
Abstract
CD133 is an antigen expressed on hematopoietic progenitor cells and on some epithelial cells. We previously reported that a commercially available antibody against CD133, CD133–2/AC141, also reacted with an intracellular protein in placental trophoblasts. Here we show by 2D electrophoresis and mass spectroscopy that this reactivity is with cytokeratin 18, a cytokeratin present in most simple epithelia. Immunohistochemistry (IHC) with CD133–2/AC141 on a trophoblast cell line displayed a staining pattern typical for the cytoskeleton. Cryostat sections of stratified epithelia lacking cytokeratin 18 did not react with CD133–2/AC141. In conclusion, care must be taken not to misinterpret staining patterns using CD133–2/AC141 in IHC.
CD133 is a cell surface glycoprotein localized in membrane protrusions or microvilli. It is expressed by stem and progenitor cells developing into hematopoietic lineages (Yin et al. 1997), endothelial cells (Peichev et al. 2000), neurons and glial cells (Uchida et al. 2000), and by some epithelial cells (Corbeil et al. 2000). Originally called AC133, it was later renamed CD133 or PROML1 (Corbeil et al. 2000). Several monoclonal antibodies (MAbs) against CD133 are commercially available (Miltenyi Biotech; Bergisch Gladbach, Germany), which have become important tools in hematology. While searching for CD133-positive endothelial progenitor cells in cryosections of early placenta, we noted clear reactivity of antibody CD133–2 (former name AC133–2, clone AC141), but not of antibody CD133–1, with trophoblasts, the epithelial layer of placental villi (Pötgens et al. 2001) (Figure 2B). Endometrial gland epithelial cells were also found to have this reactivity. The AC141-reactive protein in trophoblast was found only intracellularly and had a molecular weight much smaller than the 120 kD reported for the CD133 protein. We concluded that this protein could either be a cytoplasmic variant of the CD133 protein, generated by alternative splicing or by proteolytic processing, or some other protein coincidentally crossreacting with AC141.
We tried to identify the protein responsible for this reactivity. Cell extracts were prepared from a choriocarcinoma–trophoblast hybrid cell line, AC-1M32 (Frank et al. 2000; deposited and available at the DSMZ, www.dsmz.de, #ACC442), used as a model for trophoblast. Cells grown to confluence were harvested with a cell scraper and lysed in 7 M urea, 2 M thiourea, 40 mM Tris, 65 mM DTT, 4% CHAPS. The lysates were cleared by centrifugation for 1 hr at 17,000 × g. The proteins were separated in analytical 2D gels (Amersham Pharmacia Biotech; Uppsala, Sweden), with isoelectric focusing (IEF) in the first dimension and SDS-PAGE in the second dimension. After electrophoresis the proteins were either blotted to PVDF membranes (Roche; Mannheim, Germany) or silver stained according to Heukeshoven and Dernick (1988). Blots were immunostained essentially as in Pötgens et al. (2001). They were blocked in TTBS buffer with 2% skim milk powder, then incubated with AC141 (diluted 1:100), rabbit anti-mouse (DAKO, Hamburg, Germany; diluted 1:200), peroxidase/mouse anti-peroxidase (PAP, DAKO; diluted 1:400), and Lumilightplus substrate (Roche). X-ray films were exposed for 1–10 min and developed. On 2D blots covering a pI range of 3–10, a series of positive spots was found with a pI of 5.5–5.9 and an apparent molecular weight of 48 kD (majority) to 54 kD. For a better separation, subsequent 2D blots were prepared covering only the pI range between 4 and 7. The spots reacting with AC141 were localized on silver-stained 2D gels run in parallel (Figure 1). One spot that was well separable from neighboring spots was excised from a zink-stained (BioRad; Hercules, CA) 2D gel and sent for a commercial peptide mass fingerprinting analysis (Swiss-2Dservice, Geneva, Switzerland; http://www.expasy.ch/ch2d/service/). Its pattern turned out to be typical for cytokeratin 18 (see Table 1).
