
Abstract
Immunohistochemistry (IHC) and recently in situ hybridization (ISH) have elucidated various aspects of human pituitary adenomas, i.e., functional differentiation and classification, transcription factors and mechanism of hormone production, regulation of hormone secretion, and processing of prohormones. Recently, the use of tyramide (catalyzed signal amplification; TSA or CSA) and RT-PCR has been effective for detection of trivial amount of proteins (peptides) and mRNA, respectively. Immunomolecular histochemistry is expected to further clarify the function and biology of human pituitary adenomas.e
Keywords
Human pituitary adenomas account for about 16.3% of intracranial neoplasms and are frequently treated by neurosurgical procedures usually removal by the transsphenoidal approach. The morphology of human pituitary adenomas was first studied by staining methods that was able to clarify different morphological appearances of adenomas according to their clinical function, i.e., hormone production. Classification of human pituitary adenomas has been profoundly assisted by immunohistochemistry (IHC) and, recently, by in situ hybridization (ISH). With the use of these techniques, human pituitary adenomas have been classified into prolactin (PRL)-secreting adenomas, growth hormone (GH)-secreting adenomas, adrenocorticotropic hormone (ACTH)-secreting adenomas, thyroid hormone-stimulating hormone (TSH)-secreting adenomas, gonadotropin (follicle-stimulating hormone, FSH; luteinizing hormone, LH)-secreting adenomas, and nonsecreting adenomas, which are more commonly designated as nonfunctioning adenomas. It is noteworthy that a significant proportion of nonfunctioning adenomas have been found to secrete gonadotropin subunit α or β (Sanno et al. 1992, 1996d). Experimental models of pituitary adenomas have included estrogen-induced PRL-secreting adenomas and TSH-secreting adenomas induced by thyroidectomy. Recently, genetic engineering techniques have produced functioning pituitary adenomas, e.g., GH-secreting adenomas in human growth hormone-releasing hormone (hGHRH) transgenic mice. In addition, the recent development of specific gonadotropin subunit expression in SV40 promoter transformation and subsequent immortalization with established cell lines have contributed to clarification of transcriptional mechanisms (Windle et al. 1990). This review focuses the contribution of IHC and ISH to our understanding of human and experimental pituitary adenomas, i.e. (a) functional classification, (b) transcriptional mechanisms, and (c) secretory processes.
Functions of Human Pituitary Adenomas
Immunohistochemistry has greatly contributed to classification of functioning human pituitary adenomas. The functioning adenomas include GH-secreting adenomas, PRL-secreting adenomas, pro-opiomelanocortin (POMC)-secreting adenomas, TSH-secreting adenomas, and gonadotropin-secreting adenomas. Immunomolecular techniques have shown that GH-secreting adenomas, TSH-secreting adenomas, and gonadotropin-secreting adenomas are frequently multihormonal. Specifically, the GH-secreting adenomas (Figures 1 and 2) are frequently positive for PRL, α-subunit (SU), TSHβ-SU, and FSHβ-SU by IHC and ISH. TSH-secreting adenomas are frequently positive for GH and PRL as well as TSHβ-SU and α-SU. Gonadotropin-secreting adenomas are frequently of the FSH-secreting type and sometimes also produce GH or ACTH. The multiple hormonality of the GH-secreting adenomas may be attributed to co-production of GH, PRL, and α-SU, as well as occasional FSHβ-SU, by the same normal anterior GH cells that are classically categorized as acidophilic. This is a unique situation that could be elucidated only by IHC.
The mechanisms of multihormone production are discussed from the standpoint of transcriptional factors. PRL-secreting adenomas are inevitably monohormonal, and POMC-secreting adenomas are often monohormonal. PRL-secreting adenomas have been successfully treated by bromocriptine (a dopamine agonist), although a few are resistant to this agent. Nonfunctioning adenomas are not infrequently positive for gonadotropin subunits, i.e., α-SU and FSHβ-SU, by IHC and ISH. The histogenesis and functional differentiation of this group of human pituitary adenomas have been debated with regard to their cell lineages (discussed below). Recently, some GH-secreting adenomas have been shown to secrete GHRH from the same cells that secrete GH, and both autocrine and paracrine mechanisms have been proposed for tumorigenesis and functional differentiation (see below). Most GH-secreting adenomas have many osmiophilic secretory granules in their cytoplasm and are designated as densely granulated adenomas. By light microscopic IHC, the tumor cells contain GH diffusely in the cytoplasm. A certain proportion of GH-secreting adenomas contain aggregated keratin filaments, which have been designated as “fibrous bodies” or “keratin bodies.” In these tumors, the numbers of secretory granules are decreased, and they have therefore been designated as sparsely granulated adenomas. In contrast, most PRL-secreting adenomas contain few secretory granules and have been designated as sparsely granulated adenomas. They contain prominent Golgi complexes which have been shown by pre-embedding immunoelectron microscopy to contain PRL. These findings suggest rapid turnover of PRL in the tumor cells.
