
Abstract
The expression and distribution of the long form of Type XII collagen were investigated histochemically during chicken corneal development using a monoclonal antibody (P3D11) raised against the N-terminal domain of chicken Type XII collagen. Specificity of the antibody was confirmed by immunoprecipitation before and after bacterial collagenase digestion. Immunofluorescent microscopic studies showed that during chicken cornea formation, the long form of Type XII collagen is initially detected on Day 3 embryo (stage 19) in the sub-epithelial matrix of the corneal periphery and in the matrix around the optic cup. On Day 5 embryo (stage 27) the long form was expressed in the primary stroma. Thereafter, as the secondary stroma was formed, the long form localized in the sub-epithelial and sub-endothelial matrices and in the anterior region of the limbus (corneoscleral junction) before the formation of Descemet's and Bowman's membranes. After hatching, the immunoreactivity decreased predominantly in the sub-epithelial and sub-endothelial matrices but remained at the anterior region of the limbus. Immunoelectron microscopic examination demonstrated that the long form localizes in the Descemet's and Bowman's membranes and along the collagen fibrils in the stroma with a periodic repeat. Based on the distribution of the long form of Type XII collagen in the sub-epithelial and sub-endothelial matrices and limbus, it was suggested that the long form of Type XII collagen is involved in formation of the Descemet's and Bowman's membranes and in stabilization of the limbus.
Keywords
T
The cornea consists of five layers: corneal epithelium, Bowman's membrane, corneal stroma, Descemet's membrane, and corneal endothelium. The corneal epithelium consists of non-keratinized stratified squamous cells and overlies Bowman's membrane. The Bowman's membrane consists of homogenous lamina and is composed of randomly oriented collagen fibrils (18–20 nm in diameter). The corneal stroma consists of parallel bundles of collagen fibrils (23–25 nm in diameter). Descemet's membrane is a basement membrane underlying the corneal endothelium. The corneal endothelium is a single layer of flattened cells that face towards the anterior chamber in the eyeball. In adult human cornea, the long form is present in the corneal stroma and Bowman's membrane but is absent in the limbus (Wessel et al. 1997). In rabbit cornea, the long form appears initially on Day 12 after birth just before the eyelid opens and localizes thereafter in the corneal basement membrane, but is absent in the limbus (Anderson et al. 2000). In contrast, in chicken cornea of Day 5–18 embryos, Type XII collagen localizes in the matrix interfaces between the corneal epithelium and stroma, between the corneal endothelium and stroma, and in the limbus (Gordon et al. 1996). These reports suggest that Type XII collagen is involved in attachment of the corneal epithelium to the underlying tissue, in promotion of stromal condensation, and in stabilization of the matrix interfaces (Gordon et al. 1996; Linsenmayer et al. 1998). However, it has not yet been elucidated what biological roles the long and short forms have in chicken cornea formation, respectively. Because it has already been reported that human corneal stroma contains the long form as the predominant form of Type XII collagen (Wessel et al. 1997), we raised a monoclonal antibody against the N-terminal region of the long form and used it to examined its fine localization during chicken corneal development.
Materials and Methods
Materials
Tissue culture medium, fetal calf serum, chicken serum, and other tissue culture supplies were obtained from Gibco BRL (Grand Island, NY). Highly purified collagenase from Clostridium histolyticum was obtained from Worthington Biochemicals (Lakewood, NJ). Protein A-Sepharose 4FF, [35S]-methionine (1000 Ci/mM), and Ni-chelate affinity absorbent (Ni-NTA-agarose) were purchased from Amersham Life Science (Arlington Heights, IL) and Qiagen (Hilden, Germany), respectively.
Tissues
All experimental procedures using laboratory animals were approved by the Animal Care and Use Committee of Kyorin University School of Medicine. Fertilized eggs of White Leghorn chickens were incubated at 38C until desired stages. The embryonic stages were classified according to the nomenclature of Hamburger and Hamilton (1951). The eyeballs of Day 1.5–20 chicken embryos and of 4-day-old and adult chickens were used in the present study.
Expression of Recombinant Chicken Type XII Collagen
The cDNA encoding the P3 fragment of the N-terminal region in chicken Type XII collagen (Yamagata et al. 1991) was constructed into the pQE-9 expression vector (Qiagen) to obtain the histidine-tagged recombinant protein in bacteria. The expressed protein was purified by Ni-NTA-agarose column chromatography according to the manufacturer's instructions.
Metabolic Labeling of Fibroblasts with [35S]-Methionine
Chicken embryonic fibroblasts (CEFs) prepared by the method described previously (Hungerford et al. 1996) were cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 5% fetal bovine serum (FBS), 1% chicken serum, 50 U/ml penicillin, 50 mg/ml streptomycin, 1 mM glutamine, and 1 mM sodium pyruvate, which is referred to as a growth medium in the present study. For metabolic labeling of proteins, CEFs at a subconfluent stage were incubated for 16 hr with 200 mCi [35S]-Met in a mixture of growth medium and Met-free growth medium (1:9, v/v).
Generation of Monoclonal Antibody
Approximately 200 mg of recombinant chicken Type XII collagen dissolved in PBS was mixed with complete Freund's adjuvant and injected IP into BALB/c mice. Two booster injections with the same amount of the antigen were given at 2-week intervals. On the third day after the final booster injection, the spleens were excised and isolated splenocytes were fused with SP2/0 myeloma cells using 50% polyethylene glycol 1500 (Boehringer-Manheim; Manheim, Germany) according to the manufacturer's instructions. Hybridomas growing in HAT selection medium were initially screened by immunohistochemical study using chicken tendon tissue sections and then by immunoprecipitation using the [35S]-Met-labeled proteins. The monoclonal antibody obtained in the present study was designated as P3D11.
Immunoprecipitation of Type XII Collagen
CEFs metabolically labeled with [35S]-Met were washed four times with serum-free DMEM and lysed with 1 ml of PBS containing 2% Triton X-100, 1 mM EDTA, and 3 mM phenylmethylsulfonylfluoride (PMSF) (lysis buffer) on ice for 10 min. The following procedures were performed at 4C unless otherwise specified. The cell lysates were centrifuged at 10,000 × g for 30 min. The resultant supernatant was incubated with the antibody (2 mg/ml) for 15 min, and the immune complexes were mixed with protein A-Sepharose beads (20 μl gel for 1 ml of cell lysates). The immunocomplexes bound to the beads were washed several times with the lysis buffer and with Dulbecco's PBS, and then eluted from the beads with 100 mM acetic acid or by boiling in 1% SDS. In some experiments the immunoprecipitated Type XII collagen was digested with bacterial collagenase (0.1 U/ml in DPBS) at 37C for 60 min. The reaction was terminated by adding SDS-PAGE sample buffer containing 1 mM dithiothreitol (DTT), boiling for 90 sec, and then subjection to SDS-PAGE.
