
Abstract
Carboxypeptidases may play important role(s) in prohormone processing in normal and neoplastic adenohypophyseal cells of the pituitary. We have recently demonstrated carboxypeptidase E (CPE) and carboxypeptidase Z (CPZ) in the majority of adenohypophyseal cells with carboxypeptidase D (CPD) immunoreactivity largely confined to adrenocorticotrophs. This study evaluated the expression patterns of CPE, CPD, and CPZ immunoreactivity in 48 pituitary adenomas. Our immunohistochemistry demonstrated extensive intracytoplasmic immunoreactivity for CPE, CPD, and CPZ in adrenocorticotrophic hormone (ACTH)-producing adrenocorticotroph cells, prolactin-producing lactotroph cells, and growth hormone (GH)-producing somatotroph cell adenomas, all of which require carboxypeptide processing of prohormones to produce active endocrine hormones. In contrast to the restricted expression in the normal adenohypophysis, CPD appeared to be widespread in the majority of adenomas, suggesting that CPD levels are increased in adenomas. In luteinizing hormone/follicle-stimulating hormone (LH/FSH)-producing gonadotroph adenomas, which do not require carboxypeptidases to produce gonadotropins, only CPZ immunostaining was demonstrated. In null-cell adenomas, CPE immunoreactivity was detected in the majority of tumors, but CPD and CPZ were identified only in a minority of cases. CPE in these cells may process other peptides critical for pituitary cell function, such as chromogranin A or B. These findings suggest that CPs participate in the functioning of pituitary adenomas.
Keywords
S
CPE (enkephalin convertase, carboxypeptidase H; EC 3.4.17.10), is a 50–60 kD protein and is involved in the biosynthesis of most neurotransmitters and peptide hormone. It was first isolated and cloned in humans in 1990 (Manser et al. 1990). CPE is a carboxypeptidase B-like enzyme proposed to be involved in the synthesis of a variety of peptide hormones and neurotransmitters. Lynch et al. (1990) have characterized the immunoreactivity of CPE in the rat CNS, including the pituitary. Immunoreactive CPE was found in melanotrophs of the intermediate pituitary gland, in select cells of the anterior pituitary gland, and in axon terminals or Herring bodies in the neurohypophysis, suggesting that the distribution of CPE co-localizes with that of many neuropeptides, thus further implicating CPE in peptide processing (Lynch et al. 1990). Ultrastructurally, it is localized to the secretory vesicles in adenohypophyseal cells, where it associates with pro-hormones such as pro-opiomelanocortin (POMC), the pro-hormone for ACTH, and pro-chromogranin, and is the only CP detected in mature secretory vesicles (Cool and Loh 1998; Che et al. 2001).
We have recently identified CPE in normal ACTH-synthesizing corticotrophs, GH-producing somatotrophs, and PRL-producing lactotrophs in the human adenohypophysis, suggesting that CPE may participate in prohormone processing to produce these hormones (Fan et al. 2001).
CPD was isolated and cloned in 1995 in bovine, 1997 in human, and 1998 in mouse tissues. It is a 180-kD protein that is more than three times larger than CPE (Song and Fricker 1995). Studies by Fricker and colleagues have shown that CPD is a predominantly membrane-bound trans-Golgi network (TGN) protein that processes protein that transit the secretory pathway within TGN and immature vesicles (Song and Fricker 1996). It is believed that CPD carries out redundant processing functions of CPE. CPD has been detected in a variety of tissues with highest levels in pituitary, brain, and adrenal (Song and Fricker 1995,1996,1997; Tan et al. 1997; Xin et al. 1997; Ishikawa et al. 1998; Varlamov et al. 1999a, b). In normal human adenohypophysis, we have detected CPD immunoreactivity primarily in corticotrophs (Fan et al. 2001), where it may bind to and process POMC to form ACTH (Varlamov et al. 1999a, b).
CPZ, another novel carboxypeptidase, was cloned in human in 1997 and in 1998 in rat (Song and Fricker 1997; Xin et al. 1998). It cleaves substrates with C-terminal arginine residues. Studies by Fricker and co-workers have shown that CPZ plays a select role in the processing of extracellular peptides or proteins (Novikova and Fricker 1999). The mRNA of CPZ was found in placenta, brain, lung, thymus, and kidney by Northern blotting and was localized in leptomeningeal cells by in situ hybridization (Song and Fricker 1997; Xin et al. 1998). We have recently identified relatively widespread CPZ immunoreactivity in adenohypophyseal cells in the human (Fan et al. 2001).
