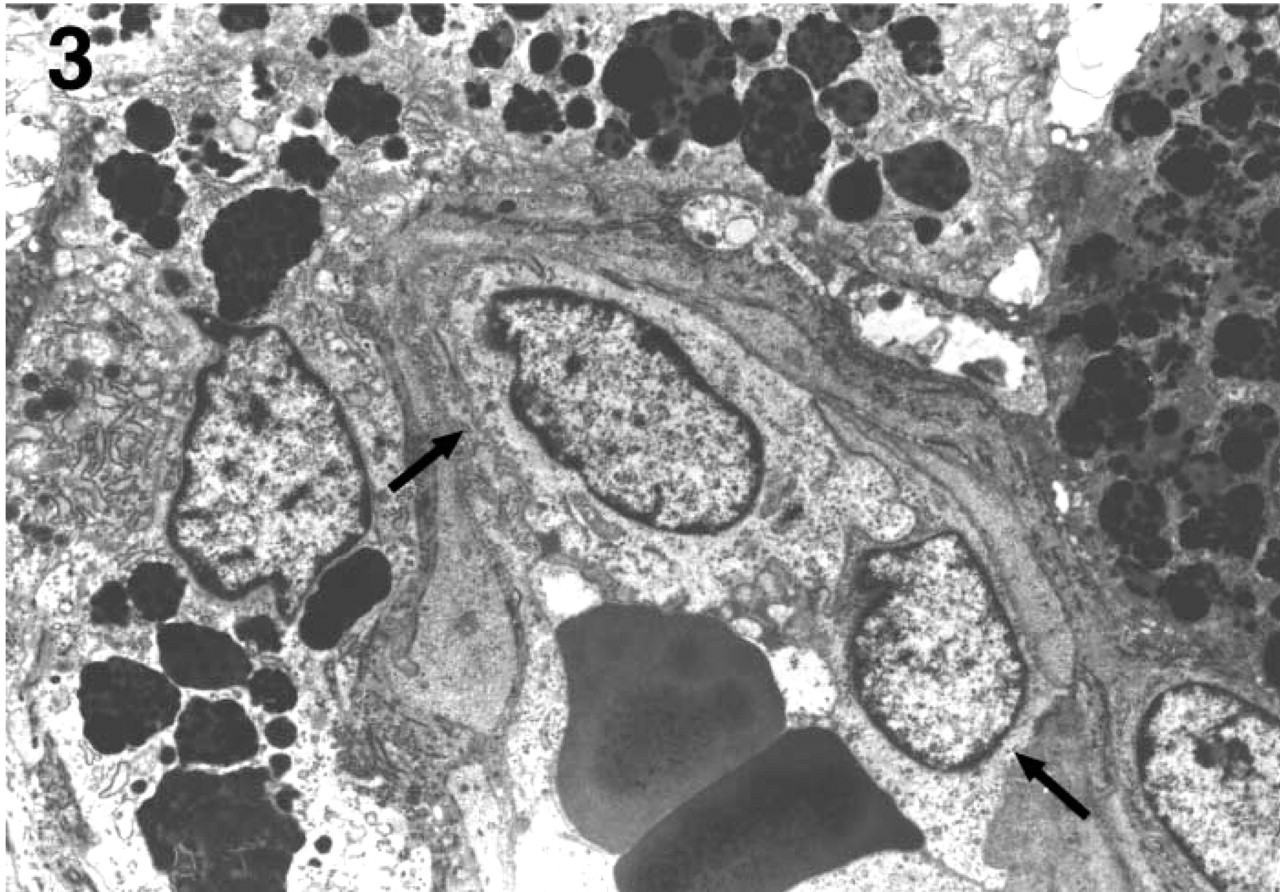
Ultrastructure of tumor-associated vessels consisting of capillary-like structures lined with endothelial cells (arrows). Magnification X6000
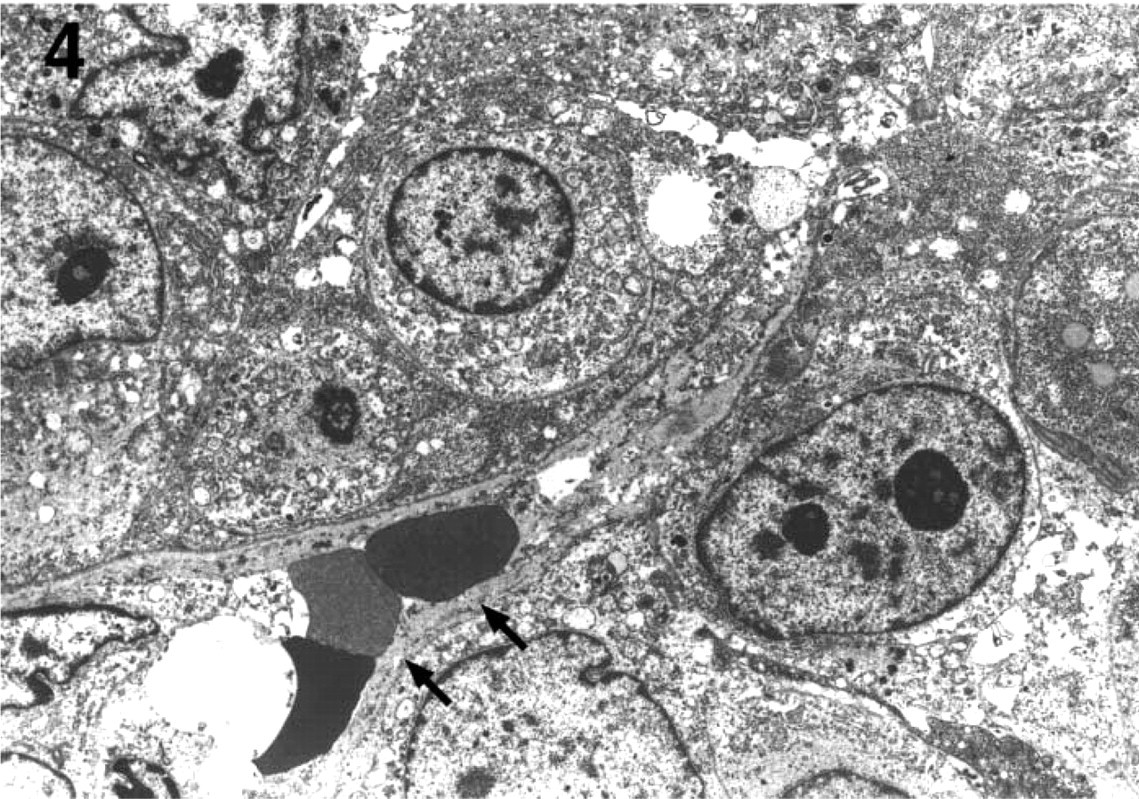
Ultrastructure of vessel-like space with extravasated erythrocytes and fibrinous materials present in the interstices of the melanoma tumor due to intratumor hemorrhage (arrows). Collagen fibrils were identified in these spaces at a higher magnification (not shown), confirming that these spaces are stromal compartments. Magnification X2100.
In this study, we also observed that endothelium-lined tumor vessels in primary oral malignant melanomas differ from those found in the normal palatal mucosa and gingival tissues in these aforementioned aspects. The normal palatal mucosa derives its blood supply from the greater and lesser palatine vessels, and the gingival tissues in the maxilla are supplied by superior alveolar blood vessels. These feeder and exit major vessels were not observed in the melanoma tumor tissue samples examined here. The tumor vessels in this study were distinguished from normal vessels entrapped within tumor tissue by their differential expression of CD105, which detects activated endothelial cells and neoangiogenesis (Fonsatti et al. 2001; Tanaka et al. 2001). However, CD105 does not distinguish vessels formed by tumor-induced neoangiogenesis from those acquired by vessel co-option (Holash et al. 1999). In the latter phenomenon, certain tumor subsets grow initially by co-opting existing host vessels which then regress, leading to a secondarily avascular tumor and massive tumor cell loss. Ultimately, the residual tumor is rescued by robust angiogenesis at the tumor margin. On the basis of this concept, we believe that tumor vessels observed at the periphery of the tumor and in close proximity to normal vessels most likely were vessels co-opted from the existing host vasculature, whereas those tumor vessels located centrally were formed by tumor-induced neoangiogenesis.