Immunofluorescent Light Microscopic Observation
Whole chicken embryos or eyeballs were embedded in OCT compound (Miles; Elkhart, IL) and frozen with liquid nitrogen. Sections (10 μm thick) were made with a cryostat (Sakura Seiki; Tokyo, Japan), placed on egg albumin-coated slides, and fixed with 2% formaldehyde-PBS at 4C for 30 min. After washing with PBS, nonspecific binding sites were blocked by incubation with 5% donkey serum for 30 min. Sections were then incubated with P3D11 antibody or with non-immunized mouse IgG for 1 hr at room temperature, washed with PBS, and subsequently incubated for 1 hr with Cy3-conjugated donkey anti-mouse IgG antibody (Jackson Immunoresearch; West Grove, PA). Some sections were further incubated with fluorescein-labeled phalloidin (Molecular Probe; Eugene, OR) to stain F-actin. Fluorescein-phalloidin strongly stained the ciliary muscle and also delineated the cell border, which aided identification of immunopositive sites. Nuclei were stained with 4,6-diamidine-2-phenylindole hydrochloride (DAPI) (Boehringer-Mannheim). After washing with PBS, the specimens were mounted in 90% glycerol-0.1 M Tris-HCl buffer (pH 8.5) containing 0.5 mM p-phenylene diamine, which prevents fading of fluorescence during microscopic examination, and observed under an epifluorescent microscope (Nikon; Tokyo, Japan) equipped with chilled color CCD camera Type C5985 (Hamamatsu Photonics; Hamamatsu, Japan).
Immunoelectron Microscopic Study
The HRP labeling study was carried out as described previously (Akimoto et al. 1992). The specimens were fixed with 2% formaldehyde at 4C for 30 min. After washing with PBS they were embedded in 10% gelatin. Non-frozen sections (40 μm thick) were cut with a microslicer DTK-1500 (DSK; Kyoto, Japan). The sections were washed with PBS and incubated with 5% bovine serum albumin-PBS for 1 hr and were then treated with P3D11 antibody at 4C for 48 hr. After washing with PBS, they were incubated with horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG antibody (Dako; Kyoto, Japan) at 4C for 24 hr. As cytochemical controls, non-immunized mouse IgG was incubated with samples. After washing with PBS, the specimens were immersed in 3,3′-diaminobenzidine (DAB) (0.5 mg/ml) solution containing 0.05% H2O2 for 5 min at room temperature and then washed with distilled water. Labeled specimens were postfixed with 1% OsO4 in 0.1 M phosphate buffer (pH 7.3) for 30 min, dehydrated through a graded series of ethanol solutions, exposed to propylene oxide, and embedded in Epon 812. Ultrathin sections were prepared, stained with lead citrate, and observed under a transmission electron microscope (JEM-1010) (JEOL; Tokyo, Japan).
For the colloidal gold labeling study, the HRP-conjugated secondary antibody was replaced by nanogold (Nanoprobes; Stony Brook, NY). The specimens were washed with PBS, then fixed in 2% glutaraldehyde. After rinsing with deionized water, silver enhancement of nanogold was carried out with HQ silver (Nanoprobes) according to the manufacturer's protocol. Development time was 3 min. The specimens were rinsed with deionized water, then fixed with 0.1% OsO4 for 30 min, dehydrated through a graded series of ethanol solutions, exposed to propylene oxide, and embedded in Epon 812. Ultrathin sections were stained with lead citrate and uranyl acetate and observed under a transmission electron microscope (JEM-1010). In some experiments, samples were fixed in 2.5% glutaraldehyde and processed for conventional EM observation as described previously (Akimoto et al. 1991).
Results
Specificity of Monoclonal Antibody Against Chicken Tendon Type XII Collagen
Occurrence of the long and short forms in Type XII collagen was reported in several animal species (Kania et al. 1999; Kato et al. 2000). To elucidate the functions of the long form, which contains the domains interacting with cells and extracellular matrix components, a monoclonal antibody was raised in mouse against the recombinant P3 fragment located at the N-terminal region of the chicken NC3 domain (Yamagata et al. 1991) (Figure 1, P3). The antibody, P3D11, was incubated with [35S]-Met-labeled chicken embryonic fibroblast lysates and the immune complexes precipitated were analyzed by SDS-PAGE under a reducing condition. Two sets of the doublet bands with approximate molecular weights of 300 K and 600 K were obtained (Figure 2, Lane A). The 300-K bands contained two structural variants, L1 and L2, of the long form shown in Figure 1, and the 600-K bands, together with a very weak band migrated slightly higher, as shown in Figure 2, Lane A appeared to be dimers (L1L1, L1L2, and L2L2) of the 300-K bands. When immune complexes were digested with bacterial collagenase followed by SDS-PAGE, all bands were shifted to one single band with an approximate molecular weight of 270 K (Figure 2, Lane B). These results indicated that P3D11 antibody specifically recognizes and can immunoprecipitate the long form of Type XII collagen. The short form with an expected molecular weight of 220 K could be expressed in chicken tendon fibroblasts but was not detected with this antibody.