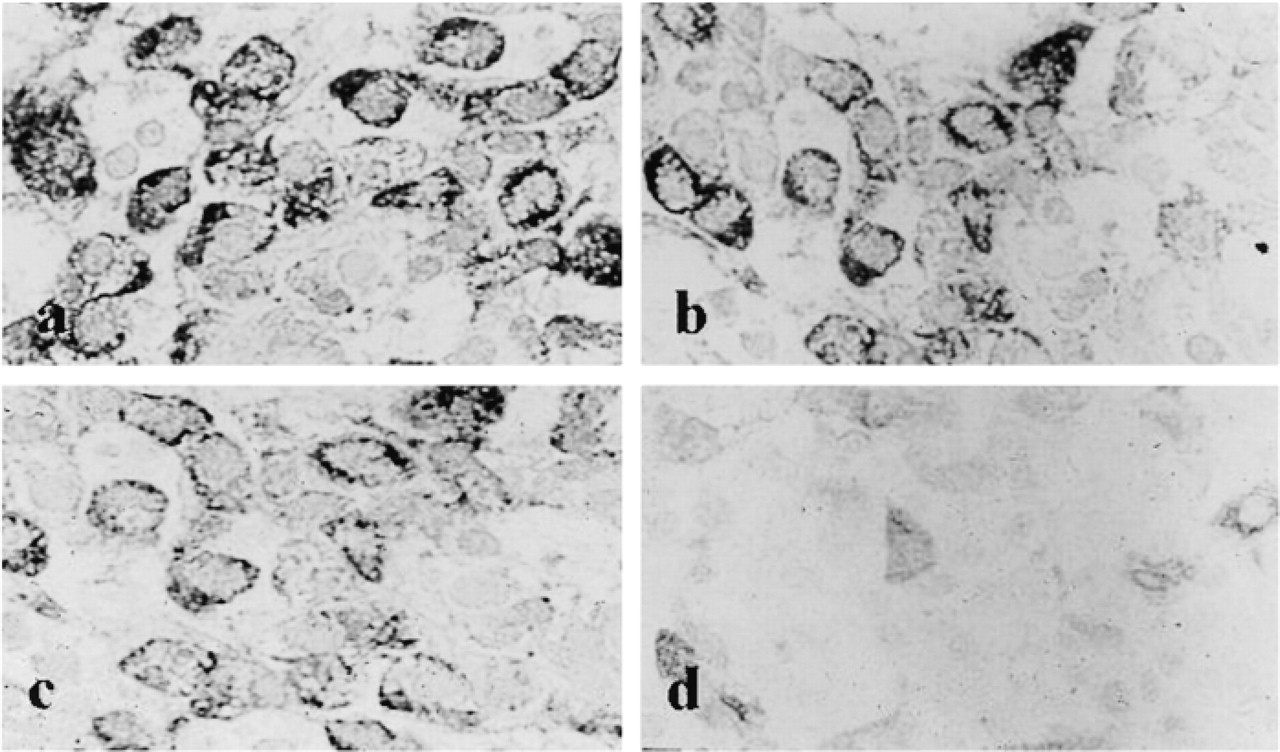
Immunohistochemical staining for GH (
By postembedding immunoelectron microscopy, GH and PRL are localized to the secretory granules in GH-secreting adenomas and PRL-secreting adenomas, respectively. When bromocriptine treatment is effective, the tumor cells of PRL-secreting adenomas exhibit an increase in the numbers of secretory granules, indicating inhibition at the time of exocytosis. The rough endoplasmic reticula and Golgi saccules also become less prominent. For human pituitary adenomas, a variety of morphological and functional classifications have been proposed (Horvath and Kovacs 1980, 1992; Kovacs and Horvath 1986, 1987; Thaper et al. 1993; Osamura et al. 1995). At first they were classified according to their characteristics, i.e., as acidophilic, basophilic, or chromophobic. This was followed by extensive electron microscopic classification, i.e., as densely granulated or sparsely granulated adenomas (Horvath and Kovacs 1980, 1992; Kovacs and Horvath 1986, 1987; Thaper et al. 1993; Osamura et al. 1995). IHC has contributed greatly to clarification of the function of each type of adenoma cell, i.e., GH, PRL, TSH, ACTH, FSH/LH, and ISH now supports the IHC analyses. We have proposed a cytofunctional classification of human pituitary adenomas in which clinical manifestation and morphological characterizations (IHC, ISH, electron microscopy) are taken into account (Tables 1 and 2) (Horvath and Kovacs 1980, 1992; Kovacs and Horvath 1986, 1987; Thaper et al. 1993). In this system, pituitary adenomas are classified as (a) GH-secreting adenomas, (b) PRL-secreting adenomas, (c) TSH-secreting adenomas, (d) ACTH-secreting adenomas, (e) gonadotropin-secreting adenomas, (f) nonfunctioning adenomas, and (g) pituitary carcinoma. IHC is the basic technique used. Each adenoma subgroup is subdivided according to whether the tumor is symptomatic, whether it secretes multiple hormones, and according to its size (macroadenoma, microadenoma, or invasive). For example, when a patient presents with acromegaly, the tumor secretes GH and is multihormonal with concomitant PRL, TSH, and α-SU secretion. The classification of the tumor would therefore be as follows: GH-secreting adenoma; symptomatic (acromegaly); multihormonal (GH, PRL, TSH, α-SU) by IHC and ISH; macroadenoma; and densely granulated. Based on IHC and ISH techniques, this classification provides the clinical and morphological characteristics at a glance. By IHC and ISH analysis, nonfunctioning adenomas not infrequently reveal production of gonadotropin subunits. In these tumors, serum gonadotropin is usually not elevated. At present, it is debatable whether tumors that are positive for gonadotropin subunits should be classified as gonadotropin-secreting adenomas or nonfunctioning adenomas. The use of more sensitive amplification methods, such as tyramide (catalyzed) signal amplification (TSA or CSA), should increase the ability to detect gonadotropin subunits (Sanno et al. 1996d).
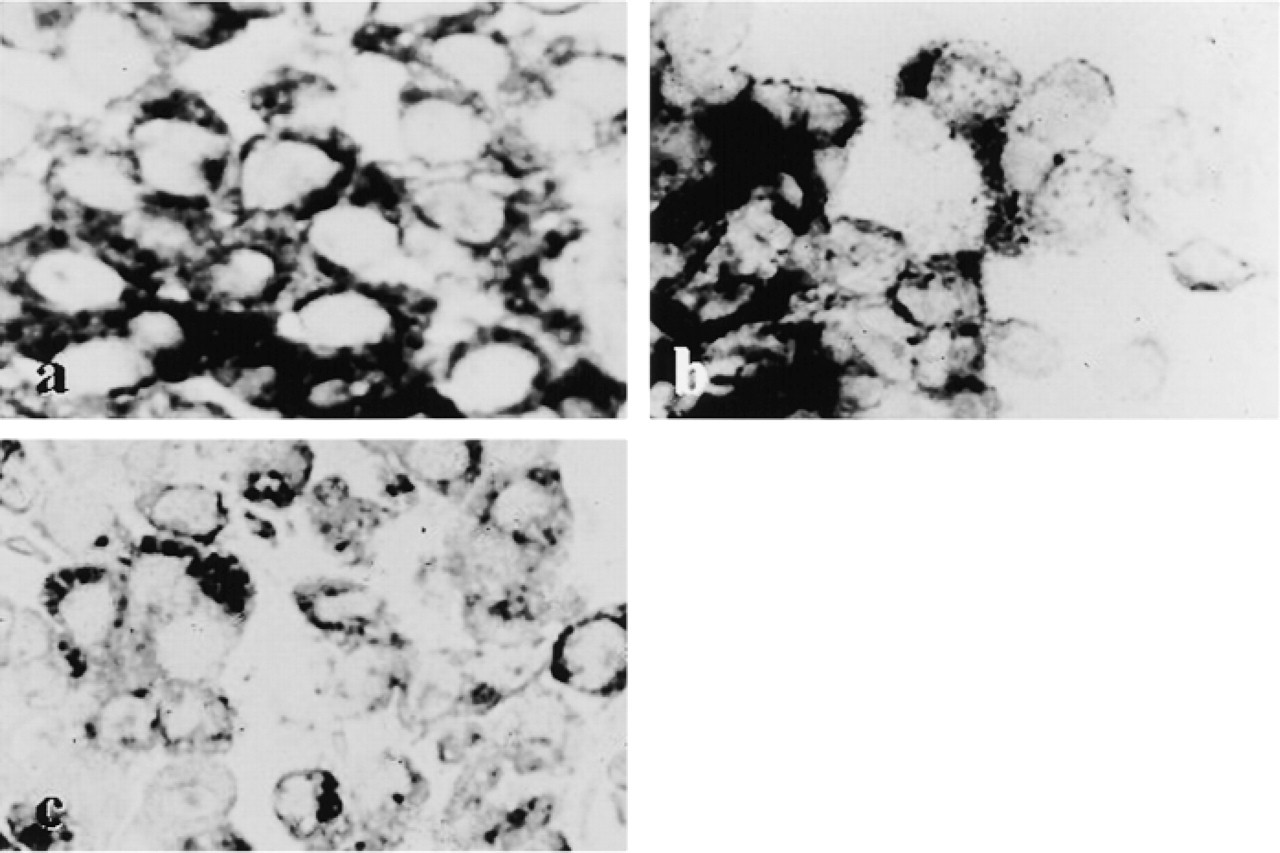
In situ hybridization in GH-secreting adenoma. The adenoma cells are positive for GH mRNA and for PRL mRNA and αSU.
Receptors in Human Pituitary Adenomas
Classification of pituitary tumors

The cells of the anterior pituitary are regulated by hypothalamic hormones and steroid hormones. The receptors corresponding to these hormones reside in the anterior pituitary cells and include the following: growth hormone-releasing hormone receptor (GHRH-R, also known as GRF-R); gonadotropin-releasing hormone receptor (GnRH-R); corticotropin-releasing hormone receptor (CRH-R); dopamine 2 receptor (D2R); estrogen receptor (ER). These receptors have been cloned. GHRH-R, GnRH-R and D2R are seven folded transmembranous proteins. ER, which include ERα and ERβ isoforms, is a nuclear protein that binds to estrogen at a DNA upstream site. ER has been detected by IHC and ISH on paraffin sections. D2R has been detected by ISH. For detection of GnRH-R and GHRH-R, present in the cell membrane in only trivial amounts, in situ RT-PCR has been used as a powerful tool to demonstrate specific mRNA (Sanno et al. 1997c). Sanno et al. (1997a) demonstrated GnRH-R mRNA in a variety of pituitary adenomas (Figure 3). The functional role of ER and D2R should be considered in view of data on bromocriptine treatment. PRL-secreting adenomas are responsive to bromocriptine when D2R is effectively present. The tumor cells of most PRL-secreting adenomas possess ER mRNA. PRL-secreting adenomas are sometimes resistant to bromocriptine treatment despite the presence of D2R mRNA and ER mRNA. The mechanism for this resistance should be further investigated from the standpoint of translation (membrane protein synthesis) or of post-D2R intracellular transduction.