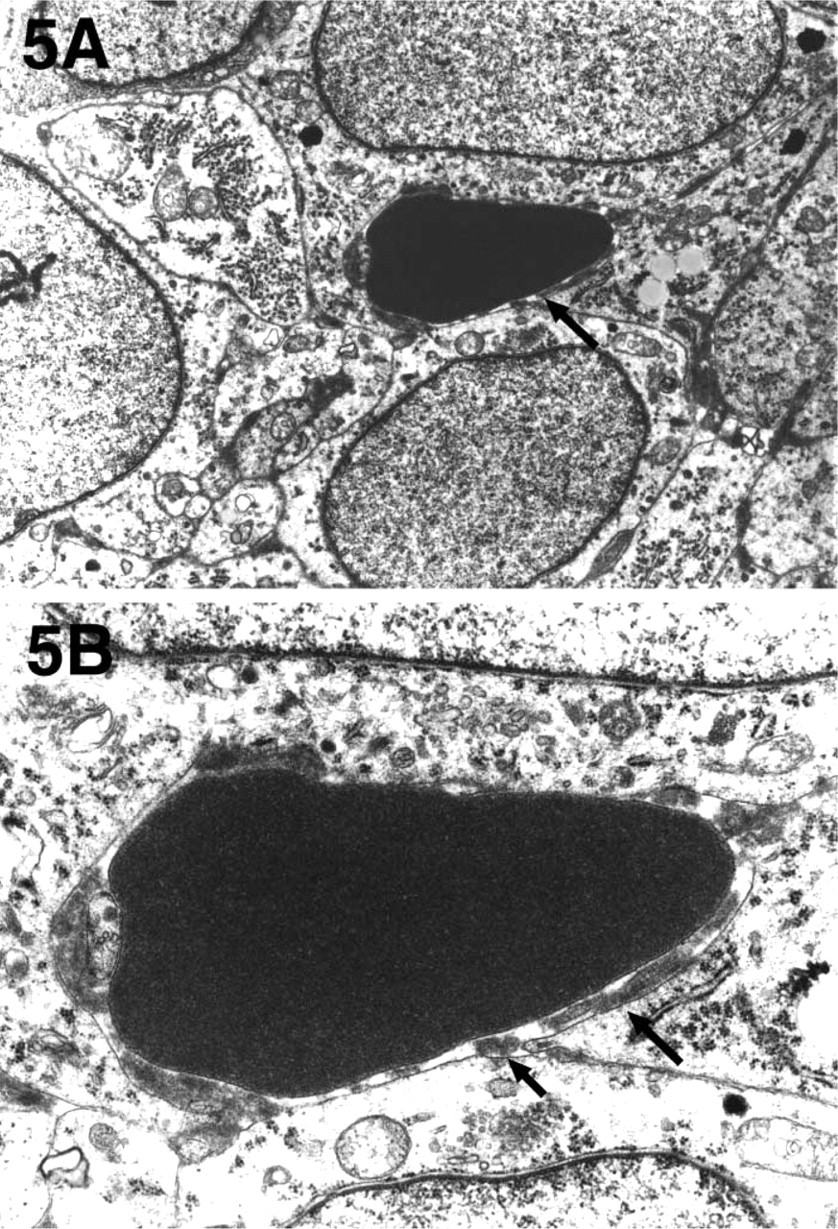
Ultrastructure of a vascular channel, with an erythrocyte identified within its lumen and surrounded externally by melanoma tumor cells (
By the criteria of Maniotis et al. (1999), vascular channels are patterned networks of interconnected loops of PAS-positive ECM, and these may be solid or hollow. In the latter, red blood cells have been detected. Endothelial cells are not identified within these matrix-embedded channels. On the basis of these criteria, we observed that vascular channels were rarely found in the two primary malignant oral melanomas examined here. Although extensive PAS-positive networks and looping patterns were identified at the light microscopic level, many of these loops were confirmed ultrastructurally to represent intratumor microhemorrhages. This finding suggests that extravasation of erythrocytes into the melanoma tumor interstices can produce abnormal vessel-like spaces and, in this instance, also produce a pattern that emulates the interconnecting vascular channel loops and networks formed by vasculogenic mimicry.