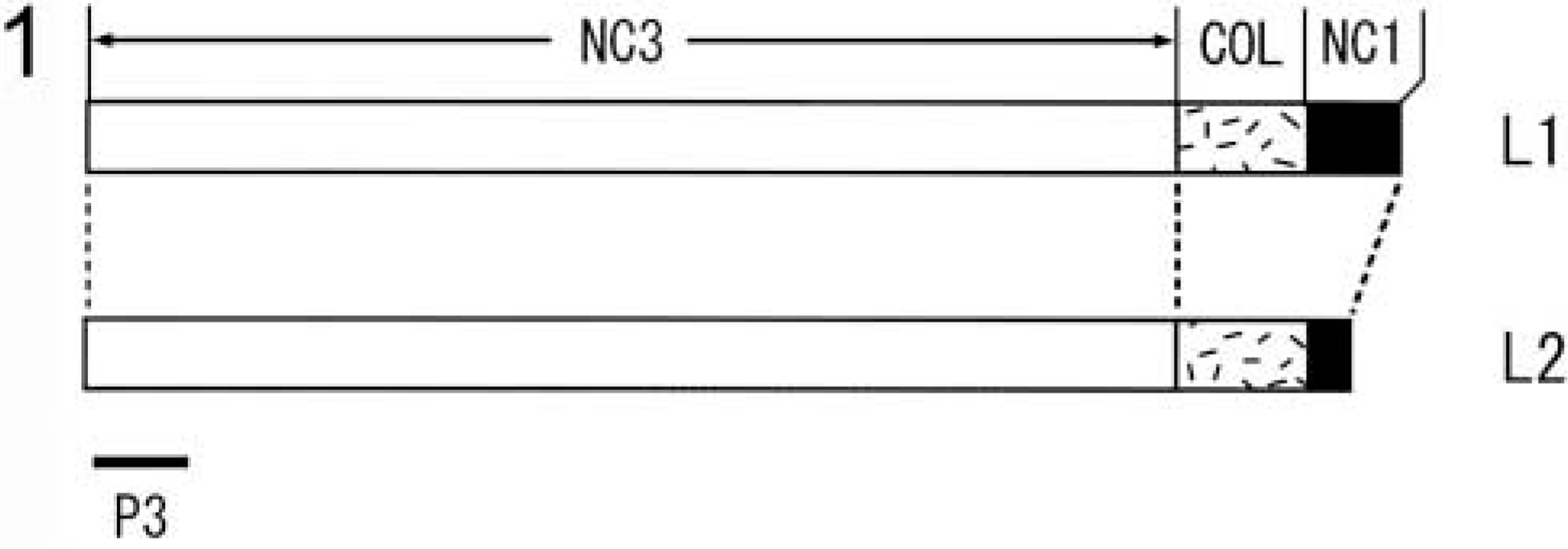
Schematic illustration of the long form of chick Type XII collagen. The long form contains two structural variants, L1 and L2, both of which consist of three non-collagenous domains (NC1, NC2, and NC3) and a collagenous domain (COL). The COL domain is interrupted by the NC2 domain, which is not shown. A solid bar indicates the P3 fragment (Yamagata et al. 1991) whose recombinant peptide was used for raising a monoclonal antibody (P3D11).
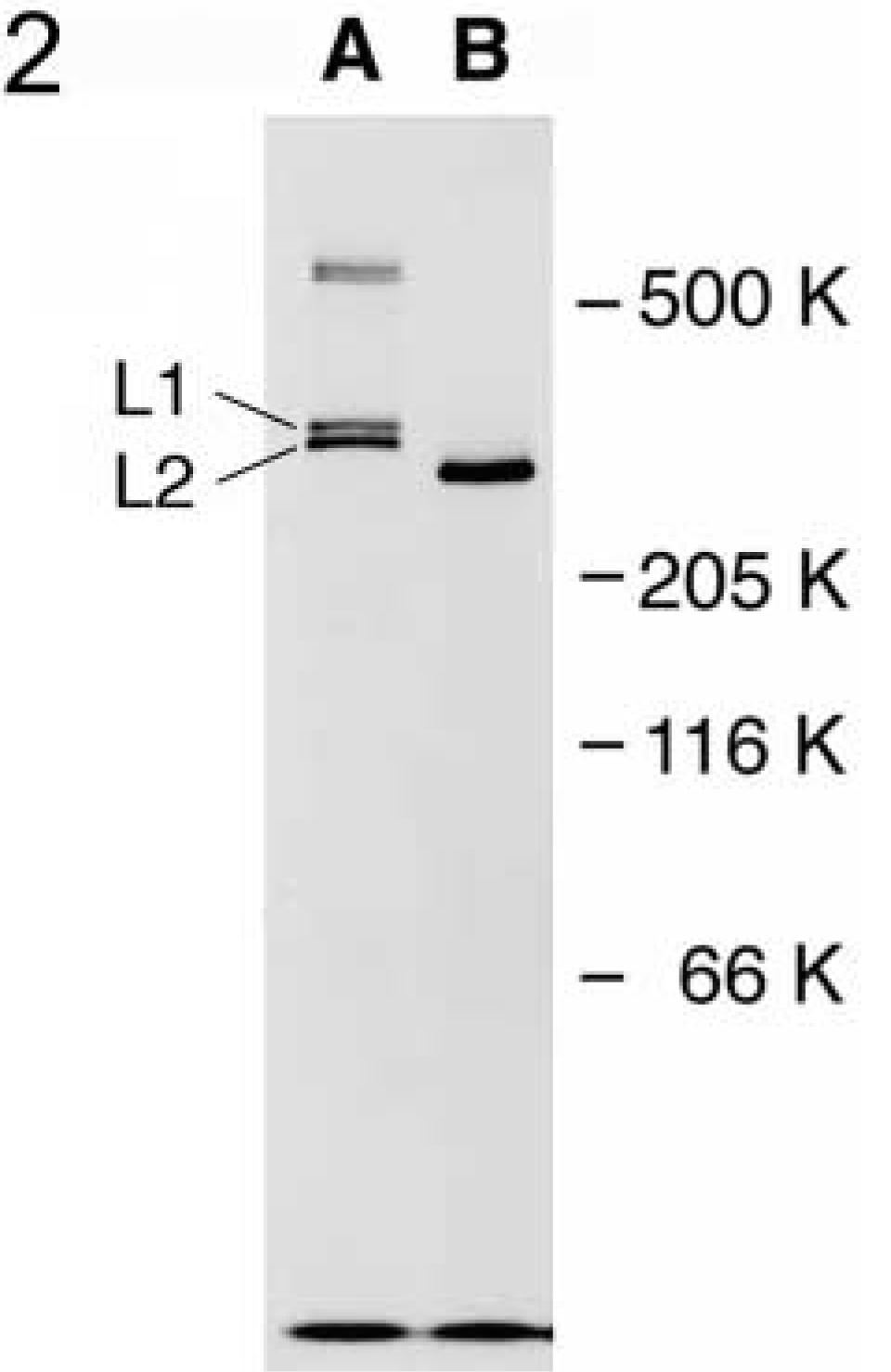
SDS-PAGE analysis of the long form of chick Type XII collagen. Type XII collagen was immunoprecipitated from chicken tendon fibroblasts labeled metabolically with [35S]-Met and subjected to SDS-PAGE under a reducing condition before (Lane A) and after (Lane B) digestion with bacterial collagenase. L1 and L2 indicate the monomers and those at 600 K are crosslinked dimeric molecules. The molecular sizes were estimated based on the positions of human fibronectin (500 K under a non-reducing condition), rabbit myosin (205 K), E. coli β-galactosidase (116 K), and bovine serum albumin (66 K) as standards.