Clinical and cytofunctional classification of pituitary adenomas
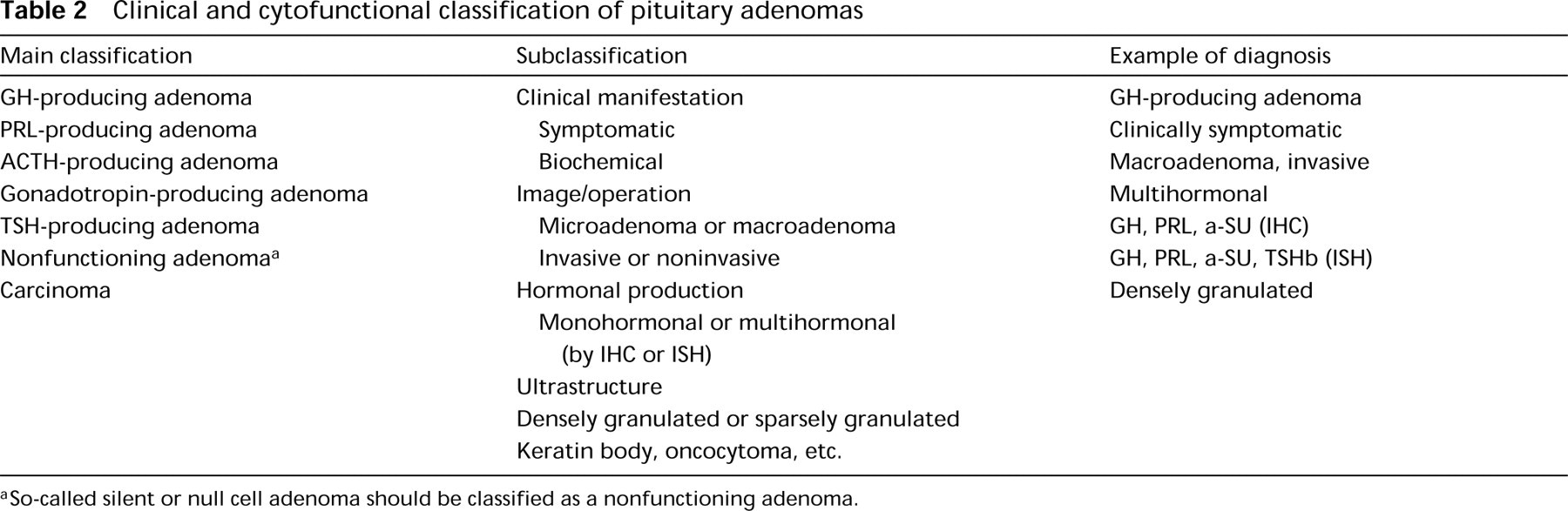
So-called silent or null cell adenoma should be classified as a nonfunctioning adenoma.
Experimental Models of Pituitary Adenomas
Experimental pituitary adenomas have been produced mainly in rodents. Classically, PRL-secreting tumors have induced by prolonged estrogen treatment. These tumors are a useful model for human PRL-secreting adenomas, especially from the aspect of functional and morphological changes induced by treatment with bromocriptine. They are also transplantable and have been established as cell lines (MtT series) (Inoue et al. 1990). Thyroidectomy also leads to induction of TSH-secreting adenomas, which are also transplantable after several passages.
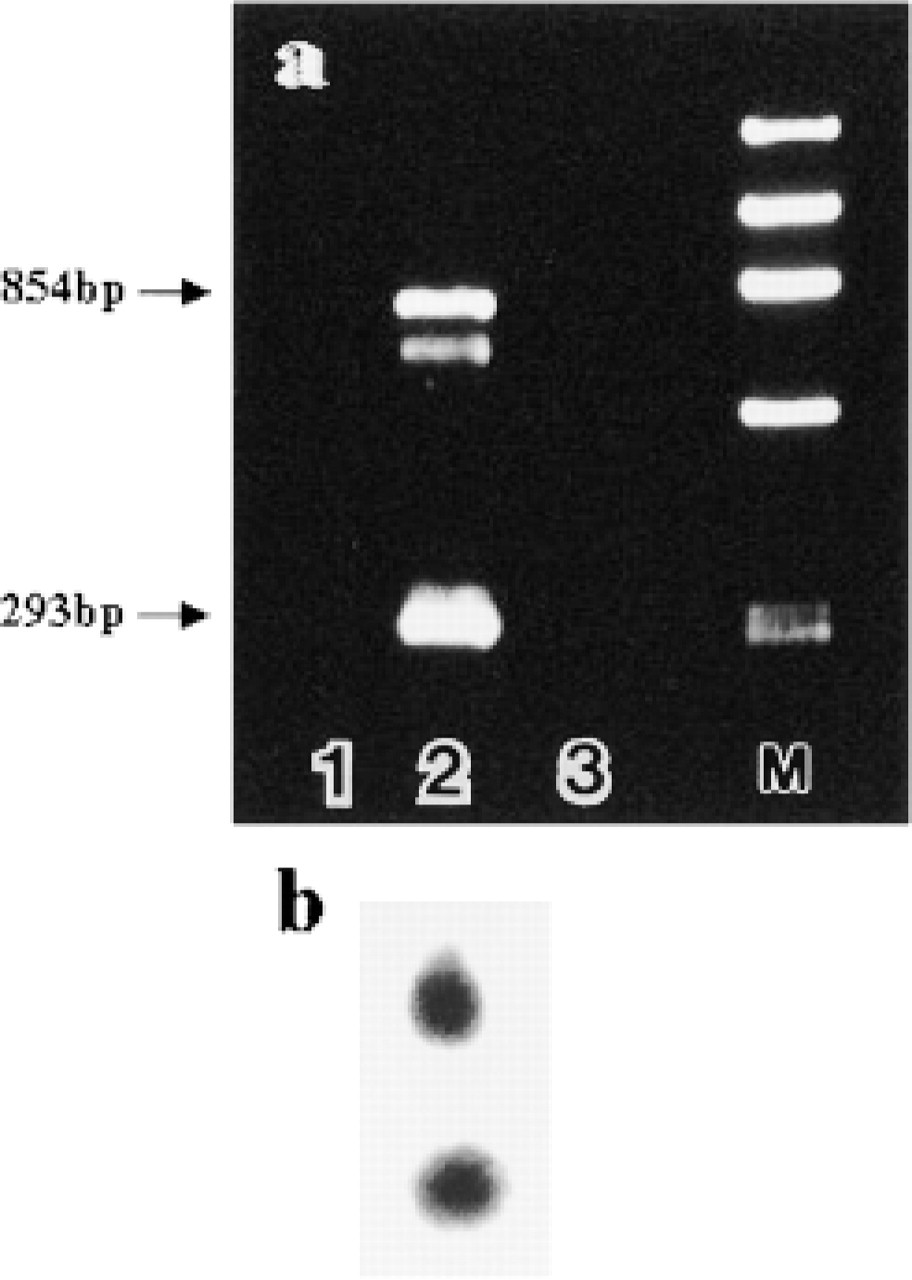
Expression of GHRH-R. (
Recently, the use of transgenic animals and genetargeting knockout animals have produced pituitary adenomas (Figures 4a-4k). In mice and rats, human GHRH transgenic techniques produced GH-secreting adenomas similar to those in human pituitary. The pituitary tumors in these transgenic animals are positive for GH, GHRH, PRL, and TSH. They also possess GHRH-R. The tumorigenic effect of GHRH through an autocrine/paracrine mechanism has been proposed. In these animals, a hyperplasia-adenoma sequence of GH cells has been shown (Umemura et al. 1995). In human pituitary adenomas, this sequence is not readily apparent because hyperplastic foci are not usually obvious. In gene targeting, p27 knockout mice are of particular interest because these animals produce intermediate-lobe tumors that are positive for POMC-derived peptides (Nakayama et al. 1996). D2 receptor knockout (D2RKO) mice exhibit PRL hyperplasia and hyperplastic intermediate lobes. The molecular mechanisms for the production of pituitary tumors in these targeted animals remains to be investigated.
