
Abstract
We examined the expression and cytolocalization of the protein phosphatase type 1 delta (PP1δ) isoform and nucleolin in human osteoblastic MG63 and Saos-2 cells. Cellular fractionation of MG63 cells was done and protein was prepared from each fraction. Anti-nucleolin antibody interacted with the 100- and 95-kD proteins present in the whole-cell lysate. The 100-kD protein was detected in nuclear and nucleolar fractions. The 95-kD protein was detected in cytosolic and nucleoplasmic fractions. PP1δ and nucleolin were co-localized in the nucleolus in MG63 and Saos-2 cells revealed by an immunofluorescence method. PP1δ and nucleolin were also co-immunoprecipitated with anti-nucleolin and anti-PP1δ antibodies. In the actinomycin D-treated cells, the subcellular localization of PP1δ and nucleolin was changed. Expression of PP1δ was upregulated with actinomycin D treatment. The level of 100-kD protein did not change in the actinomycin D-treated cells. However, the level of the 95-kD band increased with actinomycin D treatment. These results indicate that PP1δ was associated with nucleolin in the nucleolus of MG63 and Saos-2 cells and that nucleolin is a possible candidate substrate for PP1δ.
P
The expression of PP1s was previously investigated at the mRNA and protein levels in various tissues and organs (Shima et al. 1993c). Immunocytochemistry was also carried out in rat cerebellum and salivary glands for PP1γ1 (Hashikawa et al. 1993; Shirakawa et al. 1996) and in testis for PP1γ2 (Shima et al. 1993a). In a previous study, we reported the subcellular localization of PP1 isoforms in mouse osteoblastic MC3T3-E1 cells (Haneji et al. 1998). PP1-targeting subunits are believed to direct PP1 to specific subcellular components and to modulate the activity of the enzyme at these sites.
Nucleolin has a molecular mass of 100-110 kD and is an abundantly expressed phosphoprotein located mainly in the fibrillar components of nucleoli, where it associates with nascent preribosomal RNA. Nucleolin is involved in the regulation of ribosome biogenesis, cell proliferation and growth, embryogenesis, cytokinesis, and nucleogenesis (Ginisty et al. 1999; Srivastava and Pollard 1999; Derenzini 2000). Nucleolin operates as a shuttle between the cytoplasm and the nucleus (Borer et al. 1989) and is involved in the hypercondensation of chromatin that occurs during mitosis (Erard et al. 1988). Nucleolin is a substrate for several kinases, including casein kinase II, p34cdc2, and protein kinase C-ζ (Schneider et al. 1989; Zhou et al. 1997). In the present study we provide evidence showing that nucleolin is associated with PP1δ and is a candidate substrate for this enzyme.
Materials and Methods
Materials
Alpha-modified Eagle's minimal essential medium was purchased from Gibco (Grand Island, NY). Fetal bovine serum (FBS) was obtained from JRH Biosciences (Lenexa, KS). Plastic dishes were from Falcon Plastics (Los Angeles, CA). Anti-PP1δ antiserum was a gift from Drs. M. Nagao and H. Shima and was characterized previously (Shima et al. 1993c). Immunoglobulin G fraction of an anti-nucleolin monoclonal antibody (clone 4E2) was purchased from MBL (Nagoya, Japan). Actinomycin D was purchased from Sigma Chemical (St Louis, MO). Stock solution (10 mM) was prepared in methanol and added to the culture media.
Cell Culture
Human osteoblastic cell lines MG63 (Franceschi et al. 1985) and Saos-2 (Rodan et al. 1987) were purchased from the American Type Culture Collection (Rockville, MD). The cells were cultured in α-MEM containing 10% (v/v) FBS, 2 mM glutamine, 100 U/ml penicillin, and 100 μg/ml streptomycin and were maintained at 37C in a humidified atmosphere of 5% CO2 and 95% air. The medium was changed every 3 days and subculturing was performed every 6 days by treatment of the cells with 0.25% trypsin together with 1 mM EDTA in Ca2+-, Mg2+-free PBS. For experiments, sub-confluent cells were treated with the agents by adding small volumes of stock solution to the medium and culturing for appropriate periods. Cell modification was monitored by an Olympus IMT-2 phase-contrast microscope. For immunocytochemistry, the cells were plated on 18-mm round coverslips in 60-mm plastic dishes.