Immunohistochemical Observation of the Long Form of Type XII Collagen in Chicken Cornea by Light Microscopy
To elucidate when and where the long form of Type XII collagen is initially expressed and whether or not its expression changes in the cornea during chicken development, the localization of Type XII collagen in chicken cornea was examined immunohistochemically using the monoclonal antibody described above. The optic vesicles are formed on Day 1.5 of incubation (stage 10), and no immunoreactivity to Type XII collagen was detected in the optic vesicles and the lens placode (Figure 3A). After this stage the lens placode begins to invaginate, and on Day 3 (stage 19) the lens vesicle pinches off and the overlying ectoderm is induced to become corneal epithelium. At this stage the immunoreactivity was detected in the sub-epithelial matrix located peripherally in the cornea, and around the optic lip (Figures 3B–3D). In particular, intense immunoreactivity was present around the outer surface of the optic cup at the proximal side (Figure 3D). By Day 4, the corneal epithelium secretes and forms the primary stroma, and the mesenchymal cells, which are destined to become the corneal endothelium, begin to invade the area. On Day 4 (stage 24), strong immunoreactivity was detected in the mesenchyme between the corneal epithelium and optic lip (Figures 3E and 3F). Weak immunoreactivity was also present between the corneal epithelium and the lens (Figures 3E and 3F). Endothelial cell migration is almost completed by Day 4.5–5 of incubation (stage 25–26). On Day 5 of incubation (stage 27), the immunoreactivity was distributed throughout the primary stroma and in the mesenchyme (Figure 4A). After this stage, the immunoreactivity was localized in the sub-epithelial and sub-endothelial regions (Figures 4B–4D). However, the immunoreactivity disappeared in the sub-epithelial region by Day 10, but not completely (Figure 4B). In contrast, after Day 10 of incubation, the immunoreactivity increased in the sub-epithelial region. On Day 14 of incubation the cornea begins to condense, becomes transparent, ceases its rapid growth, and becomes resistant to deformation (Hay 1980). On Days 15 and 20 of incubation (stages 41 and 45), strong immunoreactivity was observed both in sub-epithelial and sub-endothelial matrices (Figures 4C–4E). During the period of corneal formation, strong immunoreactivity was also detected in the anterior region of the limbus, while in the posteior region of the limbus strong immunoreactivity was observed at Day 10 but decreased thereafter and was scarcely observed at Days 15, 20, and 4 days after hatching (Figures 4B–4D and 4F). Corneal maturation is completed by Day 19–20 of incubation, and soon after the chick hatches at Day 20–21 of incubation. Four days after hatching, the immunoreactivity decreased in both sub-epithelial and sub-endothelial matrices (Figures 4F and 4H) but remained under the corneal epithelium in the limbus (Figure 4G). A similar localization of Type XII collagen in the cornea at 4 days after hatching was observed in adult chicken (data not shown). Control experiments were conducted throughout the study using non-immune IgG and no immunoreactivity was detected (see Figure 4D, inset).
Immunohistochemical Observation of the Long Form of Type XII Collagen in Chicken Cornea by Electron Microscopy
To examine the fine localization of the long form of Type XII collagen in chicken corneal matrix, immuno-electron microscopic analysis was performed by an indirect method using the HRP-conjugated antibody. The HRP method exhibited similar distribution patterns of the long form of Type XII collagen to those revealed by immunofluorescence study as described above. The corneal epithelium laid down the primary corneal stroma on Day 3 and was located close to the optic cup (Figure 5A), and the basement membrane underlay the corneal epithelium and optic cup, in which collagen fibrils were sparsely located near the basement membrane in the stroma (Figure 5B). Immunoreactivity was detected in the basement membrane and along the collagen fibrils attached to the basement membrane, with a periodic repeat of 55–60 nm in Day 3 chicken embryonic cornea (Figures 5C and 5D). Descemet's membrane first appears subjacent to the corneal endothelium in Day 9 chick embryo, and Bowman's membrane appears subjacent to the corneal epithelium in Day 12 chick embryo (Hay 1980; Fitch et al. 1994). Collagen fibrils were enriched in the corneal stroma in Day 10 chicken embryos (Figures 6A and 6B). Strong immunoreactivity was detected along the Descemet's membrane (Figure 6C) and the Bowman's membrane (data not shown). Positive immunoreactivity was also detected in the endothelial cells (Figure 6C), epithelial cells (data not shown), and stromal fibroblasts (Figure 6C). These results indicated that the endothelial cells, epithelial cells, and stromal fibroblasts might produce the long form of Type XII collagen. In addition, immunoreactivity was also detected along the collagen fibrils near Descemet's membrane, approximately 6 μm from the endothelial cell, but was not detected in the deep stroma (Figures 6C and 6D). Collagen fibrils in regions of positive immunoreactivity were more densely arranged than those in the deeper stroma. The periodic repeat along the collagen fibrils observed by the HRP method was also confirmed by the colloidal gold method (Figures 7A and 7B). To check the specificity of the immunostaining, we replaced the anti-Type XII collagen antibody with non-immunized mouse IgG and found no positive staining (Figures 5D and 6D).