Intratumor hemorrhages are believed to be a manifestation of tumor vessel leakiness caused by a defective endothelial monolayer lining these vessels (Hashizume et al. 2000). Studies based on scanning and transmission electron microscopy demonstrated that endothelial cells in tumor vessels are highly disorganized and loosely connected, with formation of intercellular gaps, transendothelial holes, vesiculo-vacuolar organelles, and endothelial fenestrae (Dvorak et al. 1988; Hashizume et al. 2000). Intratumor hemorrhage can range from scattered extravasated erythrocytes to formation of so-called blood lakes. It is possible that the few scattered, thin-walled dilated/sinusoidal vessels seen in the current cases represented these blood lakes. Although the mechanisms of tumor vessel leakiness are unclear, of functional relevance is that vessel leakiness correlates with the malignant potential of tumors because increased endothelial permeability would promote the traffic of tumor cells into the bloodstream and the formation of metastases (Dvorak et al. 1988; Liotta 1992; Ellis and Fidler 1996).
Mosaic blood vessels are characterized by the presence of endothelial and tumor cells that jointly participate to form the internal lining of these vessels (Chang et al. 2000). In their study, Chang et al. (2000) found that approximately 15% of perfused vessels of a colon carcinoma xenograft were mosaic vessels and that they constituted about 4% of total vascular surface. Although these vessels are abundant in human tumors and animal models, their mechanism of formation remains speculative. They differ from the vascular channels of vasculogenic mimicry in that the tumor cells lining the vessel wall do not express an endothelial cell phenotype, i.e., do not stain for endothelial markers (Chang et al. 2000; Folkman 2001). In this study, mosaic blood vessels were infrequently detected. The identification of these vessels by light microscopy, transmission electron microscopy, and immunohistochemistry is not without its shortcomings. We are aware that in practice it is a challenging task to distinguish melanoma tumor cells from endothelial cells and pericytes that line vessel walls both at the light microscopic and the ultrastructural level, especially when cytoplasmic organelles, such as premelanosomes or melanosomes, which serve as distinguishing features of melanoma tumor cells, or Weibel-Palade bodies, which are diagnostic of endothelial cells, were simultaneously absent. Added to this is the problem that the melanoma tumor cell has a tendency to assume a wide range of cytological forms, i.e., epithelioid, spindle, balloon-cell, or clear cell, to name a few. Furthermore, the endothelial cell within the tumor microenvironment has been shown to be cytologically pleomorphic (Hashizume et al. 2000). Immunohistochemical staining with endothelial markers or melanoma cell markers can sometimes yield equivocal results. In addition, it is known that endothelial cells in tumor vessels differ from those of normal vessels not only morphologically but also in their immunophenotypic expression (Chang et al. 2000). To circumvent these identification setbacks, Chang et al. (2000) developed a technique combining utilization of green fluorescent protein, CD31 and CD105, and vascular perfusion to enable simultaneous and differential visualization of cancer cells, endothelial cells, and vessel morphology. Alternatively, immunoelectron microscopy, which combines immunostaining with transmission electron microscopy, would help to resolve some of these identification issues (Sauter et al. 1998).
The significant contribution of the tumor microvasculature in oral malignant melanomas as metastatic pathways remains unclear. This is because oral malignant melanoma, like its cutaneous counterpart, has a propensity to spread via the lymphatic system. Although enlarged lymphatic vessels are frequently observed at the periphery of malignant melanomas, the occurrence of intratumor lymphangiogenesis is questionable (Folkman 1995; Clarijs et al. 2001). One report suggests that the lymphatic microvessel density in cutaneous melanoma is unaltered (de Waal et al. 1997). These issues need to be addressed in future research.
In conclusion, the results of this preliminary study suggest that primary oral malignant melanoma has a heterogeneous tumor microvasculature consisting predominantly of endothelium-lined blood vessels and, to a lesser extent, tumor cell-lined blood vessels and rare matrix-rich vascular channels.
Footnotes
Acknowledgements
Supported by a grant-in-aid for scientific research B from the Ministry of Education, Science, Sports and Culture (no. 12470385 to NN).