
Abstract
The vascular endothelial growth factor (VEGF) family is involved in angiogenesis, and therefore VEGFs are considered as targets for anti-angiogenic therapeutic strategies against cancer. However, the physiological functions of VEGFs in quiescent tissues are unclear and may interfere with such systemic therapies. In pathological conditions, increased levels of expression of the VEGF receptors VEGFR-1, VEGFR-2, and VEGFR-3 accompany VEGF activity. In this study we investigated normal human and monkey tissues for expression patterns of these receptors. Immunohistochemical staining methods at the light and electron microscopic level were applied to normal human and monkey tissue samples, using monoclonal antibodies (MAbs) against the three VEGFRs and anti-endothelial MAbs PAL-E and anti-CD31 to identify blood and lymph vessels. In human and monkey, similar distribution patterns of the three VEGFRs were found. Co-expression of VEGFR-1, −2, and −3 was observed in microvessels adjacent to epithelia in the eye, gastrointestinal mucosa, liver, kidney, and hair follicles, which is in line with the reported preferential expression of VEGF-A in some of these epithelia. VEGFR-1, −2, and −3 expression was also observed in blood vessels and sinusoids of lymphoid tissues. Furthermore, VEGFR-1, but not VEGFR-2 and −3, was present in microvessels in brain and retina. Electron microscopy showed that VEGFR-1 expression was restricted to pericytes and VEGFR-2 to endothelial cells in normal vasculature of tonsils. These findings indicate that VEGFRs have specific distribution patterns in normal tissues, suggesting physiological functions of VEGFs that may be disturbed by systemic anti-VEGF therapy. One of these functions may be involvement of VEGF in paracrine relations between epithelia and adjacent capillaries.
Keywords
V
VEGFRs are tyrosine kinases that mediate signaling in endothelial cells of blood and lymph vessels, which induce cell proliferation, survival, or differentiation. This signaling is required for normal development and maintenance of the vascular bed and for angiogenic responses under (patho)physiological conditions. VEGFRs are members of a receptor tyrosine kinase family that is rather specific for endothelial cells, consisting of at least three members: VEGFR-1 (Flt-1), VEGFR-2 (KDR), and VEGFR-3 (Flt-4). However, VEGFR-1 is also present in microvascular pericytes (Takagi et al. 1996) and in hematopoietic cells, such as monocytes (Mustonen and Alitalo 1995; Barleon et al. 1996). In addition to the transmembrane form of VEGFR-1, a soluble form of VEGFR-1 (sVEGFR-1) exists and is believed to act as an antagonist of VEGF activity. Ligands that bind to these VEGFRs are members of the VEGF family, which consists at present of six members: VEGF-A, VEGF-B, VEGF-C, VEGF-D, the viral VEGF homologue VEGF-E, and placental growth factor (Senger et al. 1983; Joukov et al. 1996; Achen et al. 1998; Olofsson et al. 1998; Witzenbichler et al. 1998). VEGF-A binds with high affinity to VEGFR-1 and VEGFR-2 (Vaisman et al. 1990; Mustonen and Alitalo 1995), VEGF-B and placental growth factor bind to VEGFR-1, VEGF-C and VEGF-D bind to VEGFR-2 and VEGFR-3, and VEGF-E binds to VEGFR-2. In pathological conditions such as cancer, VEGF activity is invariably associated with high levels of VEGFR expression (Brown et al. 1993b). High levels of expression of VEGFRs in a given tissue or anatomic site were therefore suggested to be a marker of VEGF activity (Veikkola et al. 2000).
VEGF-A mRNA is constitutively expressed by kidney glomerular epithelium, choroid plexus epithelium and retinal pigment epithelium (RPE) in mice, rats, and humans (Breier et al. 1992; Simon et al. 1995; Yi et al. 1998), and constitutive expression of VEGFR-1 and −2 has been found in endothelial cells of human adult kidney glomeruli (Simon et al. 1998). We have recently shown that RPE cells in vitro secrete VEGF-A at their basolateral (choroidal) side, whereas all three VEGFRs were expressed in vivo at the side of endothelial cells of the choriocapillaris that faces the RPE (Blaauwgeers et al. 1999). VEGF-A and the VEGFR-3 ligand VEGF-C may act here to induce a permeable phenotype and act as survival factors in a paracrine relation between RPE and the choriocapillaris, because endothelial cells of the choriocapillaris have fenestrations only at their side facing the RPE (Blaauwgeers et al. 1999) and experimental destruction of RPE leads to choriocapillaris atrophy (Korte et al. 1984).