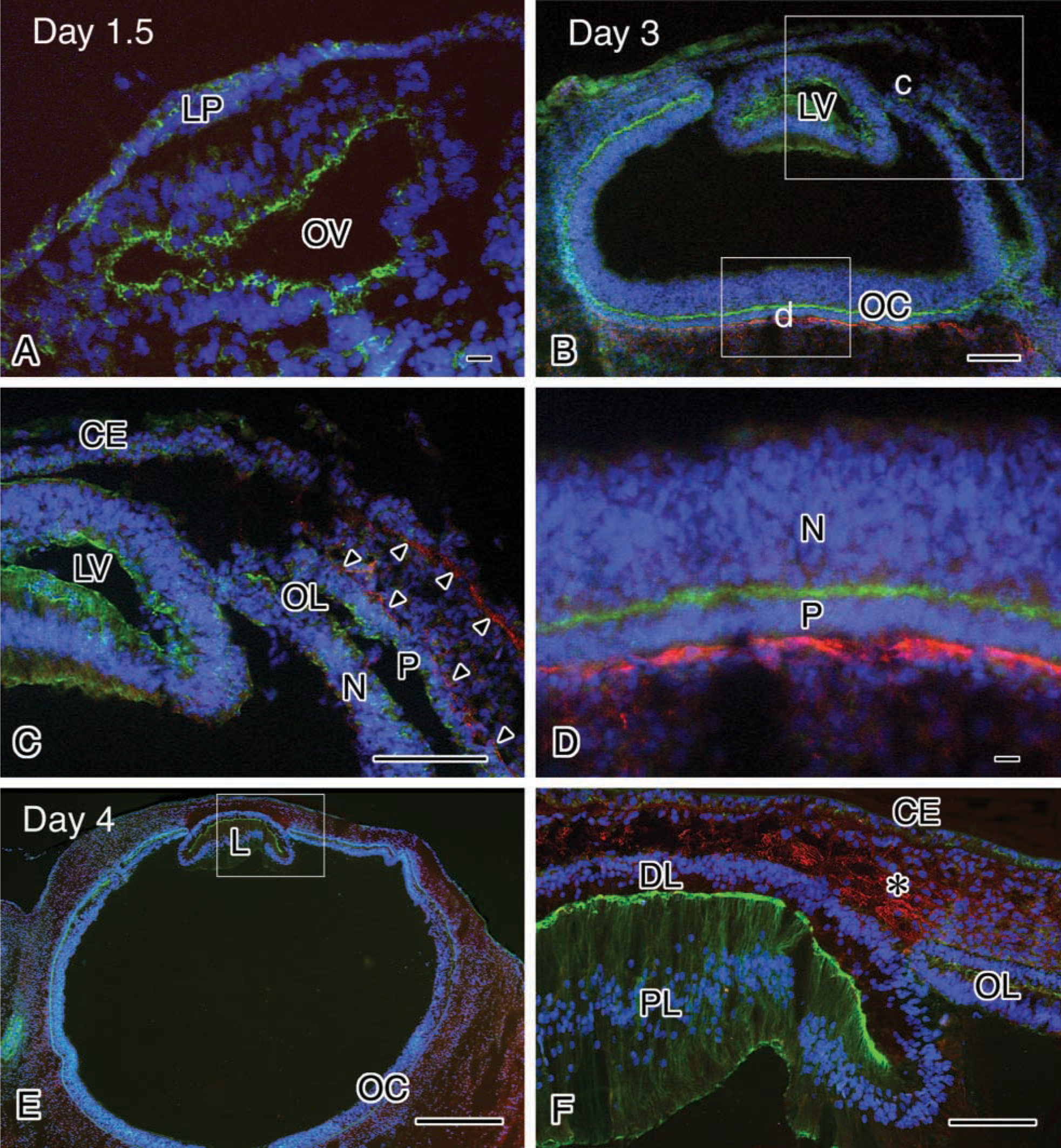
Immunohistochemical observation of the long form of Type XII collagen in eyes from day 1.5 (stage 10) (
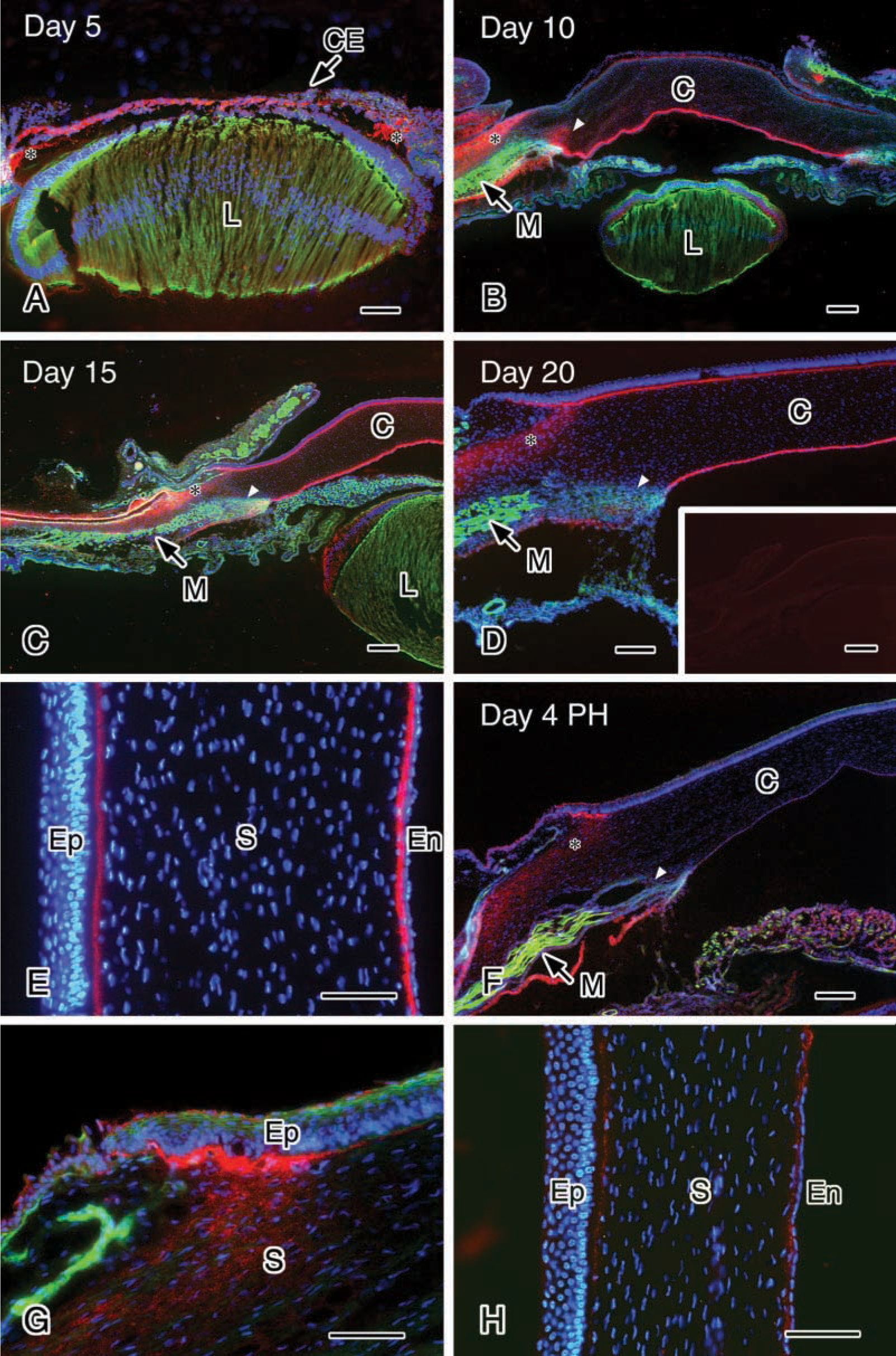
Immunohistochemical observation of the long form of Type XII collagen in eyes of Day 5 (stage 27) (
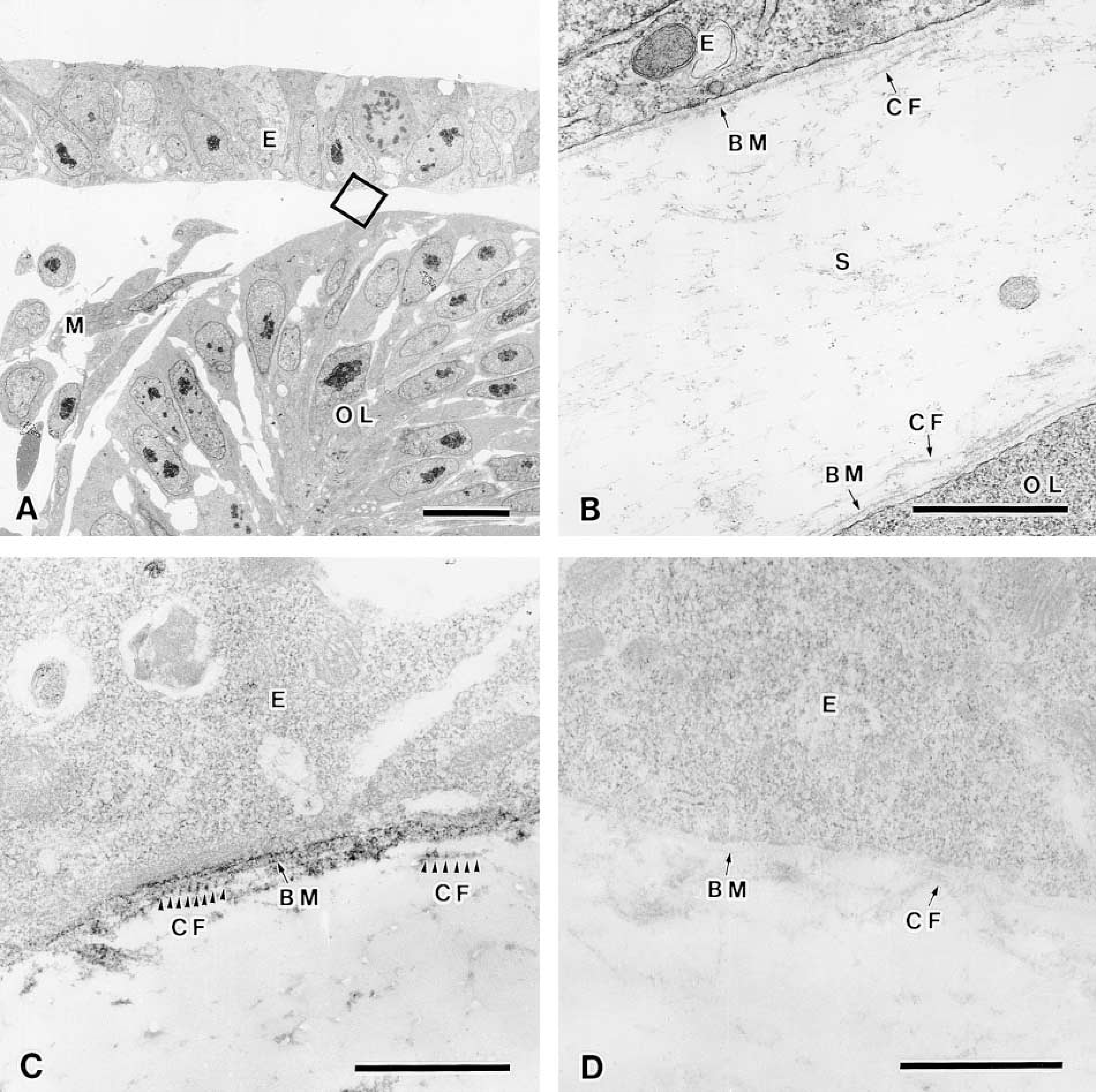
Immunoelectron microscopic observation of the long form of Type XII collagen in eyes of Day 3 chicken embryos. (
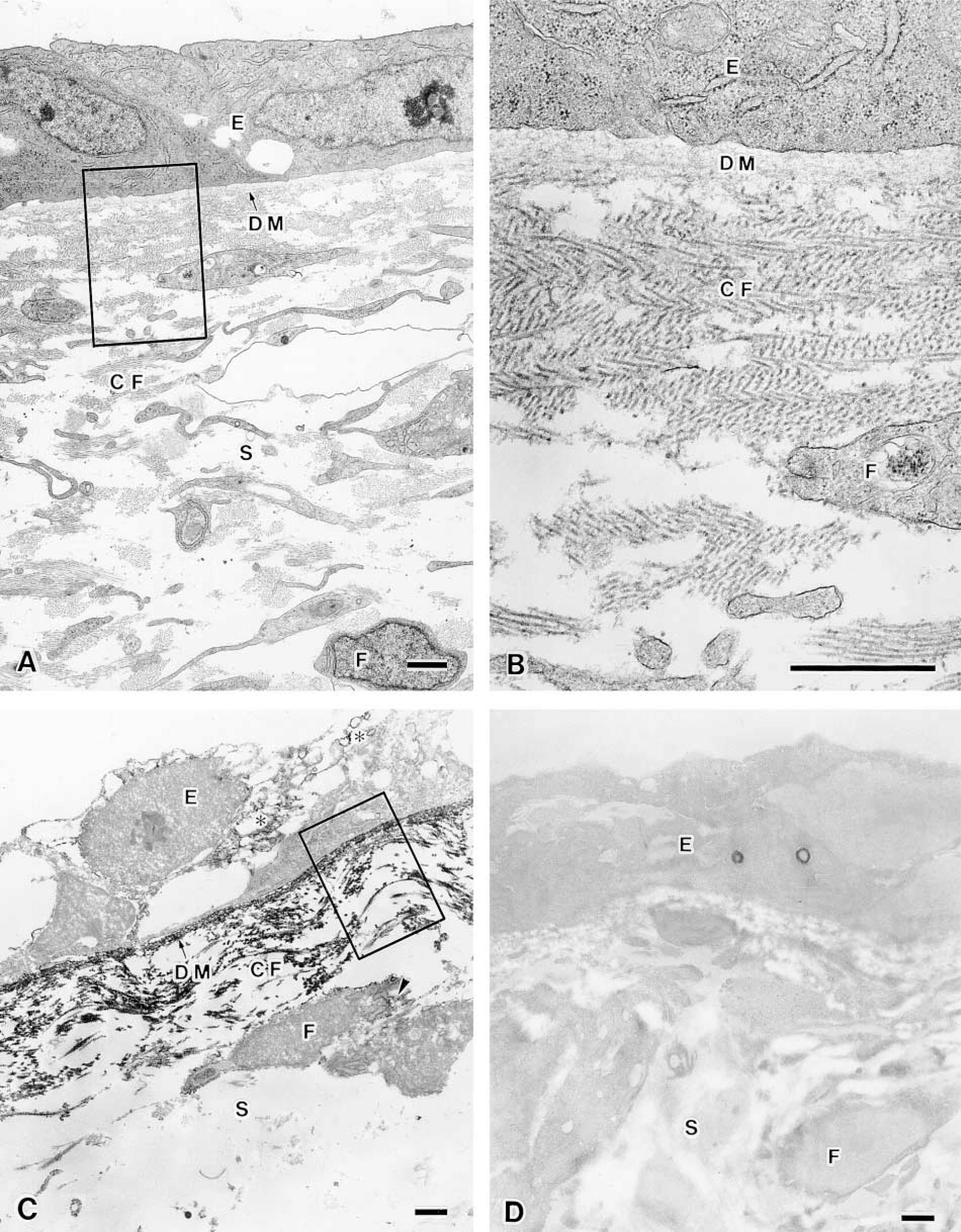
Immunoelectron micrographic observation of the long form of Type XII collagen in sub-endothelial matrix of Day 10 chicken embryonic cornea. (
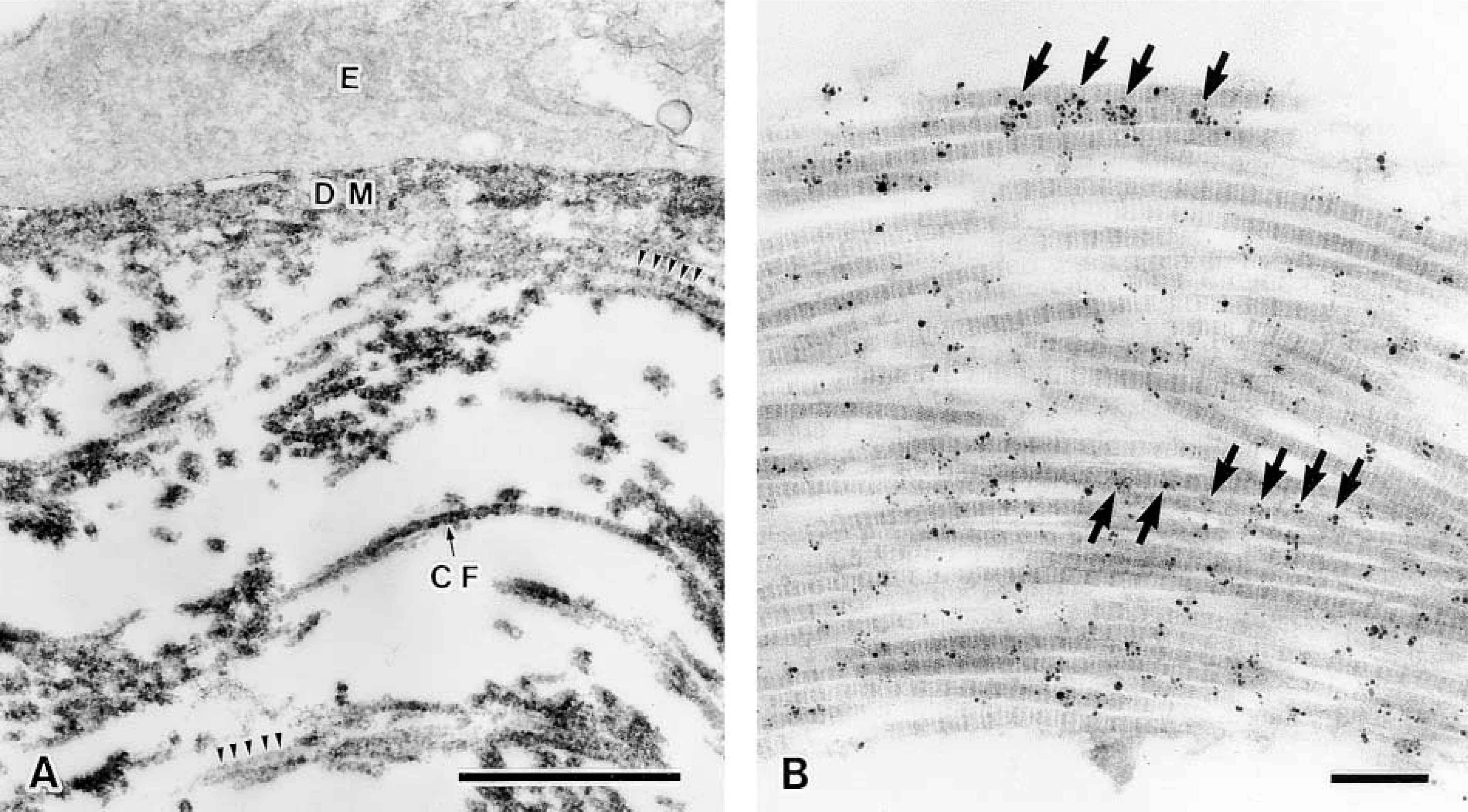
Immunoelectron micrographic observation of the long form of Type XII collagen in sub-endothelial matrix of Day 10 chicken embryonic cornea. (A) Enlargement of the area indicated by rectangle in Figure 6C. Immunoreactivities were detected by the HRP labeling method. (B) Immunoreactivities along the collagen fibrils were detected by colloidal gold labeling. Arrowheads in A and arrows in B show the 55–60-nm periodic immunoreactivity along the collagen fibrils. CF, collagen fibrils; DM, Descemet's membrane; E, endothelium. Bars: A = 1 μm;