Transcription Factors and Mechanisms of Specific Hormone Production
As previously mentioned, human GH- and TSH-secreting adenomas are frequently multihormonal with a combination of GH, PRL, and TSH. For the functional differentiation of these tumor cells, the pituitary transcription factor (Pit-1) and its synergistic action with other factors have been a focus of research. IHC and ISH have strongly contributed to clarification of the functional roles of the transcription factors.
In 1988, Rosenfeld et al. and Karin et al. separately reported the presence of a pituitary transcription factor/growth hormone factor 1 (Pit-1/GHF-1) (Bodner et al. 1988; Ingraham et al. 1988). Pit-1 belongs to the POU-domain family which is characterized by conservation of a 60-amino-acid homeodomain (POU-homeodomain) and a second region of 75 amino acids located near the N-terminus of the homeodomain called the POU-specific domain. The gene is mapped to human chromosome 3 and mouse chromosome 6. Pit-1 proteins, nuclear binding proteins, are 31 and 33 kD in size and are composed of 291 amino acids. Pit-1 was found in the nuclear extract of rat pituitary cells and was shown to enhance the transcription of GH and PRL (Mangalam et al. 1989). In addition, Pit-1 activated transcription of the gene for TSHβ (Li et al. 1990; Steinfelder et al. 1991, 1992) and plays a role in functional differentiation of these cells, maintenance of their viability, and proliferation (Dolle et al. 1990; Simmons et al. 1990; Castrillo et al. 1991). The DNA binding sites for Pit-1 are two sites, eight sites, and five sites upstream from GH, PRL, and TSH genes, respectively. In rodents, Pit-1 mRNA was detected in all types of pituitary cell. However, Pit-1 protein was limited to three cell types: somatotrophs, lactotrophs, and thyrotrophs (Simmons et al. 1990). The pituitaries of dwarf mice (Snell and Jackson mice) with deleted Pit-1 genes lack thyrotrophic, lactotrophic, and somatotrophic cells (Camper et al. 1990; Li et al. 1990), indicating the importance of the Pit-1 gene for the maintenance of these three cell lineages. Similarly, in humans, point mutations in the Pit-1 gene in patients with combined deficiency of GH-PRL-TSH have been reported (Phaffle et al. 1992; Radovick et al. 1992). Pit-1 iso-forms that result from alternative splicing have been recently identified: Pit-1α (wild-type), Pit-1 β (Konzak and Moore 1992), and Pit-1T (Haugen et al. 1994). It has been shown that Pit-1β is a better activator of the GH promoter than of the PRL promotor and that Pit-1T selectively activates the TSHβ promotor (Haugen et al. 1994). There have been several studies on the expression of Pit-1 in human pituitary adenomas. Overexpression of Pit-1 mRNA in GH- and PRL-secreting adenomas was observed by Friend et al. (1993), using Northern blotting, and by Pellegrini et al. (1994) using RT-PCR. In addition, Asa et al. (1993) showed by ISH Pit-1 mRNA was specifically expressed in GH- and PRL-secreting adenomas. Moreover, co-expression of Pit-1 mRNA with GH, PRL, and TSHβ in pituitary adenomas was detected by Sanno et al. (1996a) using combined ISH and IHC.
In our immunohistochemical studies on human pituitary adenomas from 60 patients, we found Pit-1 immunoreactivity in the nuclei of all cases of GH- and TSH-secreting adenomas (Sanno et al. 1996a,b) (Figures 4l-4n). Therefore, we suggest that the functional expression of these hormones in the adenoma cells resulted from DNA binding of Pit-1 protein. In PRL-secreting adenomas, Pit-1 mRNA and Pit-1 protein were found in 11/13 and 9/13 cases, respectively (Sanno et al. 1996b). It is believed that Pit-1-negative PRL-secreting adenomas may be related to other transcription mechanisms involved in functional differentiation (Sanno et al. 1996b). In an experimental model using human GHRH transgenic mice and rats, GHRH is tumorigenic in the pituitary gland, resulting in GH-secreting adenomas, in which Pit-1 functions synergistically with GHRH and GHRH-R in differentiation towards GH, PRL, and TSH (Osamura et al. 1993) (Figures 4a-4k).
It is known that synergistic factors exist for Pit-1 in the functional differentiation of pituitary cell types: GH receptor (GHRH-R), estrogen receptor (ER), thyroxine receptor (TR), retinoic acid receptor (RAR), and retinoid X receptor (RXR) (Figure 5). GHRH-R is a member of the Gs-protein-coupled receptor family and is located on chromosome 7. This protein is transmembranous with seven folds, and its molecular weight is 44–45 kD. Our ISH and RT-PCR studies showed that GHRH-R mRNA expression is common in GH- and TSH-secreting adenomas (Figures 4o and 4p). Lopes et al. (1997) also reported that GHRH-R mRNA is highly expressed in GH-secreting adenomas.
ER belongs to the nuclear receptor superfamily and its ligand is estradiol (E2). ER is located on chromosome 17 on which many oncogenes and anti-oncogenes are also situated. The ER protein includes two isoforms, α and β, with 595 amino acid residues, and a molecular size of 65 kD. In normal human anterior pituitary, ER protein is found in the nuclei and ER mRNA is found in the cytoplasm of PRL-secreting cells. In PRL-secreting adenomas, Pit-1 mRNA and ER nuclear protein co-exist in the same tumor cells, which suggests a synergistic action of Pit-1 and ER (Sanno et al. 1996c) (Figures 4q-4t). In 15 cases of PRL-secreting adenomas, Pit-1 mRNA was detected in 12 cases and three Pit-1 mRNA-negative adenomas showed positive ER protein (Sanno et al. 1996c). This suggests that PRL transcription may be alternatively regulated by ER without Pit-1.