Cell Fractionation, SDS-PAGE, and Western Blotting
Cells cultured in 90-mm plastic dishes were washed twice with PBS, scraped into PBS, pelleted at 3000 × g, and resuspended in hypotonic buffer [20 mM Hepes (pH 7.2), 10 mM KCl, 1 mM MgC12, 1 mM DTT, 0.5 mM EDTA, 50 mM NaF, and 1 mM Na3VO4]. The cells were allowed to swell for 10 min on ice before lysis by addition of 0.1% NP-40 and 100 mM potassium acetate. After 5 min on ice and vortexing, their nuclei were pelleted by centrifugation for 10 min at 8000 × g, resuspended in lysate buffer containing 1 mM DTT, 1 mM PMSF, 1 μg/ml leupeptin, 2 μg/ml aprotinin, and 5 mM EGTA in PBS, and used as the nuclear fraction, whereas the supernatant was taken as the cytosolic fraction. The nucleolar and nucleoplasmic fractions were prepared from purified nuclei according to the method of Muramatsu et al. (1974). The protein concentration of each fraction was evaluated by using Protein Assay Reagent (Bio-Rad; Hercules, CA) and was diluted to a protein concentration of 1 mg/ml with lysate buffer before the addition of Laemmli's 5 × sample buffer (Laemmli 1970). Twelve μg of each sample and prestained molecular weight markers (GIBCO BRL; Gaithersburg, MD) were separated by SDS-PAGE and transferred electrophoretically to polyvinylidene difluoride (PVDF) transfer membranes (Immobilon; Millipore, Bedford, MA) according to the procedure of Towbin et al. (1979). The membranes were incubated in a blocking solution containing 5% non-fat skim milk in PBS-Tween for 2 hr at 20-22C. Then they were washed briefly in PBS-Tween and incubated overnight at 4C in PBS-Tween containing anti-PP1δ antibody diluted 1:2000 or 0.5 μg/ml of anti-nucleolin antibody. The membranes were washed four times within 30 min in PBS-Tween on a rotary shaker at 20-22C. The washed membranes were incubated in PBS-Tween containing horseradish perioxidase (HRP)-conjugated anti-rabbit IgG or anti-mouse IgG (diluted 1:10,000) for 1 hr at 20-22C. The membranes were washed as described and the proteins recognized by the anitbodies were visualized by using an ECL detection kit (Amersham Pharmacia Biotech; Uppsala, Sweden) according to the manufacturer's directions.
Immunoprecipitation
Cells cultured in 90-mm plastic dishes were washed twice with PBS, scraped into PBS, pelleted at 3000 × g, and resuspended in 500 μl of lysis buffer [150 mM Nacl, 1.0% NP-40, 50 mM Tris-HCl (pH 8.0), 50 mM NaF, and 1 mM Na3 VO4]. The lysate was preincubated for 30 min with 20 μl of protein A/G PLUS agarose (Santa Cruz Biotechnology; Santa Cruz, CA) and then incubated with 2 μl of anti-PP1δ antiserum or 1 μl of anti-nucleolin antibody conjugated to 10 μl of protein A/G PLUS agarose. The reaction mixture was incubated for 6 hr at 4C. The immunocomplexes were washed four times with lysis buffer and resuspended in 40 μl of SDS electrophoresis sample buffer. The samples were boiled for 3 min and analyzed by SDS-PAGE and Western blotting using the anti-nucleolin or anti-PP1δ antibody.
Immunocytochemistry
The cells on the coverslips were washed three times with PBS and fixed with 3.7% formaldehyde for 10 min at 20-22C, followed by methanol permeabilization for an additional 20 min at −20C. Nonspecific binding sites were blocked with 4% BSA in PBS for 10 min at 20-22C in a humidified atmosphere. Having been rinsed with cold PBS, the coverslips were incubated simultaneously with anti-PP1δ antibody diluted 1:200 and 5 μg/ml of the IgG fraction of anti-nucleolin antibody in 4% BSA for 45 min at 20-22C. After three washes with 0.1% BSA in PBS containing 0.05% Tween-20 (PBS-Tween) over a 15 min period at 20-22C, the cells were incubated with a mixture of tetramethylrhodamine isothiocyanate (TRITC, rhodamine)-conjugated sheep anti-rabbit IgG (Chemicon International; Temecula, CA) and fluorescein isothiocyanate (FITC)-conjugated goat anti-mouse IgG (Cappel-Organon Teknika; Turnhout, Belgium), both diluted 1:300 in 4% BSA in PBS for another 45 min at 20-22C. The coverslips were washed as described above and mounted while wet with PermaFluor aqueous mounting medium (Lipshow; Pittsburgh, PA). The samples were examined under an Olympus BX50 microscope equipped with epifluorescence illumination (BX-FLA) with a U-MWIG filter for rhodamine and a U-MNIBA filter for FITC. The U-MNIBA filter separates FITC from rhodamine or Texas Red. The staining reaction was not observed when FITC-labeled cells were examined with a filter for rhodamine (a U-MWIG filter). Rhodamine-labeled cells were not detected with a filter for FITC (a U-MNIBA filter). Photomicrographs were taken on Fuji Sensia 400 film using the automatic exposure (PM-30).