Discussion
Type XII collagen contains the C-terminal region in which the triple-helical domains (COL1 and COL2) are interrupted by non-triple-helical domains (NC1 and NC2) (Shaw and Olsen 1991). The occurrence of two structural variants in the NC1 domain (the long form) and in the NC1 and NC3 domains (the short form) generated by tissue-specific alternative splicing has been reported in mouse and rat Type XII collagen molecules (Kania et al. 1999). The present study showed that the P3D11 monoclonal antibody immunoprecipitates two sets of doublet bands with molecular weights of 300 K and 600 K from chicken embryonic tendon fibroblast lysates. The 600-K doublet bands, as shown in Figure 2, appeared to be dimeric forms of the 300-K bands crosslinked by a yet unknown mechanism, as observed also in other types of collagen molecules (Koch et al. 1992). When collagen Type XII was pulse-labeled with [35S]-Met in cultured chicken tendon fibroblasts, the 300-K and 600-K doublet protein bands were detected with the antibody within 10 min in the lysates (data not shown), indicating that the doublets of the Type XII collagen are not the consequence of protein degradation. The doublets present at 300 K and 600 K probably represent alternatively spliced variants of the long form. Conversion of these protein bands to a single band with an approximate molecular weight of 270 K by bacterial collagenase digestion showed that the alternate splicing and intermolecular crosslinking sites are within the collagenase-digestive polypeptide region including the COL2, NC2, COL1, and NC1 domains. These results indicated that the 300-K doublet bands correspond to the isoforms Types XIIA-1 and XIIA-2 as shown in rat and mouse Type XII collagen molecules (Kania et al. 1999). Because monoclonal antibody P3D11 was raised against the N-terminal region of the NC3 domain, the antibody recognizes only the long form (300-K doublets) of Type XII collagen, equivalent to Types XIIA-1 and XIIA-2 and their dimeric forms.