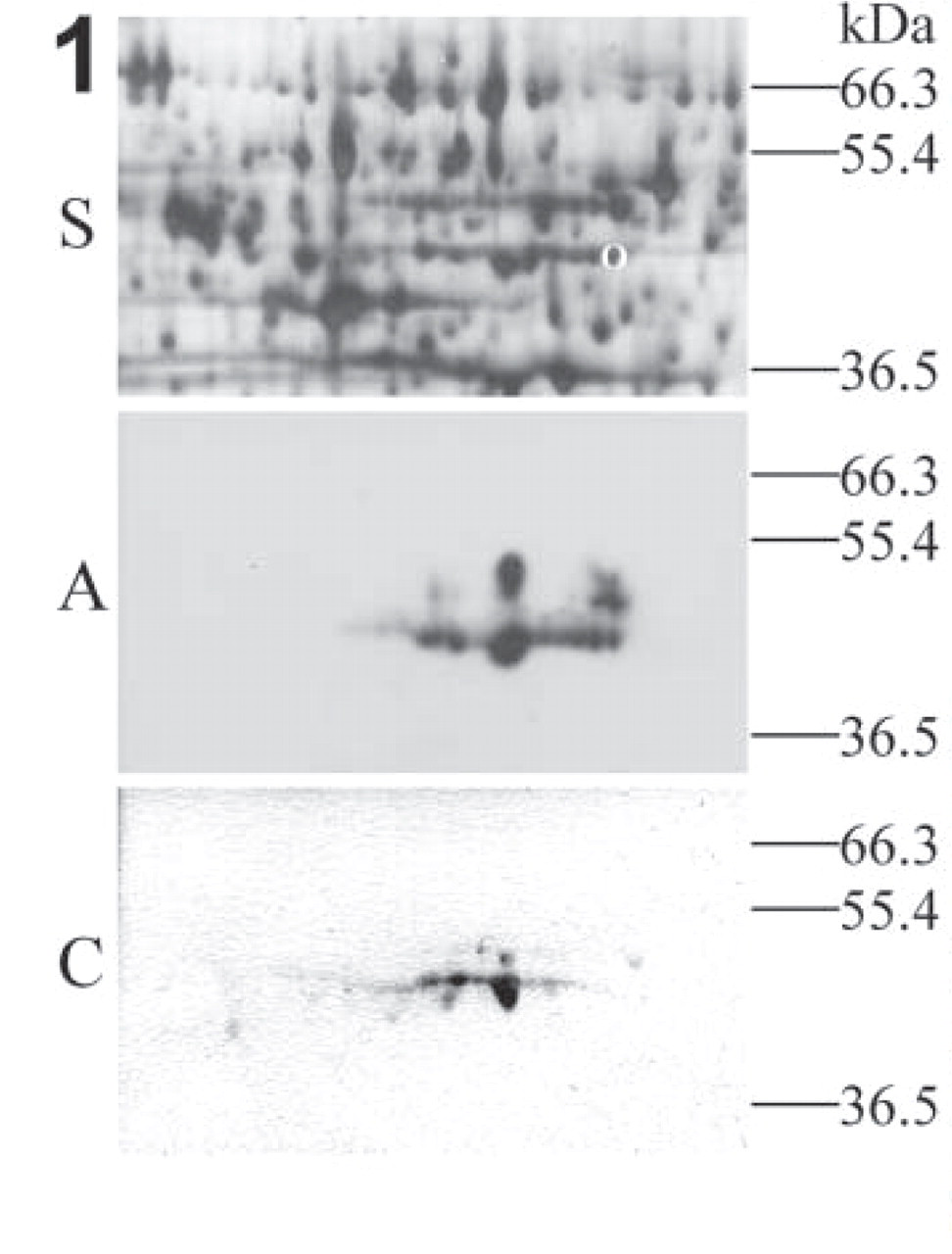
2D PAGE and immunochemistry on 2D blots. Cellular extracts from cell line AC-1M32 were separated on 2D gels and either silver-stained (S) or blotted and incubated with AC141 (A) or anti-cytokeratin 18 antibody (C). Only relevant parts are shown. The pI range shown here is 5.0–6.1. The circle in the silver-stained gel depicts the spot excised for peptide mass fingerprinting analysis. The AC141 staining was made visible by chemiluminiscence, whereas the anti-CK18 reaction was developed with AEC substrate.