An alteration of prohormone processing has been implicated in the pathogenesis of some pituitary adenomas. Studies by Hashimoto et al. (1994) have demonstrated altered or insufficient prohormone processing in some human pituitary adenomas. Altered processing of the growth-inhibitory prosomatostatin has been observed in GH-producing adenomas (Benlot et al. 1991). Lloyd et al. (1990) have shown that some silent ACTH-secreting adenomas contain abundant POMC mRNA without secreting biologically active ACTH. These findings raise the possibility that some pituitary adenomas may exhibit abnormal carboxypeptidase expression, contributing to the symptomatology associated with ACTH-, GH-, or PRL-secreting adenomas. In this study we evaluated the expression pattern of CPE, CPD, and CPZ in human pituitary adenomas.
The findings presented here suggest significant CPE and CPZ expression in ACTH-, GH-, and PRL-producing pituitary adenomas, but only CPE in null cell-type adenomas. CPD immunoreactivity, which is restricted to ACTH-producing corticotrophs in the normal adenohypophysis, is also extensive in ACTH-, GH-, and PRL-secreting adenomas, suggesting induction of CPD expression in adenomas. This is the first study to demonstrate expression patterns of CPs in human pituitary adenomas.
Materials and Methods
Human Pituitary Adenomas
Forty-eight cases of pituitary adenomas were included in this study. Among them, eight were GH-producing somatotroph cell adenomas, 11 were ACTH-producing corticotroph cell adenomas, 13 were PRL-producing lactotroph cell adenomas, three were luteinizing hormone/follicle stimulating hormone (LH/FSH)-producing gonadotroph cell adenomas, and 13 were null-cell adenomas. Adenomas were classified as “null-cell” adenomas if they exhibited no granular eosinophilic cytoplasm, chromogranin immunostaining, but no or rare LH and or FSH immunoreactivity (Asa 1998). The specimens were obtained at surgery through the transsphenoidal approach. A portion of specimen was used for touch preparation and frozen section diagnoses. The remaining tissue and frozen section controls were further processed for routine formalin-fixed, paraffin-embedded tissue sections. All cases were evaluated with H&E stain and immunohistochemistry for a panel of pituitary hormones, including ACTH, GH, PRL, thyroid-stimulating hormone (TSH), LH, FSH and, in most cases, chromogranin A. Somatotroph cell adenomas were also evaluated with antan, a monoclonal antibody to Cam 5.2. Reticulin and PAS stains were used for evaluation in some cases. Formalin-fixed, paraffin-embedded tissue blocks were retrieved from archival files. For each case, 12 serial sections were cut at 6 μm and placed on positively charged slides. Cases that did not have enough tissue in the blocks for further sectioning (12 serial sections) were excluded from the study. For controls, sections of placenta, normal adrenal gland, and a tumor of A431epidermoid carcinoma were used.