In earlier studies on Type XII collagen during chicken corneal formation (Sugrue 1991; Gordon et al. 1996), it was not established when and where the long form of Type XII collagen is initially expressed, whether or not its expression changes after hatching when the eye begins to function, and whether the long form is present in the limbus. In the present study, we demonstrated that the long form of Type XII collagen is expressed initially on the optic cup and cornea on Day 3 of incubation (stage 19) when the lens vesicle pinches off, the overlying ectoderm is induced to become the corneal epithelium and the optic cup is completed. The expression levels of the long form decrease in the sub-endothelial and sub-epithelial matrices after hatching, and the long form is present in the limbus. That the long form of Type XII collagen localizes mainly in the sub-epithelial and sub-endothelial matrices during chicken embryonic development is consistent with the previous report using Day 5–18 chicken embryos and an antibody that recognizes the long and short forms (Gordon et al. 1996). It was also reported that immunoreactivity to the Type XII collagen antibody that recognizes both long and short forms is present in the limbus both anteriorly and posteriorly (Sugrue 1991; Gordon et al. 1996). However, in the present study we observed that the immunoreactivity of the long form was very weak after Day 10 in the posterior region of limbus that was termed by Sugrue (1991) the corneoscleral angle, whereas it was strong in the anterior region of the limbus. This indicates that the posterior region of limbus may be composed mainly of the short form of Type XII collagen.
Further studies on the localization of the short form are to be carried out by using the antibody specific to the form. Because the limbus is a transitional zone between the cornea and sclera and secretes aqueous humor, and because stem cells of the corneal epithelium are stored in the limbus, the long form of Type XII collagen expressed in the anterior region of the limbus could be involved in induction of the stem cells to differentiate and in adjustment of the tensile strength (Trächslin et al. 1999). The reasons why the expression levels of the long form decrease in the sub-epithelial and sub-endothelial matrices of the cornea after hatching and why its expression levels are unchanged in the anterior region of the limbus even after hatching remain to be elucidated.
The long form of Type XII collagen also exists in the cornea of other animal species (Sugrue 1991; Oh et al. 1993; Böhme et al. 1995; Zhan et al. 1995; Gordon et al. 1996; Wessel et al. 1997; Kato et al. 2000). For example, in rabbit it localizes in the corneal basement membrane but not in the corneal stroma (Anderson et al. 2000). In human it is present in the corneal stroma, epithelial basement membrane, and Bowman's membrane (Wessel et al. 1997). In bovine its expression is restricted to the corneal stroma and it is not present in the epithelial basement membrane and Descemet's membranes (Kato et al. 2000). Thus, the localization of the long form of Type XII collagen in the cornea is differs among species. This may indicate that the molecular mechanisms that form the eye are different from species to species. Further studies are required to establish this, together with the elucidation of discrete functions of the long and short forms of Type XII collagen, if any, during eye formation.
The present immunoelectron microscopic study revealed that the long form of Type XII collagen localizes in the basement membrane and along the collagen fibrils that are associated with basement membrane in chicken cornea. Interestingly, the long form of Type XII collagen was expressed periodically at 55–60 nm intervals along the surface of bundled collagen fibrils. Similar results were also obtained in human and bovine tissues (Keene et al. 1991; Wessel et al. 1997; Kato et al. 2000). Such localization indicates that the long form of Type XII collagen mediates periodic interactions between collagen fibrils and other extracellular matrix components to stabilize them. In fact, Types I, II, III, IV, V VI, IX, XIV, and XVII collagen molecules were detected immunohistochemically in avian cornea (Conrad et al. 1980; Linsenmayer et al. 1984, 1986; Birk et al. 1986; Fitch et al. 1988, 1990; Sugrue 1991; Chen et al. 1993; Watanabe et al. 1994; Gordon et al. 1996, 1997; Doane et al. 1998). Among them, Types IX and XIV are associated with collagen fibrils, Type IX collagen forms a complex with Type II collagen in the primary corneal stroma (Fitch et al. 1988, 1998; Cai et al. 1994), and structurally related Types XII and XIV collagen molecules also form complexes with Type I collagen (Koch et al. 1995). Quite interestingly, Type XIV collagen is distributed throughout the corneal stroma, whereas Type XII collagen is limited to the sub-epithelial and sub-endothelial regions and limbus (Gordon et al. 1996), indicating the discrete functions of Types XII and XIV collagen molecules.
Because the N-terminal and C-terminal regions of Type XII collagen have been shown to interact with cells and extracellular matrix components, the generation of the splicing variants during the development and differentiation could contribute to dramatic changes in the macromolecular interactions and their arrangements in connective tissues, which is quite important for the tissue construction. Therefore, the temporal and spatial expression of the long form of Type XII collagen, as observed in the present study, imply its crucial roles in chicken eye formation by a yet unestablished mechanism.
Footnotes
Acknowledgments
Supported by Grants-in-Aid for Scientific Research (10877004 to YA and 12680708 to KF) from the Ministry of Education, Culture, Sports, Science and Technology, Japan.
We wish to express our appreciation to Mr M. Fukuda, Ms S. Matsubara, Ms M. Kanai, and Ms T. Shibata in the Laboratory for Electron Microscopy and Department of Anatomy at Kyorin University School of Medicine for their technical assistance. We thank Dr A. Kudo for valuable discussions.