Results
Expression of PP1δ and Nucleolin in MG63 Cells
Figure 1 shows that the anti-PP1δ antibody interacted with a band corresponding to the 37-kD protein present in the nuclear and nucleolar fractions prepared from MG63 cells. However, very few immunoreactive proteins were detected in the cytosolic fraction. The anti-nucleolin antibody interacted with the 100- and 95-kD proteins present in the whole cell lystate. The antibody recognized the 100-kD protein in nuclear and nucleolar fractions, and the staining intensity was stronger in the latter. Normal rabbit serum and normal mouse serum did not recognize any proteins prepared from cultured MG63 cells (data not shown).
Co-localization of PP1δ and Nucleolin in MG63 and Saos-2 Cells
In previous reports, we and others demonstrated that PP1δ localized in the nucleolus in mouse osteoblastic MC3T3-E1 cells and Swiss 3T3 cells (Haneji et al. 1998; Kotani et al. 1998). This distribution pattern was similar to that of nucleolin. To examine whether PP1δ and nucleolin would be localized in the same site in the nucleolus, we fixed, permeabilized, and stained cultured MG63 and Saos-2 cells with the anti-PP1δ polyclonal antibody and the anti-nucleolin monoclonal antibody. Figure 2 shows the distribution of PP1δ and nucleolin in MG63 and Saos-2 cells. With the anti-PP1δ antibody, nucleolus-like bodies were intensely stained and visible as red fluorescence due to the rhodamine-conjugated second antibody (Figure 2a). A monoclonal anti-nucleolin antibody stained the same sites of the PP1δ-positive regions, which gave a green fluorescence because an FITC-conjugated second antibody was used (Figure 2b). The merged view confirmed that PP1δ and nucleolin were localized in the same sites in MG63 cells because the reaction was visible as yellow fluorescence (Figure 2c). Figure 2 also shows that the anti-PP1δ antibody and the anti-nucleolin antibody stained the nucleolus-like materials in Saos-2 cells (Figures 2d and 2e). The positive sites were stained as yellow fluorescence in the merged view, indicating that PP1δ and nucleolin were also localized in the same sites in Saos-2 cells (Figure 2f). The staining reaction was not observed when normal rabbit serum or normal mouse serum was used as a primary antibody in both MG63 cells and Saos-2 cells (data not shown).
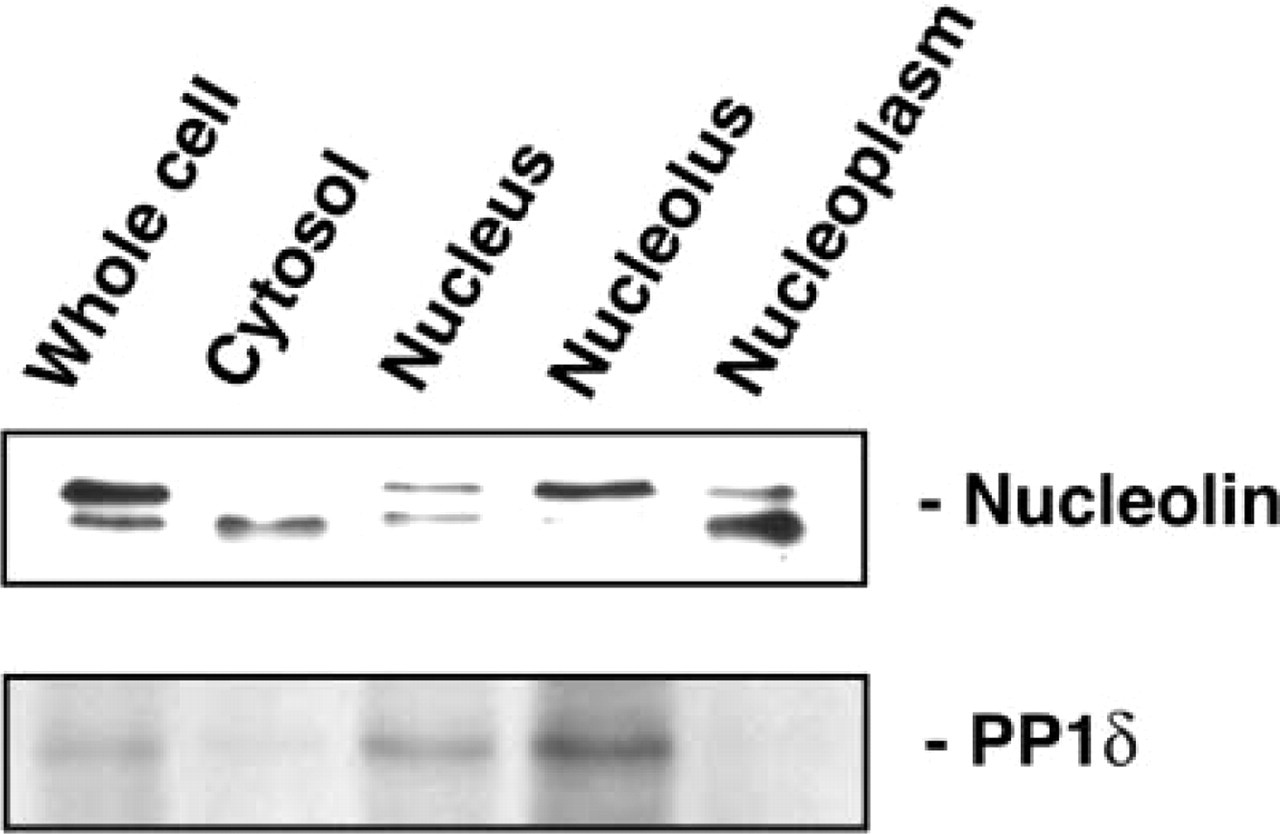
Identification of PP1δ and nucleolin in cell fractions from cultured MG63 cells. The cultured cells were fractionated and proteins were obtained from whole-cell lysate, cytosolic, nuclear, nucleolar, and nucleoplasmic fractions as described in the text. Twelve μg of each fraction was separated on a 12.5% SDS-PAGE, transferred to a PVDF membrane, and incubated with each antibody.