(
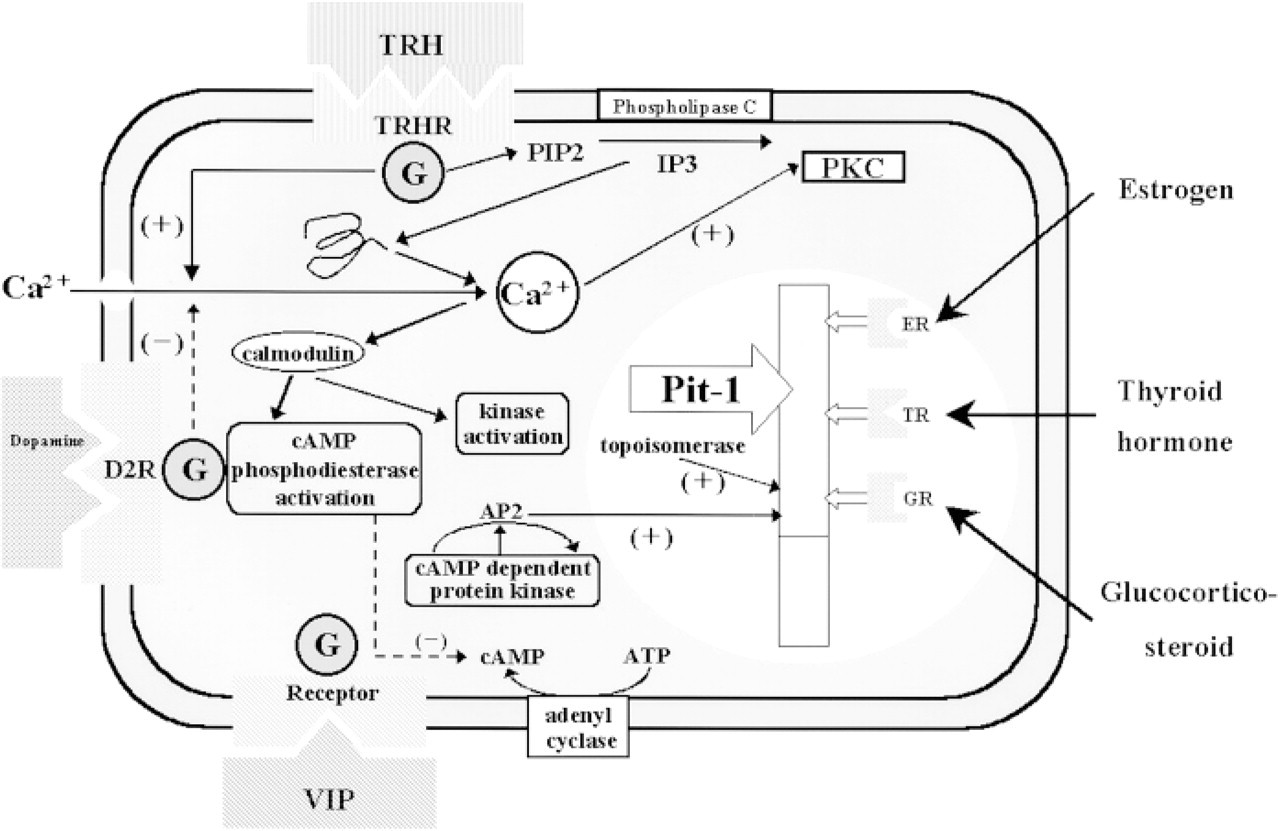
The synergy of Pit-1 and other factors in PRL expression. TRH, thyrotropin-releasing hormone; TRHR, TRH receptor; IP3, inositol 1,4,5-triphosphatase; PIP2, phosphatidyl inositol 4,5-bisphosphatase; PKC, protein kinase C; ER, estrogen receptor; TH, thyroid hormone receptor; GR, glucocorticoid receptor; VIP, vasoactive intestinal polypeptide; D2R, dopamine receptor.
RXRs are transcription factors that belong to the steroid/thyroid hormone receptor superfamily (Hamada et al. 1989; Mangelsdorf et al. 1990, 1992; Yu et al. 1991; Kliewer et al. 1992; Leid et al. 1992; Marks et al. 1992; Zhang et al 1992). These receptors are ligand-inducible, and 9′cis-retinoic acid has been identified as their ligand (Heyman et al. 1992; Levine et al. 1992; Mangelsdorf et al. 1992). RXRs are located on chromosome 17 and three isoforms are known: RXRα, β, and γ. RXRα, β, and γ proteins consist of 446, 448, and 463 amino acid residues, respectively. RXRs bind to the hormone-responsive elements of specific genes and form homodimers (Levine et al. 1992) as well as heterodimers with other receptors, including thyroxine receptors (TRs), retinoic acid receptors (RARs), and vitamin D receptors (Berrodin et al. 1992; Kliewer et al. 1992; Marks et al. 1992; Zhang et al. 1992; Davis et al. 1994). For the rat GH gene, it has been suggested that endogenous RXRs can function as hormone receptors that regulate the transcription of GH (Allenby et al. 1993; Davis et al. 1994; Sugawara et al. 1994). Signaling crosstalk between RXRs and other receptors, including TRs, RARs, vitamin D receptors, and ER, has been also reported (Berrodin et al. 1992; Mano et al. 1994; Keller et al. 1995). In the three known isoforms of the RXRs, RXRα and β mRNAs are widely expressed (Mangelsdorf et al. 1990, 1992; Yu et al. 1991). In contrast, RXRγ mRNA is restricted to tissues such as skeletal muscle, heart, brain, and fetal pituitary (Mangelsdorf et al. 1992; Allenby et al. 1993). Sanno et al. (1997) have reported the immunohistochemical expression of RXRs isoforms in human non-neoplastic pituitaries and pituitary adenomas using isoform-specific polyclonal anti-RXR antibodies. In normal human pituitary gland, RXRα was observed in all types of human anterior pituitary cells but RXRγ was mainly localized in TSHβ-, GH-, and α-SU-positive cells by double immunohistochemical staining. In all TSH-secreting adenomas, most adenoma cells showed Pit-1 mRNA in the cytoplasm and RXRγ protein in the nuclei. Therefore, RXRγ may promote the functional differentiation of TSH as indicated by the TSH-secreting adenomas.
Other Transcription Factors
Recently, several transcription factors relative to pituitary development have been discovered. Transcription factors transiently expressed during pituitary development include the prophet of Pit-1 (Prop-1) and Rpx/Hesx-1 (Thomas et al. 1995; Hermesz et al. 1996; Sornson et al. 1996). It is known that Prop-1 acts as an early enhancer of Pit-1. The Prop-1 gene is located on chromosome 11 and its anticipated protein consists of 223 amino acids. Prop-1 is expressed before Pit-1 from embryonic Day 10–10.5 (e10-10.5), and expression of Prop-1 peaks on e12. Pit-1 is first expressed from e13.5, and expression of Prop-1 decreases as the expression of Pit-1 increases (Sornson et al. 1996). It has been debatable whether or not Prop-1 is expressed in mature pituitary gland, although it was initially reported that Prop-1 expression is not present in the mature gland. Rpx/Hesx-1, a homeobox gene, is also expressed transiently from e9 to 14.5 during differentiation of the pituitary gland, although its function is not yet fully clarified (Hermesz et al. 1996). Rpx/Hesx-1 has been reported to form a heterodimer with Prop-1 and to inhibit Pit-1 activation by Prop-1 (Sornson et al. 1996), which was inferred from the finding that Rpx/Hesx-1 was expressed throughout the embryonic period in Prop-1-deficient mice (Gage et al. 1996; Sornson et al. 1996). Because it has also been reported that pituitary hypoplasia occurs in transgenic mice with Rpx/Hesx-1 expression throughout the embryonic period, the disappearance of Rpx/Hesx-1 appears to be necessary for proper differentiation of the various types of pituitary cells (Tremblay et al. 1998). Therefore, activation of Pit-1 by Prop-1 may be triggered by the disappearance of Rpx/Hesx-1.
Pituitary homeobox 1 (Ptx1) is a transcription factor first reported by Lamonerie et al. (1996), which has been suggested to play a role in the transcription of POMC because it specifically binds to the nucleus of AtT-20 cells, a murine corticotrophic cell line. Ptx1 is closely related to the mammalian Otx gene, which is expressed in the rostral brain during development (Lala et al. 1992; Morohashi et al. 1992), and Otx2 is a fundamental factor related to forebrain and midbrain development in mice (Honda et al. 1993). Ptx1 mRNA is strongly expressed in adult corticotrophic cells, which express the POMC gene, and Ptx1 expression in Rathke's pouch has also been detected by ISH (Lamonerie et al. 1996). Therefore, the possibility remains that Ptx1 is expressed in pituitary cells other than corticotrophs. The relationship of Ptx1 to other transcription factors related to pituitary cells needs to be investigated.