On the basis of these findings, we hypothesized that the VEGF family and its receptors are involved in paracrine functions of epithelia regulating survival and permeability of adjacent endothelium in physiological conditions. To find further evidence for this hypothesis, we investigated the tissue distribution patterns of VEGFRs in the adult human and monkey, with special emphasis on endothelia adjacent to epithelia.
Materials and Methods
Tissue Samples
Normal human tissue samples were obtained from fresh surgical specimens and autopsies performed within 10 hr of death (Table 1). Cryostat sections of the tissue samples were judged by a certified pathologist and no indications of disease were found. Normal human eyes from 10 donors were kindly provided by the Corneabank Amsterdam. For donor eyes and autopsy tissues, the cause of death was unrelated to the tissues used. For electron microscopy, human tonsil samples (n = 3) were obtained with informed consent within 30 min after surgical removal to keep good morphology at the ultrastructural level. The use of human material was in accordance with the Declaration of Helsinki on the use of human material for research.
List of organs used for immunohistochemical localization of VEGFRs in normal human and monkey tissues
Normal tissue samples from two cynomolgus monkeys (Macaca fascicularis) (Table 1) were used for this study. Both animals had been used in behavioral studies in the past. All experiments were carried out in accordance with the guidelines established for animal care by the University of Nijmegen, The Netherlands.
All tissue samples were snap-frozen in liquid nitrogen and stored at −70C until used.
Light Microscopic Immunohistochemistry
Air-dried serial cryostat sections (8 μm thick) were fixed in cold acetone for 10 min and stained by an indirect immunoperoxidase procedure (Witmer et al. 2001), using the following antibodies: monoclonal antibodies (MAb) Flt-19 (against VEGFR-1, dilution 1:400) and KDR-1 (against VEGFR-2, dilution 1:400) (Simon et al. 1998), MAb 9D9F9 (against VEGFR-3, dilution 1:1500) (Lymboussaki et al. 1998), and the anti-endothelial MAbs PAL-E (dilution 1:1000) (Schlingemann et al. 1985) and EN-4 (against CD31; dilution 1:500) (Ruiter et al. 1989). Flt-19 and KDR-1 were kindly provided by Dr. H.A. Weich (National Research Center for Biotechnology; Braunschweig, Germany), 9D9F9 by Prof. K. Alitalo (Haartman Institute; Helsinki, Finland), and EN-4 was purchased from Sanbio (Uden, The Netherlands). In negative control incubations, the primary antibodies were omitted or replaced by an antibody against a non-human bacterial protein (mouse negative control immunoglobulins; Dako, Glostrup, Denmark) in the same dilutions. Peroxidase activity was visualized by incubation in a solution of 3-amino-9-ethyl carbazole (AEC, red color) or 3,3-diaminobenzidine (DAB, brown color) with 0.01% H2O2 as substrates in 0.1 M PBS (pH 7.4) for 10 min. Counterstaining was performed with hematoxylin.
To determine the extent of vascular staining and the type of positive vessels, the vascular markers PAL-E and anti-CD31 were used. Staining of serial sections with PAL-E was performed to identify capillaries and venules that are not involved in a blood-tissue barrier, whereas this antigen is absent in lymphatic endothelium. Anti-CD31 was used as a marker for all blood and lymph vessel endothelium (Schlingemann et al. 1985, 1997; Ruiter et al. 1989).
All sections were examined by two independent observers. Microvascular staining was graded as follows: no staining (-), weak staining (±), distinct staining (+), intense staining (+ +), very intense staining (+ + +).