PP1δ Associates with Nucleolin
To determine whether PP1δ could associate with nucleolin, the whole cell lysate prepared from cultured MG63 cells was immunoprecipitated with the anti-PP1δ antibody or with the anti-nucleolin antibody. The immunoprecipitants were analyzed by Western blotting using the anti-nucleolin antibody (Figure 3a) or the anti-PP1δ antibody (Figure 3b). The anti-nucleolin antibody interacted with a protein precipitated with the anti-PP1δ antibody. However, this antibody did not recognize any proteins precipitated with normal rabbit serum (Figure 3a). The molecular weight of the interactive protein was estimated as 100 kD, corresponding to the molecular weight of nucleolin. The anti-PP1δ antibody interacted with a protein precipitated with the anti-nucleolin antibody. The interacted protein was estimated as 37 kD, corresponding to PP1δ. The anti-PP1δ antibody did not interact with the proteins precipitated with normal mouse serum (Figure 3b). As positive controls, the results for whole cell lysate stained with the anti-nucleolin antibody or anti-PP1δ antibody are also shown in Figures 3a and 3b, respectively.
Alteration of PP1δ and Nucleolin in Actinomycin D-treated MG63 Cells
The changes of PP1δ and nucleolin expression in the actinomycin D-treated MG63 cells were examined by histochemical and Western blotting procedures. With the PP1δ antibody, the positive bodies became smaller, increased in number, and were distributed over the nuclei instead of being located in nucleoli (Figure 4a). The anti-nucleolin antibody also recognized the same bodies as nucleolin-containing materials and was distributed all over the nuclei (Figure 4b). The merged view confirmed that PP1δ and nucleolin were distributed in the same bodies in the nuclei, as these bodies appeared as yellow fluorescence (Figure 4c). Figure 5 shows the results obtained from Western blotting analysis for nucleolin and PP1δ in the actinomycin D-treated cells. The amount of 100-kD nucleolin did not change in the actinomycin D-treated cells. However, the intensity of the 95-kD band increased in the actinomycin D-treated cells in a dose-dependent manner up to 10 nM (Figure 5, upper panel). The staining intensity of PP1δ in the actinomycin D-treated cells also increased in a dose-dependent manner (Figure 5, lower panel).