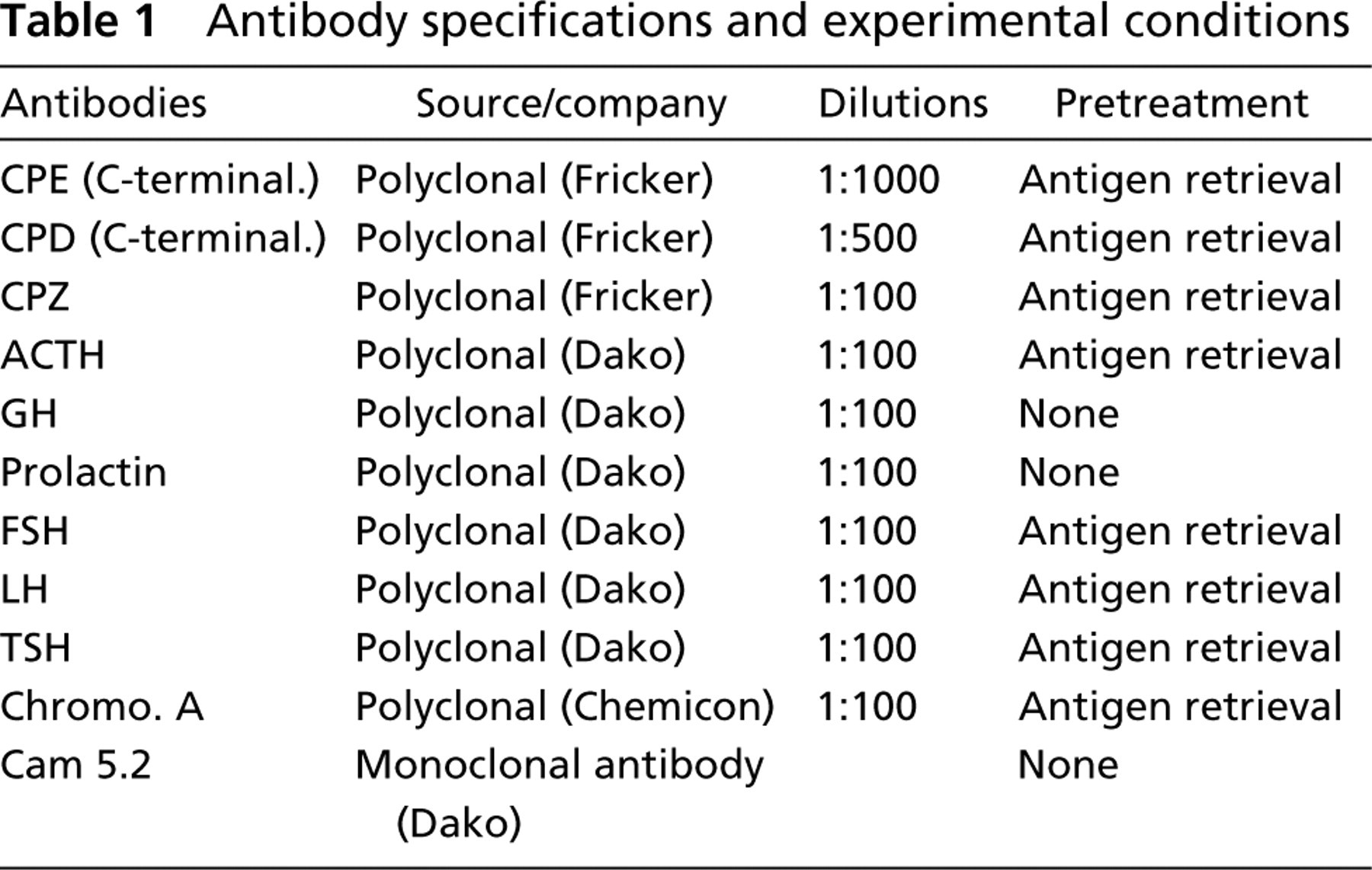
Antibodies
Antibodies against CPE, CPD, and CPZ were gifts from Dr. Lloyd Fricker (Albert Einstein College of Medicine, Bronx, NY). A rabbit polyclonal antiserum raised against the C-terminal portion of CPE was used at a dilution of 1:1000; an antibody raised against the C-terminal portion of the CPD was used at 1:500. A rabbit polyclonal antibody raised against CPZ was used at a dilution of 1:100. The specificity of these antisera has been characterized previously by Western blotting analysis (Song and Fricker 1996; Varlamov et al. 1996; Xin et al. 1997,1998) and immunohistochemistry (Reznik et al. 1998). To assess the possibility of nonspecific binding to keratin, CPD immunoreactivity was also evaluated in a section of epidermoid carcinoma cells and in sections of sommatotroph cell adenomas after preabsorption of the CPD antibody with cytokeratin (Sigma; St Louis, MO). The antibodies raised against ACTH, GH, FSH, LH, TSH, PRL, chromogranin A, and Cam 5.2 were purchased commercially. The technical information and experimental conditions for each antibody are listed in Table 1.
Immunohistochemistry
Immunostaining was performed on a Ventana ES automated immunostainer (Ventana; Tucson, AZ). After trials of different pretreatments for each antibody, the most appropriate conditions for each antibody were chosen, as shown in Table 1. Deparaffinized sections were pretreated with a microwave retrieval system (Antigen Retrieval; Biogenex, San Ramon, CA) and were then incubated with the primary antibodies. The antigen-antibody complexes were visualized by an ABC detection system using diaminobenzidine (DAB) as the chromogen. Normal human pituitary, known to express CPE, CPD, and CPZ, served as positive controls. In addition, sections of normal adrenal gland and placenta were used as a positive control (Song and Fricker 1995,1996), and a section of keratin-rich human epidermoid carcinoma cells was used as a negative control for CPD immunoreactivity. With each of the antibodies, no significant background staining was observed when the primary antibodies were substituted with non-immune rabbit serum or by skipping the primary antibodies.