Electron Microscopic Immunohistochemistry
For electron microscopy, samples of tonsil tissue were fixed for 40 min at room temperature (RT) in 2% paraformaldehyde in Sørensen's phosphate buffer (pH 7.4) (Ruiter et al. 1989; Schlingemann et al. 1991). After fixation, tissue samples were washed in 0.1 M PBS, pH 7.4, incubated overnight in a 20% sucrose-PBS solution at 4C, snap-frozen in liquid nitrogen, and stored at −70C until use.
A pre-embedding immunoperoxidase technique was used to demonstrate subcellular distribution patterns of VEGFR-1 and VEGFR-2 in tonsil tissue as described previously (Ruiter et al. 1989; Schlingemann et al. 1990; Witmer et al. 2001). Cryostat sections (60 μm thick) were cut from the paraformaldehyde-fixed samples, washed in PBS, and incubated for 1 hr in a blocking buffer containing normal horse serum (Vector; Burlingame, CA) with 0.05% saponin. Then free-floating sections were incubated with MAbs against VEGFR-1 and VEGFR-2 (see above) for 1 hr at RT and then overnight at 4C. Control sections were incubated with MAb PAL-E (see above) as a positive control, or with the antibody against the non-human bacterial protein in the same dilutions as the negative control (see above). After extensive washings in PBS, sections were incubated with biotinylated horse anti-mouse immunoglobulins for 1 hr at RT. Sections were washed and incubated with a streptavidin-horseradish peroxidase complex for 90 min at RT. Sections were washed for 2 hr in Tris-HCl buffer (pH 7.8) and peroxidase activity was