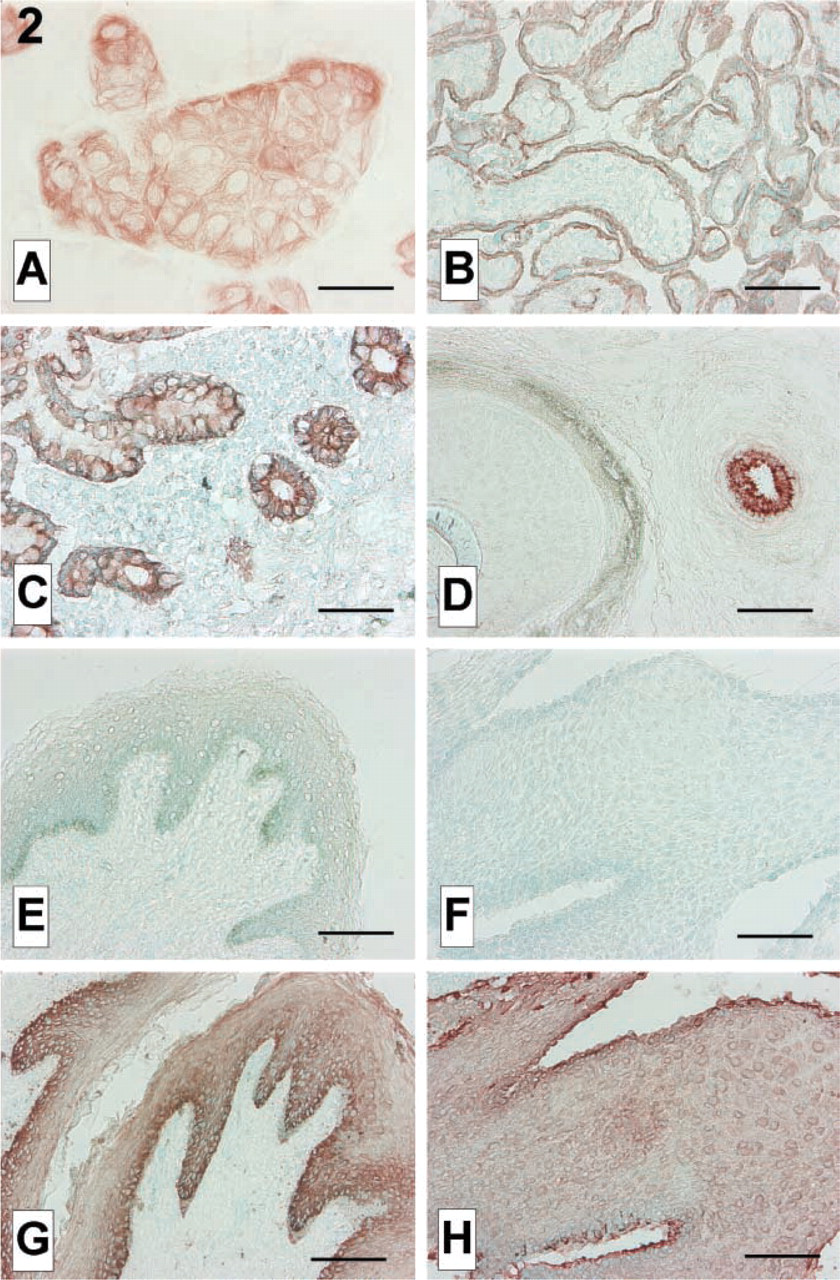
IHC on cultured cells and cryostat sections. (
Cytokeratin 18 is a member of the family of acidic cytokeratins (cytokeratins 9–20) that join with basic cytokeratins (1–8) to produce the intermediate filaments of epithelial cells. Cytokeratin 18 is expressed mainly in simple epithelia, combines with cytokeratin 8 (Moll et al. 1982; Cooper et al. 1985), and is one of the cytokeratins present in trophoblasts, along with cytokeratins 7, 8, and 19 (Mühlhauser et al. 1995; Haigh et al. 1999).
IHC confirmed reactivity of AC141 with a cytoskeletal protein in trophoblast cell line AC-1M32. Cells cultured on eight-chamber glass slides were fixed for 10 min in acetone at −20C, dried, rehydrated in PBS, pretreated in methanol/hydrogen peroxide, and immunostained with AC141 essentially as in Pötgens et al. (2001). In short, cells were blocked in human serum and rabbit serum and incubated with AC141 (diluted 1:20), rabbit anti mouse-biotin, streptavidin–peroxidase, and AEC substrate (Zymed; San Francisco, CA), respectively. In both cell lines a cytoplasmic network reminiscent of cytoskeleton was stained (Figure 2A).