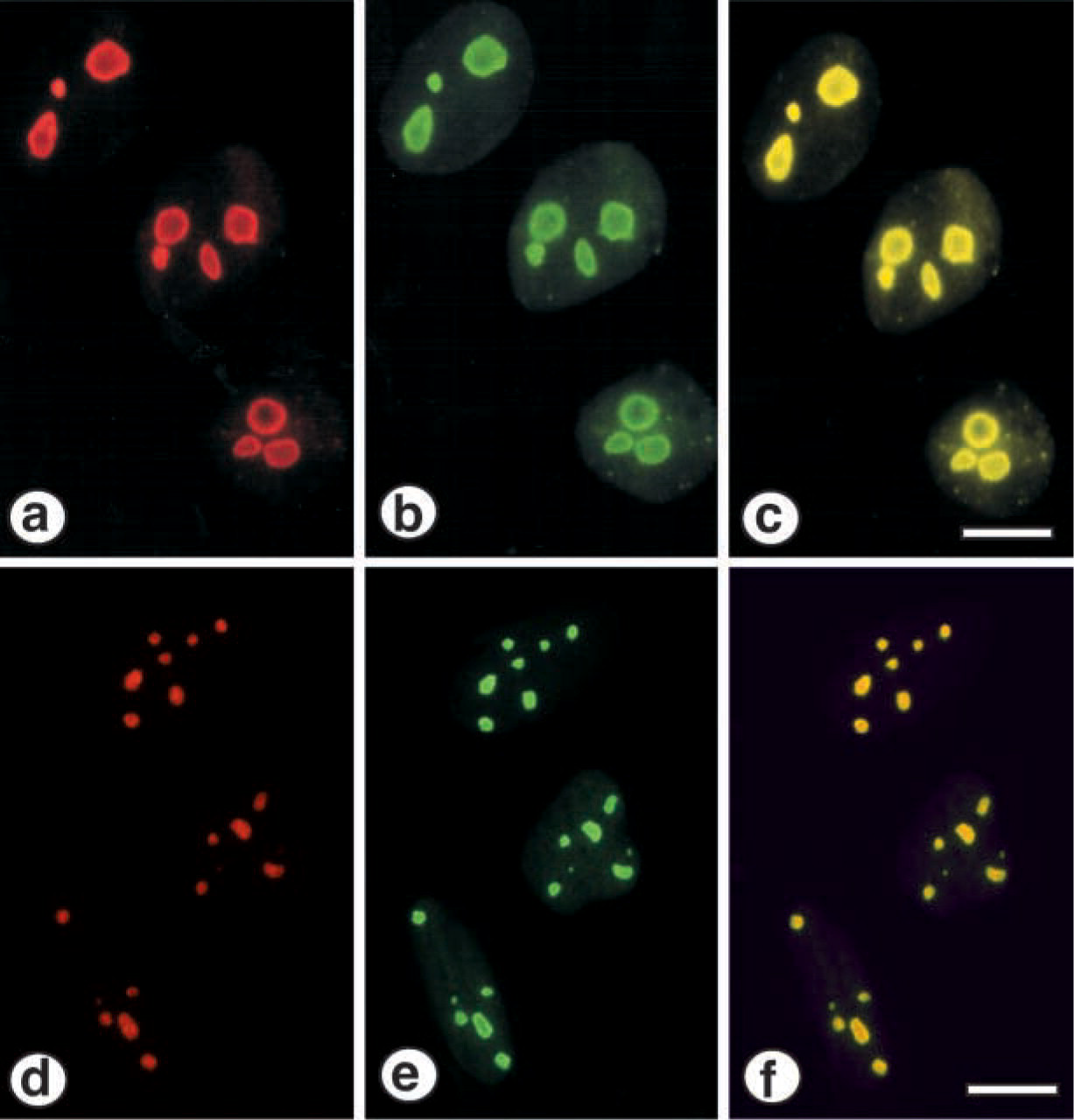
Immunocytochemical identification of PP1δ and nucleolin in MG63 and Saos-2 cells. The staining method is described in the text. (
Discussion
The specific localization of the catalytic subunit of PP1δ in nucleoli in MG63 cells and Saos-2 cells indicates that the nucleolus is the site at which this enzyme functions. These findings confirm our previous results (Haneji et al. 1998) and those of others (Kotani et al. 1998; Murányi et al. 1998). The antibody used in the latter reports was raised against the peptide with an additional three amino acids to the peptide that we had used to raise the antibody. However, other studies reported that PP1γ1 rather than PP1δ was localized within the nucleolus (Andreassen et al. 1998; Trinkle-Mulcahy et al. 2001). The difference of the results obtained from the independent laboratory may be explained in part by the specificity of the antibody used and different conditions used in the independent experiment. The process regulating the nucleolar location of PP1δ is quite interesting. It should be noted that PP1δ does not exhibit any evidence of nuclear localization signals in its sequence (Sasaki et al. 1990; Shima et al. 1993b,c). However, proteins the sizes of the catalytic subunit of PP1δ have been shown to freely diffuse into the nucleus without the necessity for specific translocation sequences. The non-catalytic subunits of PP1s have a targeting function, enabling the phosphatases to associate with a particular cellular fraction. Once in the nuclei, PP1s bind to immobile components to maintain their nuclear localization. The highest concentration of PP1s is found in the nucleus, where the enzymes are both in the nucleoplasm and associated with heterochromatin (Jagiello et al. 1995). PP1 activity increased in the nuclei of hepatocyte primary cultures stimulated by EGF, whereas no isoforms showed any changes in concentration under these conditions (Kakinoki et al. 1994). The nucleolar activity of PP1s, including PP1δ, is not known at present.