Recent investigations on cultured cells have shown that Ptx1 is involved not only with POMC transcription in corticotrophs but also with the transcription of other anterior pituitary hormones. Tremblay et al. (1998) investigated Ptx1 expression in AtT-20 cells, a corticotrophic cell line used for Ptx1 cloning, and in αT3-1 cells (gonadotroph precursor), αTSH cells (thyrotroph precursor), GHFT1.5 cells (somatolactotroph precursor), GH3 cells (a somatolactotrophic cell line), and TtT-97 cells (thyrotrophic tumor) These authors showed by Northern blotting that the Ptx1 mRNA level was higher in αT3-1, αTSH, and GHFT1.5 cells than in AtT-20 cells, indicating that Ptx1 may be actively expressed in pituitary cells other than corticotrophs. They also investigated cultured cells by Western blotting and showed that there was no discrepancy between Ptx1 protein expression and Ptx1 mRNA expression (Tremblay et al. 1998). Furthermore, Acampora et al. (1998) reported that Otx1 is a pituitary transcription factor involved at the prepubescent stage in the control of GH, FSH, and LH levels. Their data suggest that a complex regulatory mechanism exists to control the physiological need for pituitary hormones at specific postnatal stages. We also detected Ptx1 protein in the nuclei of all types of anterior pituitary hormone-secreting cells by IHC, indicating that Ptx1 is a nuclear binding protein. This protein was also observed with variable frequency in all types of pituitary adenoma. It appears that Ptx1 has no hormone-related specificity and is involved as a basic factor in transcription of all types of anterior pituitary cells. Synergistic actions of other transcription factors, such as Pit-1, SF-1, Neuro D, and Dax1, have been proposed (Figure 6).
SF-1/Ad4BP is a transcription factor that regulates the P-450 gene (Lala et al. 1992; Morohashi et al. 1992, 1993; Honda et al. 1993), which plays a role in steroid hormone biosynthesis and belongs to the steroid hormone receptor supergene family containing a Zn-finger domain in the DNA binding region (Evans 1988). RT-PCR has shown that this factor is expressed not only in the adrenal glands but also in the pituitary and the hypothalamus (Honda et al. 1993). In nontumorous pituitary cells, expression of SF-1/Ad4BP with FSHβ was detected by IHC (Ingraham et al. 1994). A remarkable decrease in the numbers of gonadotrophs has been observed in Ad4BP knockout mice (Luo et al. 1994), suggesting involvement of SF-1/Ad4BP in transcriptional activity in gonadotrophs. Asa and co-workers (1996) investigated the expression of SF-1/Ad4BP in human pituitary adenomas using IHC and RT-PCR. By RT-PCR, SF-1/Ad4BP mRNA was found in the pituitary adenomas expressing gonadotrophs. By IHC, SF-1/Ad4BP protein was localized in the nuclei of scattered gonadotrophic cells of the anterior pituitary. In gonadotropin subunit-positive adenomas, SF-1/Ad4BP protein was diffusely expressed in the nuclei of the adenoma cells. Furthermore, Ptx1 has been reported to be in synergy with SF-1/Ad4BP to enhance the promoter activity of LHβ (Tremblay et al. 1998).
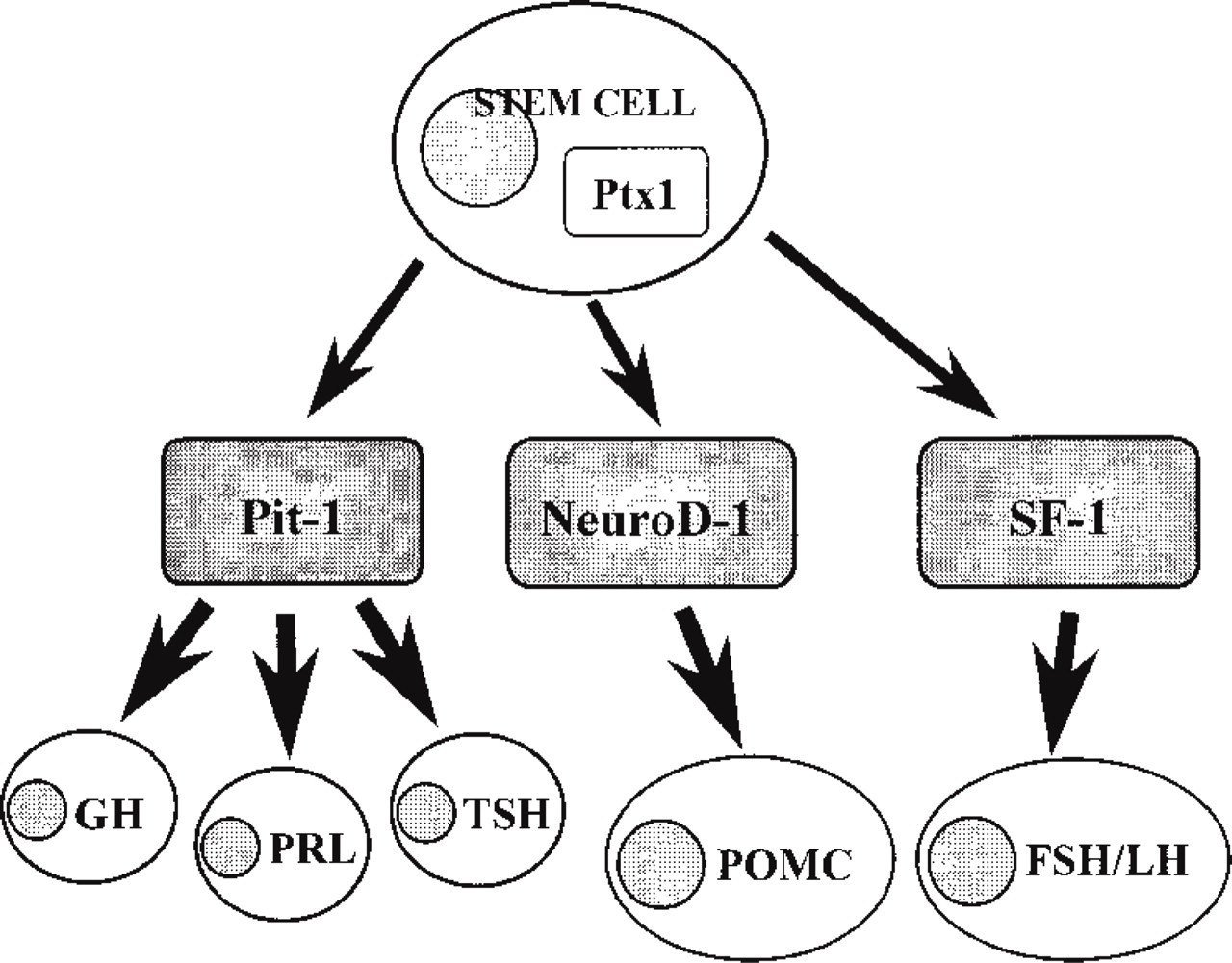
Schematic drawing of functional differentiation of anterior pituitary cells and synergistic action of Ptx1 with other transcription factors.
NeuroD1/β2 is a transcription factor containing a helix-loop-helix heterodimer (Lee et al. 1995). It was first found in pancreatic islet cells and is also known to be a transcription factor specific for insulin-secreting cells (Naya et al. 1995), and it has also been detected in some nerve cells. In the pituitary, synergistic action between NeuroD1/β2 and Ptx1 has also been reported, particularly Ptx1 synergy with NeuroD1/β2 to activate POMC transcription (Therrien and Drouin 1993; Poulin et al. 1997).