To further evaluate the specificity of CPD immunoreactivity and possible nonspecific binding to keratin, keratin from human epidermis (Sigma) was resuspended in distilled H2O and added to Ventana diluent containing CPD antibody (1:500 dilution) at a saturating concentration of 200 μg/ml, then incubated at 37C for 32 min (the time and temperature at which antibody was incubated with the tissue sections). Then sections from a “sparsely granular” and typical somatotrophic cell adenoma were subjected to immunohistochemistry as described above.
Scoring for Immunostaining Intensity
The staining intensity and number of stained cells were examined independently by two reviewers (XF and MJ) and were scored as 0 for no significant staining, + for scattered or focal intense immunostaining, and ++ for intense immunostaining in approximately 50% or more of tumor cells.
Antibody specifications and experimental conditions
Summary on CPE and CPD and CPZ immunoreactivity in pituitary adenomas

NA, not available; In, increased.
Results
Expression of CPs in ACTH-producing Corticotrophic Adenomas
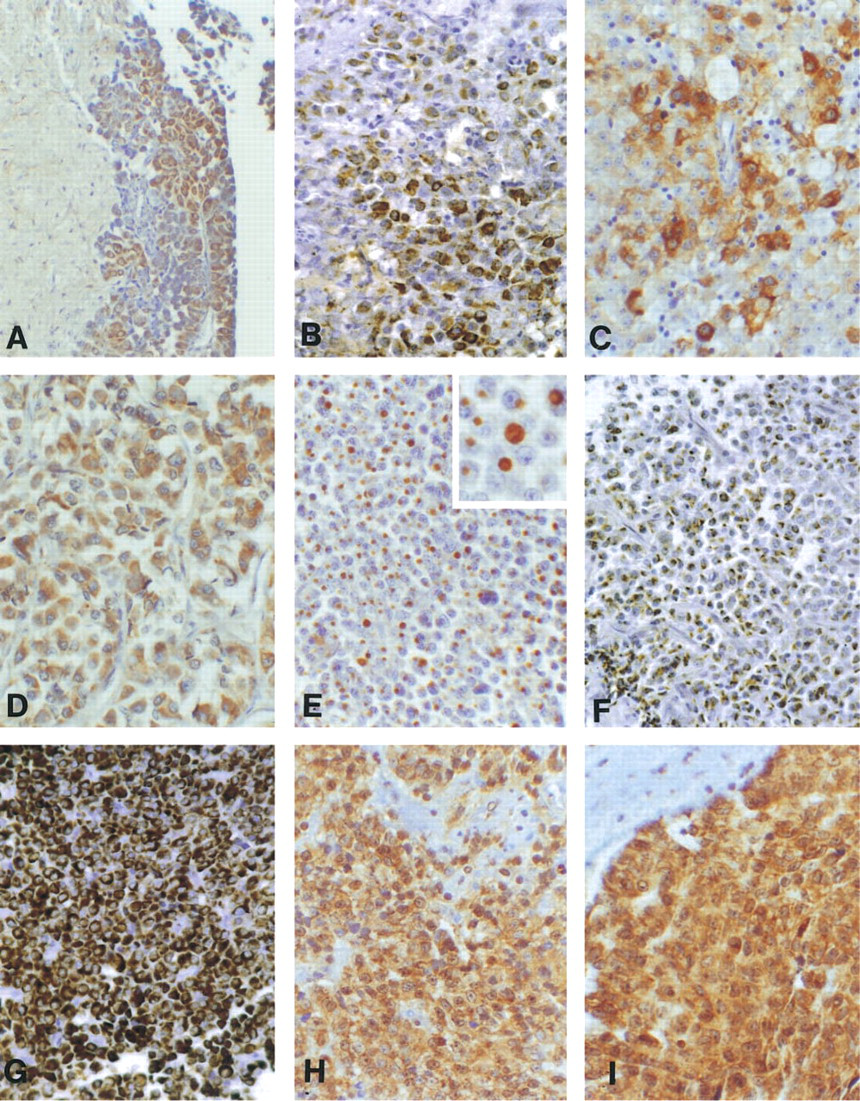
CPE immunoreactivity was detected in 10/11 adenomas. CPD immunoreactivity, which was usually significant, was seen in 10/11. CPZ immunostaining was seen in 8/11 corticotrophic adenomas (Table 2; Figure 1).