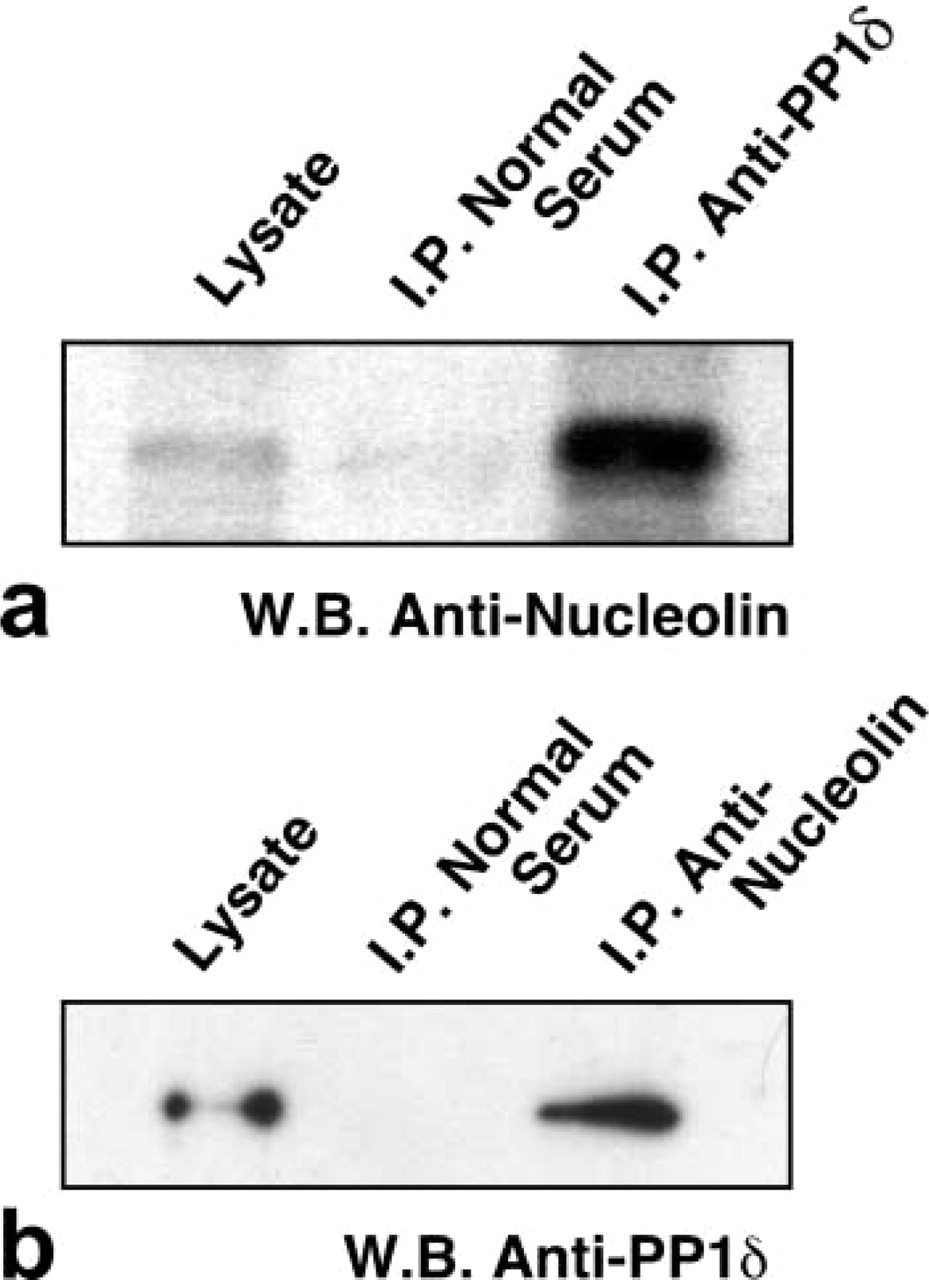
Interaction of PP1δ and nucleolin in MG63 cells. A whole-cell lysate was immunoprecipitated with normal rabbit serum or with anti-PP1δ antibody (
We paid much attention to the relationship between PP1δ and nucleolin in the present study. We demonstrated that PP1δ was localized in the nucleolus in cultured MG63 and Saos-2 cells and that this enzyme was associated with nucleolin. The function of PP1s depends on the localization of these enzymes in cells. Because, like protein kinases, protein phosphatases should be associated with or located near to their substrates (Inagaki et al. 1994), the substrates of PP1δ should be present in the nucleolus. PP1 binding proteins are believed to have an R/KV/IXF, R/KXV/IF (Eg loff et al. 1997) or IKGI (Huang et al. 1999; Shirato et al. 2000) motif; however, such motifs are not identified in nucleolin. The association between PP1δ and nucleolin should be indirect, and some regulatory sub-units might exist that allow PP1δ to bind to nucleolin. However there remains the possibility that nucleolins bind directly to PP1 by unknown binding domains or mechanisms. The interaction between PP1δ and nucleolin demonstrated by immunoprecipitation and Western blotting analysis suggests that PP1δ might be the enzyme that dephosphorylates nucleolin. Although whether a 95-kD protein is a dephosphorylated form of nucleolin is not clear at present, the amount of a 95-kD protein decreased after calyculin A treatment in MG63 cells (unpublished data). These findings suggest that nucleolin could be partially dephosphorylated into a 95-kD protein detected with an anti-nucleolin antibody. Another possibility, that nucleolin is partially proteolyzed into a 95-kD protein, is not ruled out. It was also reported that localization of PP1δ was similar to that of nucleophosmin and that nucleophosmin could be a candidate substrate for PP1δ (Kotani et al. 1998).