P-Lim/Lhx-3 mRNA initially appears in Rathke's pouch on e8.5–9, and it is known that four types of anterior pituitary cells, including α-SU, are completely depleted in P-Lim/Lhx-3-deficient knockout mice (Sheng et al. 1996). An experiment using a Ptx1 knockout cell line showed that expression of the Lhx-3 gene was markedly decreased compared with that of the α-SU gene (Tremblay et al. 1998). This indicates that Ptx1 is essential for Lhx-3 gene expression. Evidence has been accumulated on the involvement of Ptx1 in the transcription of α-SU, and P-Lim/Lhx-3 appears to be involved in α-SU transcription, as a co-factor of Ptx1 (Bach et al. 1997).
Processing of Prohormones
It is well known that peptide hormones are produced as functionally inactive prohormones that are subjected to post-translational modification and then secreted as biologically active forms (Chretien and Li 1967; Steiner et al. 1967). These post-translational modifications include proteolytic digestion of dibasic amino acid residues (Chance et al. 1968) and amidation. The KEX 2 family, which is the processing endopeptidase, includes seven different kinds of proteins. Prohormone convertase (PC) 3 was later indicated to be the same as PC 1 and now is designated as PC1/3. In endocrine tumors, the functions of PC1/3 and PC2, which are present in the neuroendocrine tissues, have been the subject of several investigations (Bloomquist et al. 1991; Ohagi et al. 1992; Seidah and Chretien 1992; Hashimoto et al. 1994; Scopsi et al. 1995; Lloyd et al. 1995). Morphological studies have been done in pancreatic and pituitary tumors (Lloyd et al. 1995; Otoh et al. 1996). In the pancreatic islets, PC1/3 and PC2 function to produce insulin and C-peptide. POMC is cleaved to ACTH by PC1/3 and further to αMSH by PC2 (Figure 7). Lloyd et al. (1995) reported the presence of PC1/3 and PC2 mRNA in a variety of endocrine tumors, including pituitary adenomas. Takumi et al. (1998) studied immunohistochemical expression of PC1/3 and PC2 in various pituitary adenomas. In these cases, 72% and 34% of pituitary adenomas expressed PC1/3 and PC2, respectively. The incidence of PC1/3-positive cases was higher in ACTH-secreting adenomas, TSH-secreting adenomas, and nonfunctioning adenomas. On the other hand, the incidence of PC2-positive cases was higher in nonfunctioning adenomas and ACTH-secreting adenomas. In ACTH-secreting adenomas, ACTH and αMCH were localized to the same tumor cells in which PC1/3 and PC2 were expressed (Takumi et al. 1998). Confocal laser scanning microscopy (CLSM) showed a granular pattern suggesting that processing occurs in the secretory granules. This was demonstrated by immunoelectron microscopy in rat pancreatic islet cells (Itoh et al. 1996).
Secretory Granule-related Proteins
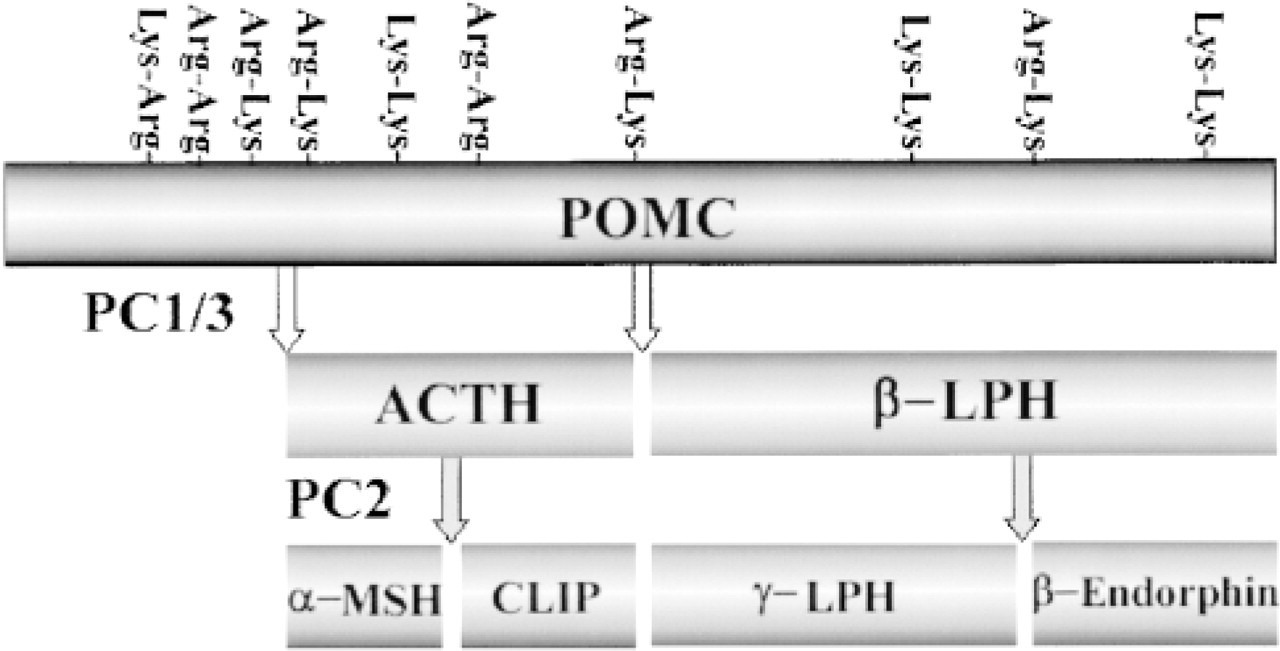
Processing of pro-opiomelanocortin(POMC) by prohormone convertase (PC)1/3 and PC2.
Secretion is the phenomenon by which the cell synthesizes and concentrates bioactive substances and, after a certain period of storage, releases them in response to external stimulation. This process can be classified as constitutive secretion, or regulated secretion, depending on the involvement of secretory granules. In regulated secretion, secretory proteins are packaged in secretory granules, transferred to the plasma membrane, and then released in response to extracellular stimulation. This process is observed in neuropeptide and peptide hormone-secreting cells. The granin family, including chromogranins A and B (CgA and B), plays an important role in the packaging of these substances into secretory granules (Rosa et al. 1985). The process has been extensively studied in pituitary cell and in various endocrine tumors, including human pituitary adenomas (Destephano et al. 1988; Osamura et al. 1988; Stefaneanu et al. 1988; Lloyd et al. 1989; Lloyd and Jin 1994, 1995). In human pituitary adenomas, expression of CgA was found in glycoprotein hormone-producing adenomas but not in PRL-secreting adenomas, whereas by IHC and ISH CgB was expressed in PRL-secreting adenomas (Lloyd et al. 1989, 1990, 1991).
At present, well-known Ras oncogene products are believed to be low molecular weight GTP binding proteins that serve as a “switch” for signaling cell proliferation and differentiation. These Ras-related proteins, the Rab family, are considered to be involved in regulation of vesicular transport in the cell. At present, about 30 proteins have been identified in this family. Among these, Rab3 has been reported to be involved in the release of neurotransmitters from nerve cell synapses and in the exocytosis of secretory granules from endocrine cells, including those of the anterior pituitary (Figure 8). These functions of Rab3 were first proposed by Bourne (1988). GDP-Rab3, the inactive form, is bound to Rab3-GDI (Rab3-GDP dissociation inhibitor). When the inactive form is transformed into the active form, GTP-Rab3, Rab3 is bound to secretory granules, and is transferred to and fuses with the plasma membrane to cause Ca++-dependent exocytosis. Then Rab3 is once again converted to the inactive form. This cyclic process is considered to regulate the secretion of hormones from secretory granules.