Expression of CPs in GH-producing Somatotrophic Cell Adenomas
Cam 5.2 immunoreactivity was dot-like and juxtanuclear, consistent with the pattern of “sparsely granular” adenomas, in two cases. Widespread Cam 5.2 immunoreactivity throughout the cytoplasm was seen in the remaining adenomas studied.
CPE immunoreactivity was detected in 5/8 adenomas. Extensive CPD immunoreactivity was seen in the adrenal gland and epithelium of placenta (positive controls), but no immunostaining was seen in the section of keratin-rich epidermoid carcinoma used as a control for nonspecific binding. In adenomas, CPD immunoreactivity was extensive, seen in 8/8 (Table 2; Figure 1). Moderate to strong juaxtanuclear dot-like immunoreactivity was seen in the two cases exhibiting dot-like Cam 5.2 immunostaining, although the extent of the immunoreactivity varied and often involved a larger zone than that seen with Cam 5.2. In the other adenomas, CPD immunoreactivity was juxtanuclear in a dot-like pattern, sometimes accompanied by perinuclear or cytoplasmic immunostaining. This contrasted with the largely diffuse Cam 5.2 immunostaining in these cases (Figure 1). Preincubation of CPD antibody with keratin had no effect on the distribution or intensity of CPD immunoreactivity in an apparent “sparsely granular” and typical adenoma. CPZ immunostaining was seen in 5/8 somatotrophic adenomas.
Expression of CPs in PLR-producing Lactotrophic Cell Adenomas
Twelve of 13 adenomas exhibited CPE immunoreactivity in tumor cells. The extent of immunoreactivity appeared similar to that in normal adenohypophyseal cells. CPD immunoreactivity, which was usually significant, was seen in 12/13; CPZ immunostaining was seen in 12/13 lactotrophic adenomas (Table 2; Figure 1).
Expression of CPs in LH/FSH-producing Gonadotrophic Cell Adenomas
One of three gonadotrophic cell adenomas exhibited focal CPE immunoreactivity. None of the tumors exhibited CPD immunostaining. In contrast, 2/3 exhibited significant CPZ immunostaining (Table 2).
Expression of CPs in Non-functioning Null-Cell Adenomas
CPE immunoreactivity was detected in 11/13 adenomas. Limited, scattered CPD immunoreactivity was seen in 3/13 adenomas. Six of 13 exhibited limited CPZ immunostaining (Table 2; Figure 1). In contrast, as reported previously, focal CPD and more widespread CPZ immunoreactivity was seen in normal tissue (Table 2).
Discussion
In the present study, extensive CPE, CPD, and CPZ immunoreactivity was detected in ACTH-synthesizing corticotrophic cell adenomas, GH-producing somatotrophic cell adenomas, and PRL-producing lactotrophic cell adenomas, suggesting that they may participate in the processing of a number of precursor peptides in these tumors, including their characteristic hormones and the chromogranins. In a previous study we found extensive CPE and CPZ immunoreactivity in adenohypophyseal cells that secrete ACTH, GH, and PRL. Their expression in adenomas, which in most cases secreted increased amounts of these hormones, suggests that they play a role in possible prohormone processing of these hormones. We have previously found that CPD immunoreactivity in the normal adenohypophysis is largely confined to ACTH-producing corticotrophs (Fan et al. 2001). The findings presented here suggest that its expression is more widespread and is increased in pituitary adenomas that require post-translational processing, and may participate in the hypersecretory activity of these adenomas. In contrast, LH/FSH-secreting gonadotrophic cell adenomas, which synthesize these gonaotrophins as hormones rather than as prohormones, exhibited primarily CPE immunoreactivity but no significant CPD and reduced CPZ immunostaining. Because CPE processes a number of precursor peptides important to pituitary cell function, including prochromogranins A and B, it is likely that CPE expression in these cells participates in their post-translational modification. Taken together, these findings suggest that CPs contribute to the function of adenomas that require post-translational processing.
(
Recent studies in CPE-deficient mice [Cpe(fat)/Cpe(fat)] indicated the existence of CPE-like carboxypeptidases, such as CPD and CPZ. Carboxypeptidase D is a potential candidate to carry out redundant processing functions of CPE based on comparative distribution studies in the rat central nervous system (Dong et al. 1999). The co-localization of carboxypeptidases D and E suggests that carboxypeptidase D may, at least in part, compensate for CPE processing functions in Cpe(fat)/Cpe(fat) mice and may represent a redundant CP. In our study, significant CPD immunoreactivity was found in the majority of functioning adenomas containing CPE. This is in contrast to the distribution of CPD in the normal adenohypophysis, in which CPD immunoreactivity is largely confined to ACTH-producing corticotrophs. This suggests that CPD is more widely expressed in neoplastic adenoma cells, may serve a redundant function there, and may be induced in adenomas that arise from cell types that do not normally express easily detectable CPD.