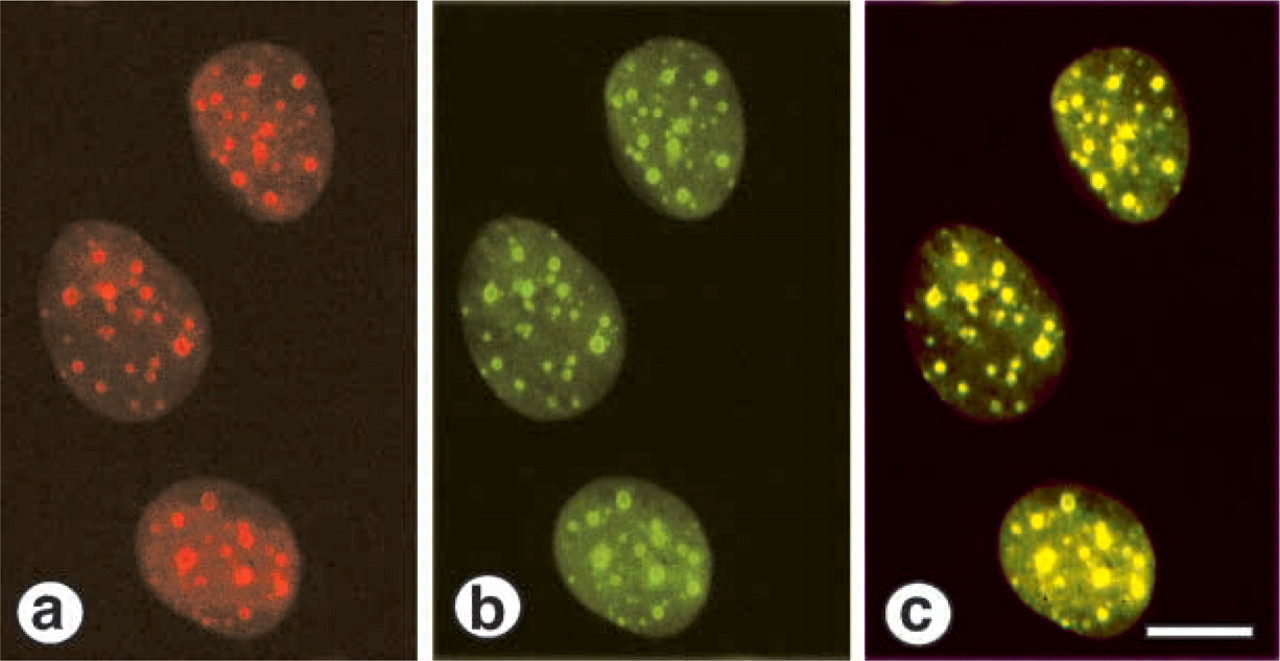
Immunocytochemical identification of PP1δ and nucleolin in actinomycin D-treated MG63 cells. The cells were treated with 10 nM actinomycin D for 4 hr. The cells were stained for PP1δ and nucleolin as described in the text. (
Interestingly, the 95-kD protein was present in the nucleoplasm and cytoplasm in MG63 cells. Once nucleolin is dephosphorylated, it might be translocated from the nucleolus to the nucleoplasm or cytoplasm and remain there by some mechanism yet to be examined. The relocation mechanism might involve the reversible phosphorylation of the nucleolin in the cytoplasm by cell cycle-dependent kinases. It was reported that nucleolin is phosphorylated by several kinases, including casein kinase, p34cdc2 kinase, and protein kinase C (Schneider et al. 1989; Zhou et al. 1997). PP1δ is reported to be associated with Rb protein (Puntoni and Villa-Moruzzi 1997), indicating that PP1δ may be the enzyme that dephosphorylates Rb proteins (Nelson et al. 1997).