Peaks found after peptide mass fingerprinting analysis of the protein excised from 2D gel corresponding with peaks from a cytokeratin 18 spot in the Swiss-2Dservice database a
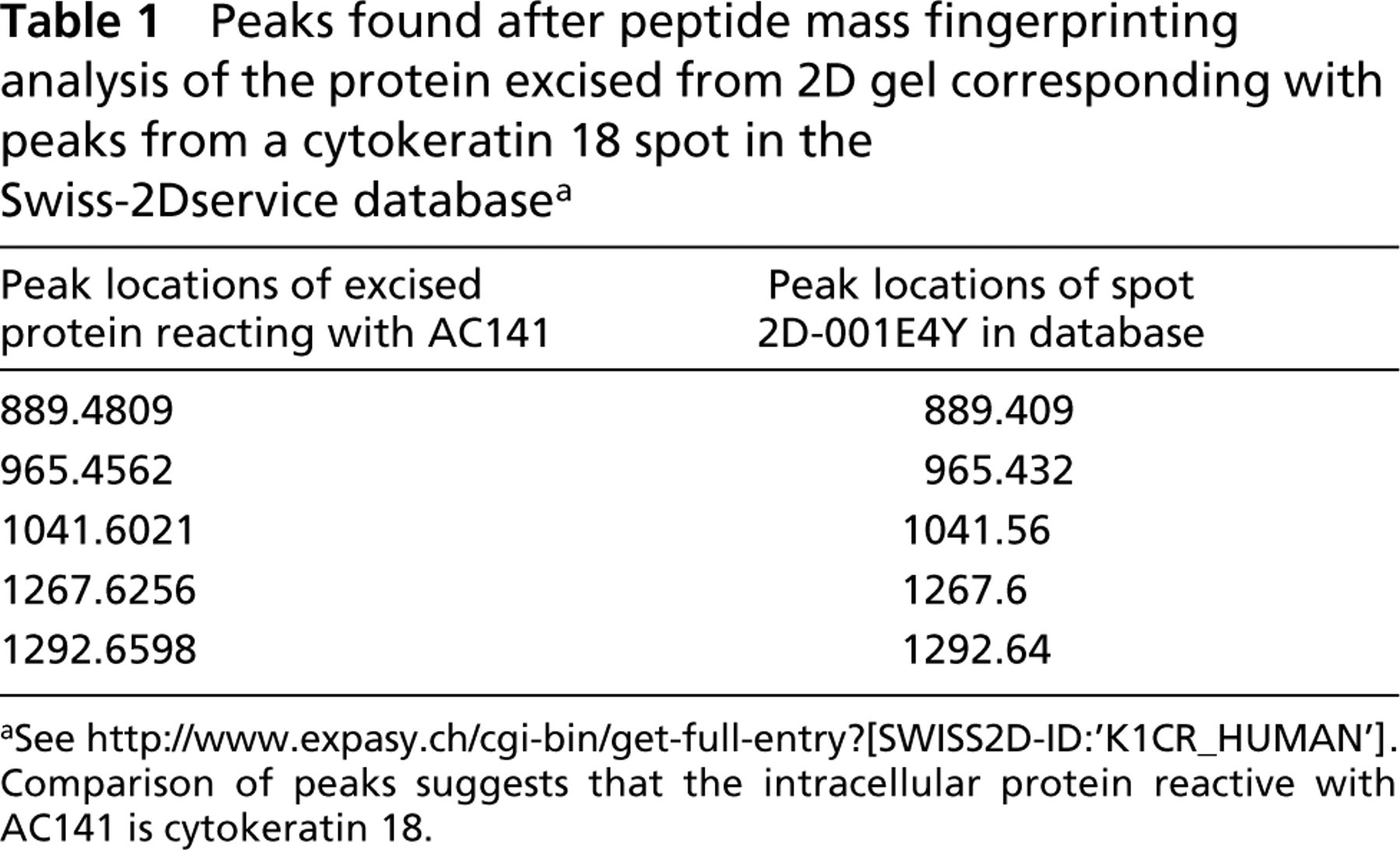
aSee http://www.expasy.ch/cgi-bin/get-full-entry?[SWISS2D-ID:‘K1CR_HUMAN’]. Comparison of peaks suggests that the intracellular protein reactive with AC141 is cytokeratin 18.