In our study, the Rab3 protein was extensively expressed as the granular form in the cytoplasm of human anterior pituitary hormone-secreting cells. It was not expressed in S-100-positive FS cells, which do not contain secretory granules in their cytoplasm (Tahara et al. 1999) (Figures 4u-4z). Immunoelectron microscopic studies on human pituitary adenomas localized this protein to the limiting membrane of the secretory granules in adenoma cells. These findings provide morphological evidence for the involvement of Rab3 in hormone secretion from secretory granules in pituitary adenomas. In particular, Rab3 was expressed in 100% of GH-secreting adenomas which are usually densely granulated adenomas. In adenomas having fewer secretory granules indicating a Golgi pattern, such as PRL-secreting adenomas, the Rab3 protein was expressed in as few as 33.3% of adenomas. This observation and the results of immunoelectron microscopic studies suggest that the expression of Rab3 is correlated with the number of secretory granules (Tahara et al. 1999).
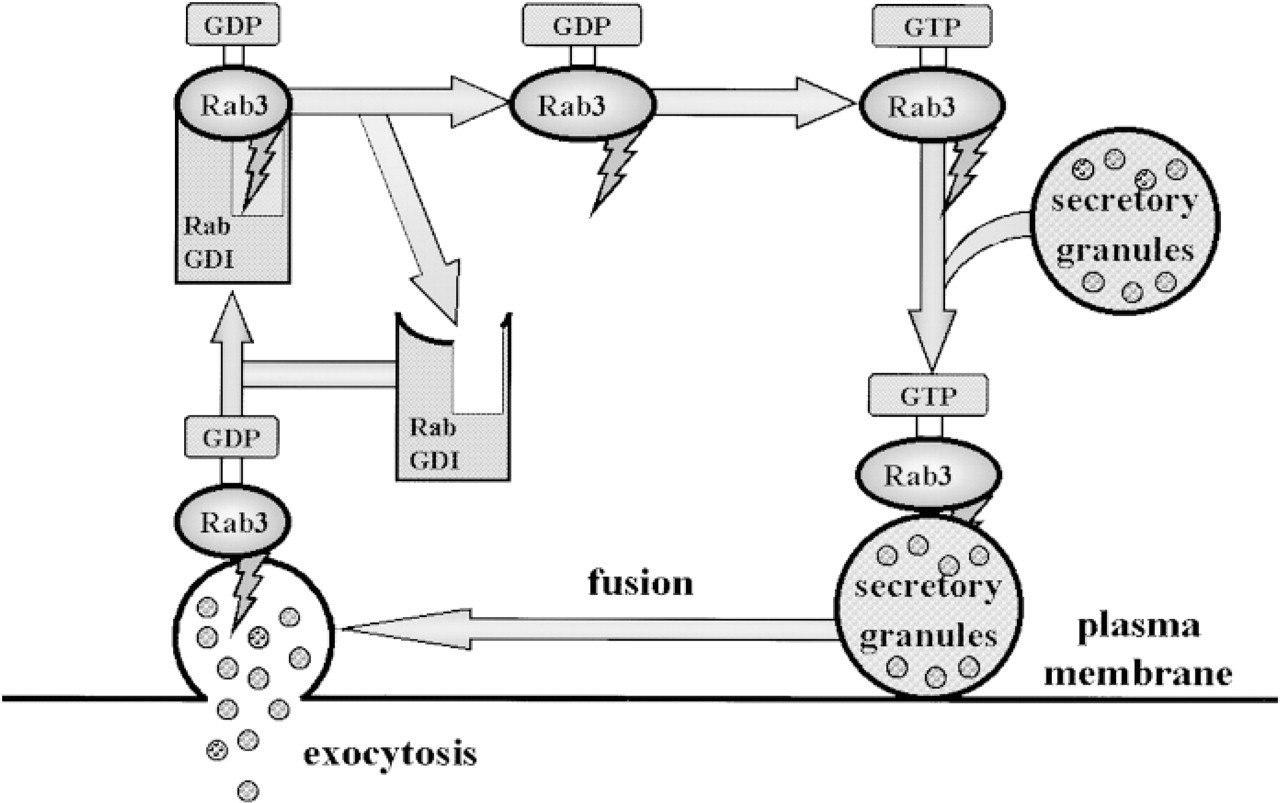
Functional role of Rab3 protein in secretory pathway of secretory granules.
Four types of isoforms, Rab3A, 3B, 3C, and 3D, are known for Rab3 protein and these have homology of as high as 77–85% (Zahraoui et al. 1989; Fischer von Mollard et al. 1994b). Rab3A is localized mainly in synaptic vesicles of the central nervous system and is currently most extensively studied for its regulation of neurotransmitter release (Mizoguchi et al. 1990; Fischer von Mollard et al. 1990, 1991; Matteoli et al. 1991; Geppert et al. 1994). Rab3C and Rab3A are also co-expressed in synaptic vesicles of nerve cells (Fischer von Mollard et al. 1994a,b). Rab3B is known to be involved in exocytosis of secretory granules of anterior pituitary cells (Lledo et al. 1993), and Rab3D was observed to be expressed mainly on adipocytes (Baldini et al. 1992). As mentioned above, of the Rab3 isoforms, Rab3B has attracted attention because it is believed to hold the important key, and this has been elucidated in studies by Lledo et al. (1993). When a trace amount of antisense oligonucleotide of Rab3B was injected into anterior pituitary cells, the expression of Rab3B Ca++-dependent exocytosis was inhibited. On the other hand, a similar study performed using Rab3A showed no alteration, suggesting that Rab3B promotes hormone secretion by anterior pituitary cells. Another study has indicated that Rab3B is involved in basal and gonadotropin-releasing hormone-induced gonadotropin release (Tasaka et al. 1998). In our study, immunoelectron microscopy revealed that Rab3 was localized along the limiting membrane of secretory granules in Rab3-positive, gonadotropin subunit-positive adenomas. These results suggested that the Rab3 isoforms, especially Rab3B, were involved in gonadotropin secretion by human pituitaries under neoplastic conditions. However, recent studies have shown that Rab3A was expressed, although weakly, on mammalian anterior pituitary cells (Redecker et al. 1995; Aguado et al. 1996), and a study with ISH demonstrated that Rab3A mRNA, in addition to that of Rab3B, was expressed weakly (Stettler et al. 1995). Rab3A has been reported to inhibit GH secretion in chromaffin cells into which the GH gene was transfected (Holz et al. 1995), and also had an inhibitory effect on exocytosis by other neuroendocrine cells (Holz et al. 1994; Johannes et al. 1994).
At present, proteins that have been shown to be involved in exocytosis of secretory granules from endocrine cells include synaptotagmin, SNAP-25, syntaxin, and synaptobrevin/VAMP, in addition to the Rab3 isoform. Synaptotagmin possesses nine isoforms (Mizuta et al. 1994; Craxton and Goedert 1995; Li et al. 1995; Marqueze et al. 1995). One of these isoforms, synaptotagmin I, has been reported to be found in somatotrophs, corticotrophs, thyrotrophs, and gonadotrophs of rat pituitary cells but was not observed in lactotrophs (Redecker et al. 1995). SNAP-25, syntaxin, and synaptobrevin/VAMP are proteins involved in exocytosis by synaptic vesicles in the central nervous system (Thomas and Betz 1990). SNAP-25 and syntaxin have been shown to be present on the presynaptic membrane, and synaptobrevin/VAMP on the synaptic vesicle membrane. These three proteins have been reported to be expressed in rat pituitary cells, and SNAP-25 has been shown by IHC to be co-expressed with gonadotrophic cells (Aguado et al. 1996). Elucidation of the role of these proteins in pituitary adenomas and their synergism with the Rab3 isoform remains a future important subject to be investigated.