Actinomycin D, an anti-tumor antibiotic, is known to inhibit RNA synthesis. When cultured mammalian cells are exposed to relatively low concentrations of actinomycin D, the synthesis of the 45S nucleolar precursor of 18-28s rRNA is inhibited, whereas there is little or no effect on the synthesis of tRNA and 5S rRNA (Perry and Kelley 1970). In the present study PP1δ and nucleolin redistributed from the nucleoli to the nucleoplasm after treatment with 10 nM actinomycin D for 4 hr. This condition is enough to cause inhibition of rRNA rather than mRNA synthesis and causes translocation of nucleophosmin (B23) from the nucleolus to the nucleoplasm in HeLa cells (Yung et al. 1985). At the onset of mitosis, nucleolin localized diffusely in the nucleus, a pattern that was similar to the redistribution pattern of nucleolin in the actinomycin D-treated cells. This pattern is shared with a number of other nucleolar proteins, including nucleophosmin, fibrillarin, and perichromonuclein (Norbury and Nurse 1992; Wu et al 1995; Liu and Yung 1999). Phosphorylation of these proteins is regulated by p34cdc2/cyclin B or casein kinase II (Caizergues-Ferrer et al. 1987; Belenguer et al. 1989). It is likely that alternative localization of these proteins and the increased levels of PP1δ and 95-kD nucleolin are regulated by the action of kinases and phosphatases. Although the mechanisms of PP1δ expression and the role of 95-kD nucleolin are not known, the interaction between PP1δ and nucleolin may be correlated with dephosphorylation of nucleolin in MG63 cells. The idea that the function of nucleolin is coupled with the phosphorylation status is supported by the observation that active rRNA transcription is correlated with the phosphorylation level of nucleolin (Belenguer et al. 1990).
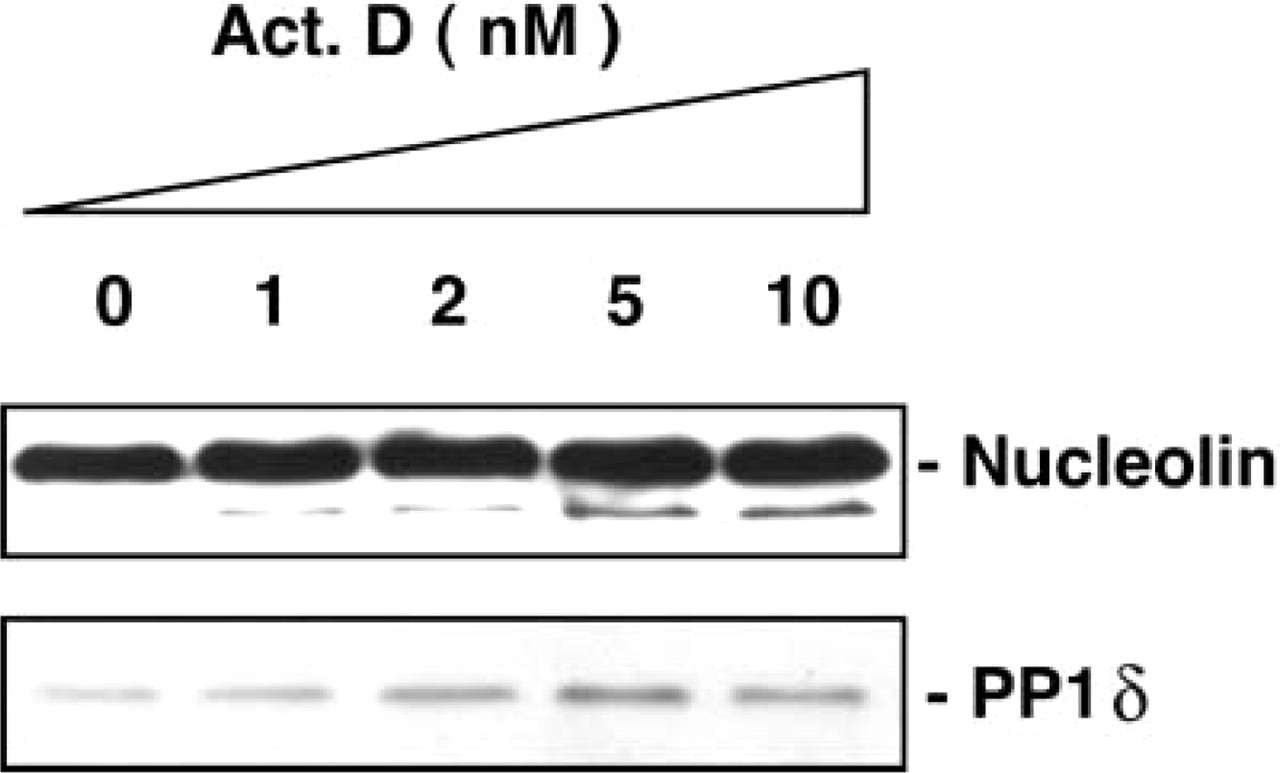
Western blotting analysis of PP1δ and nucleolin in acti-nomycin D-treated MG63 cells. The cells were treated for 4 hr with actinomycin D at the various concentrations indicated.
Footnotes
Acknowledgments
Supported in part by a Research Fellowship of the Japan Society for the Promotion of Science for Young Scientists (HM) and by a grant from the Grant-in-Aid for Scientific Research from the Ministry of Education, Science, Sports, and Culture of Japan (TH).
We thank Ms Eiko Sasaki for skillful technical assistance.