In the present study, a dot-like staining pattern was seen in the two “sparsely granular” GH-producing somatotrophic cell adenomas. This pattern, along with perinuclear and variable cytoplasmic immunoreactivity was observed in the other somatotroph cell adenomas. The subcellular structures recognized are not known but may represent juxtanuclear/perinuclear and cytoplasmic binding to secretory granules, including those associated with fibrous bodies. The fibrous body (FB), named by Cardell and Knighton in 1966 >, was first described by Racadot in 1964. Despite many subsequent studies, the functional roles of FBs are not established. They are spherical bodies consisting of intermediate filaments (cytokeratins), secretory granules, mitochondria, and lysosomes intermingled with the microfilaments. They localize adjacent to the nucleus in the Golgi region and have frequently been seen in sparsely granulated somatotrophic adenomas, somatolactotrophic adenomas, acidophilic stem-cell adenomas (Asa 1998), and occasionally in variants of densely granular somatotrophic cell adenomas (Kovacs and Horvath 1986). Varlamov et al. (1999a,b) demonstrated CPD in the trans-Golgi network and in nascent secretory vesicles budding from this network in GH3 pituitary adenoma cells. Therefore, the dot-like immunostaining for CPD in somatotrophic adenomas may represent, in part, accumulation of CPD in the secretory granules within the fibrous bodies and may reflect a role in processing of prohormones (Varlamov et al. 1999a, b). Nonspecific binding to keratins is also a possibility. Low molecular weight keratin is typically diffuse or perinuclear in densely granular somatotrophic cell adenomas but is globular and perinuclear in sparsely granulated somatotrophic cell adenomas expressed in fibrous bodies at that site (Ogawa et al. 1990; Asa et al. 1986). Nevertheless, the specificity of the CPD antibody has been extensively characterized (Varlamov and Fricker 1998; Song and Fricker 1995,1996,1997; Reznik et al. 1998). Moreover, the lack of CPD immunostaining in control keratin-rich epidermoid carcinoma cells and the inability of a high concentration of keratin to ablate or reduce CPD immunostaining suggests that the immunostaining seen is at least predominantly to CPD.
LH and FSH are synthesized in their final forms without precursors. Therefore, it is not surprising that LH- and FSH-producing gonadotrophic cell adenomas exhibit little CPE and CPD immunoreactivity. The presence of CPZ in these cells most likely reflects a role in processing other prohormones, such as chromogranin A and B, which are ubiqutiously present in anterior pituitary cells and are synthesized as preprochromogranin. After cleavage by PC1 and PC2, the carboxyl terminals of these prohormones are modified by carboxypeptidases such as CPE, CPD, and CPZ (Che et al. 2001).
Null-cell adenomas are believed to arise from gonadotropin-producing stem cells with very low functional activity (Asa et al. 1986). These adenomas are either devoid of immunoreactivity for anterior pituitary hormones or exhibit only scattered immunoreactivity for FSH, LH, or both. Symptoms in these patients are sequelae of a mass effect rather than due to excess hormone secretion, and therefore it is not surprising that CP immunoreactivity is also limited in null-cell adenomas. CPD was detected in only 2/13 cases; CPZ was demonstrated only in six and was limited in many of these. Studies by Takumi et al. (1998) have demonstrated the presence of PC1 and PC2 in non-functioning adenomas, which they suggested might be related to the processing of chromogranin A. Extensive chromogranin A immunoreactivity was observed in all cases of pituitary adenomas, including null-cell adenomas. There were no significant differences in staining intensity for chromogranin A between normal and neoplastic pituitary cells and cells in different types of adenoma, suggesting that chromogranin A is largely expressed constitutively in normal pituitary and pituitary adenomas. Therefore, the limited expression of CPs in null-cell adenomas may, at least in part, reflect their role in processing of chromogranin A and other peptides critical for adenohypophyseal cell function.
CPD and CPZ are involved in post-translational processing, not only in the normal pituitary gland but also in neoplastic pituitary gland, especially in POMC-, GH-, and PRL-producing adenomas.
Footnotes
Acknowledgements
We wish to thank Ms Rebecca Meitus, Tracy Moss, and Mr Brent Weedman for technical assistance.