To further confirm that cytokeratin 18 is the AC141-reactive protein in trophoblast, we incubated 2D blots with an MAb against cytokeratin 18 (clone Cy90, Sigma, Taufkirchen, Germany; diluted 1:10,000), rabbit anti-mouse, and PAP, as above, and developed the blot in AEC substrate for 5 min (Figure 1). Anti-cytokeratin 18 incubation revealed a row of spots of 48 kD having a pI of 5.5–5.9, as well as some weaker spots of about 52 kD. The staining pattern after incubation with anti-cytokeratin 18 was very similar to that seen after AC141 staining. According to the literature (Moll et al. 1982; Cooper et al. 1985), cytokeratin 18 is 45 kD and has an average pI of 5.7. Multiple spots with slightly different pI occur due to the presence of differential phosphorylation. The small difference in molecular weight with the main spots determined by us (48 kD) may be due to the use of other markers and to slight differences in the performance of electrophoresis.
Crossreactivity of AC141 with other cytokeratins than cytokeratin 18 cannot be excluded. Trophoblasts and other simple epithelia express only a subset of all cytokeratins, including cytokeratin 18. Other sets of cytokeratins are found in stratified epithelia, such as cytokeratins 1, 2, and 10 (typical for keratinized skin-type epithelia) or 4 and 13 (typical for unkeratinized stratified squamous epithelia) (Moll et al. 1982; Cooper et al. 1985), whereas cytokeratin 18 is absent in most stratified epithelia.
To study reactivity of AC141 with various types of epithelia, we immunostained acetone-fixed cryostat sections of placenta (obtained after spontaneous birth), ilium, gingiva, axillar skin (all three removed for diagnostic purposes during surgery), and foreskin of penis (removed by circumcision) with AC141 (diluted 1:50; method as above). Placental trophoblast, mucosal epithelium of ilium, and eccrine sweat glands, all of which are simple epithelia, reacted with AC141 (Figures 2B–2D). Keratinized and unkeratinized stratified epithelia, such as those in hair follicle, epidermis, and gingiva, did not show reactivity with AC141 (Figures 2D–2F). The stratified epithelia did react, however with a pan-cytokeratin antibody (MNF116, DAKO; diluted 1:300), as shown in Figures 2G and 2H. AC141 therefore reacted with various epithelia expressing cytokeratin 18 but not with epithelia lacking cytokeratin 18.
In conclusion, an MAb, AC141, raised against membrane antigen CD133, displays additional reactivity with cytokeratin 18, a cytoskeletal protein present in many epithelia. Although some epithelial cells actually do express CD133 (Corbeil et al. 2000), the data here show that not all epithelial reactivity with AC141 points to the presence of CD133. In flow cytometry, this crossreactivity of AC141 with trophoblasts was only seen if cells to be analyzed had damaged cell membranes or had been fixed with 70% methanol such that cell membranes had become leaky (Pötgens et al. 2001). In hematological applications, there is a theoretical chance of dead circulating epithelial cells (e.g., from epithelial tumors, or circulating trophoblast fragments in pregnant women) reacting with AC141, yielding false-positive events if hematopoietic progenitors are intended to be marked. However, numbers of circulating tumor or trophoblast cells are expected to be extremely low, because these cells are rapidly removed from the circulation. For a few circulating trophoblast cells to be detected at all, a prior enrichment step is necessary (Johansen et al. 1999).
AC141 can be used as an alternative antibody against cytokeratin 18 in IHC. It should be noted, however, that it displays this reactivity only in cryostat sections, not in paraffin sections (Pötgens et al. 2001; and data not shown). When AC141 is used to find CD133-expressing cells in cryostat sections, one could easily be misled. Antibody CD133–1, which does not show this crossreactivity, or other antibodies against CD133 would be a better choice in IHC applications.
Footnotes
Acknowledgements
Supported by funds from the University of Technology Aachen (START 78/2000 to AP), the Deutsche Forschungs-gemeinschaft (DFG, grants no. FR1245/3–2,3 to HGF and PK), and the Rockefeller Foundation (grants no. RF 96020#76, RF98024#99, RF99021#114 to PK and HGF).
We thank Angela Rüben and Anke Gärtner for excellent technical assistance and Drs Frank Jugert, Thorsten Laurin, Dennis von Heimburg, and Sören Gärtner (University Hospital Aachen) for their generous gifts of tissue specimens.