demonstrated by incubation in a solution of DAB and 0.01% H2O2 for 10 min. The incubation was terminated by rinsing with distilled water. Sections were then postfixed in 1% OsO4 supplemented with 1% ferricyanide in 0.1 M sodium cacodylate buffer for 30 min, dehydrated in a series of graded ethanols, and flat-embedded in epoxy resin. Ultrathin sections (90–160 nm thick) were cut and photographed using a Philips 201 electron microscope (Philips; Eindhoven, The Netherlands) at 80 kV. Counterstaining was omitted.
Results
Light Microscopy
Staining patterns of all VEGFRs were similar in human and monkey tissues and are summarized in Table 2.
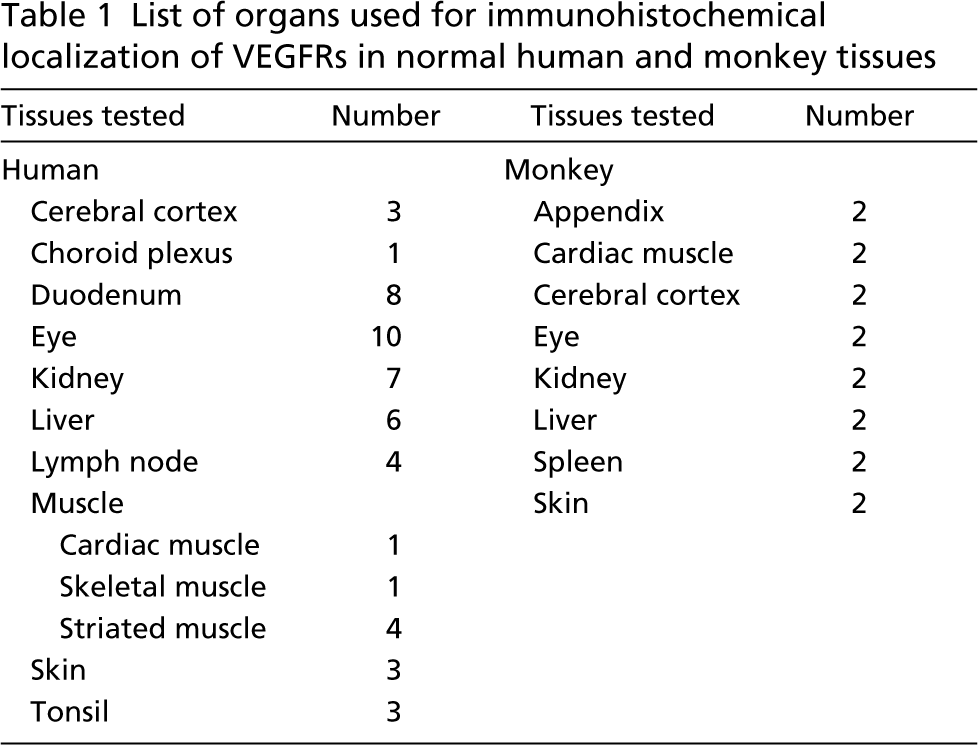
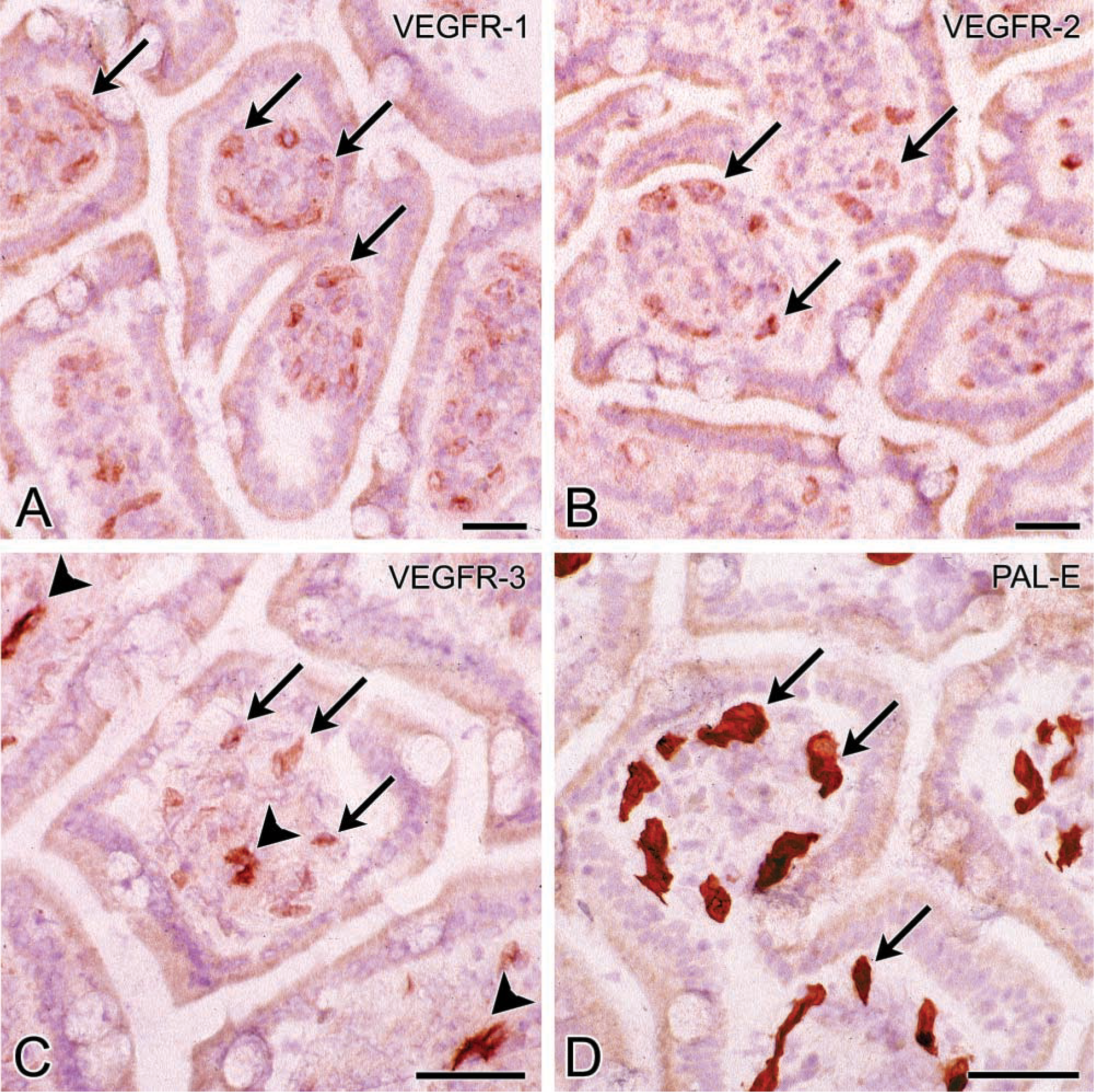
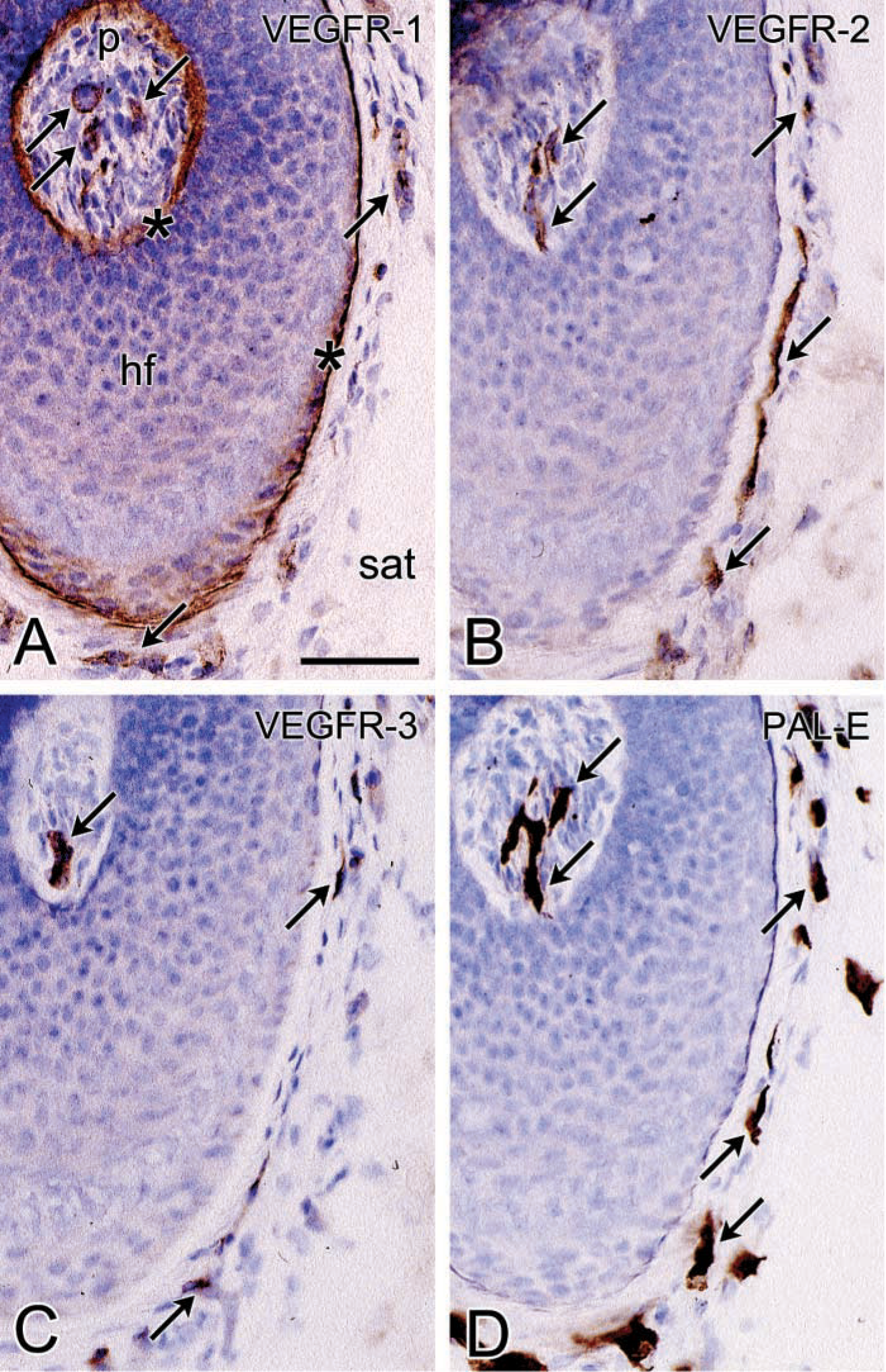
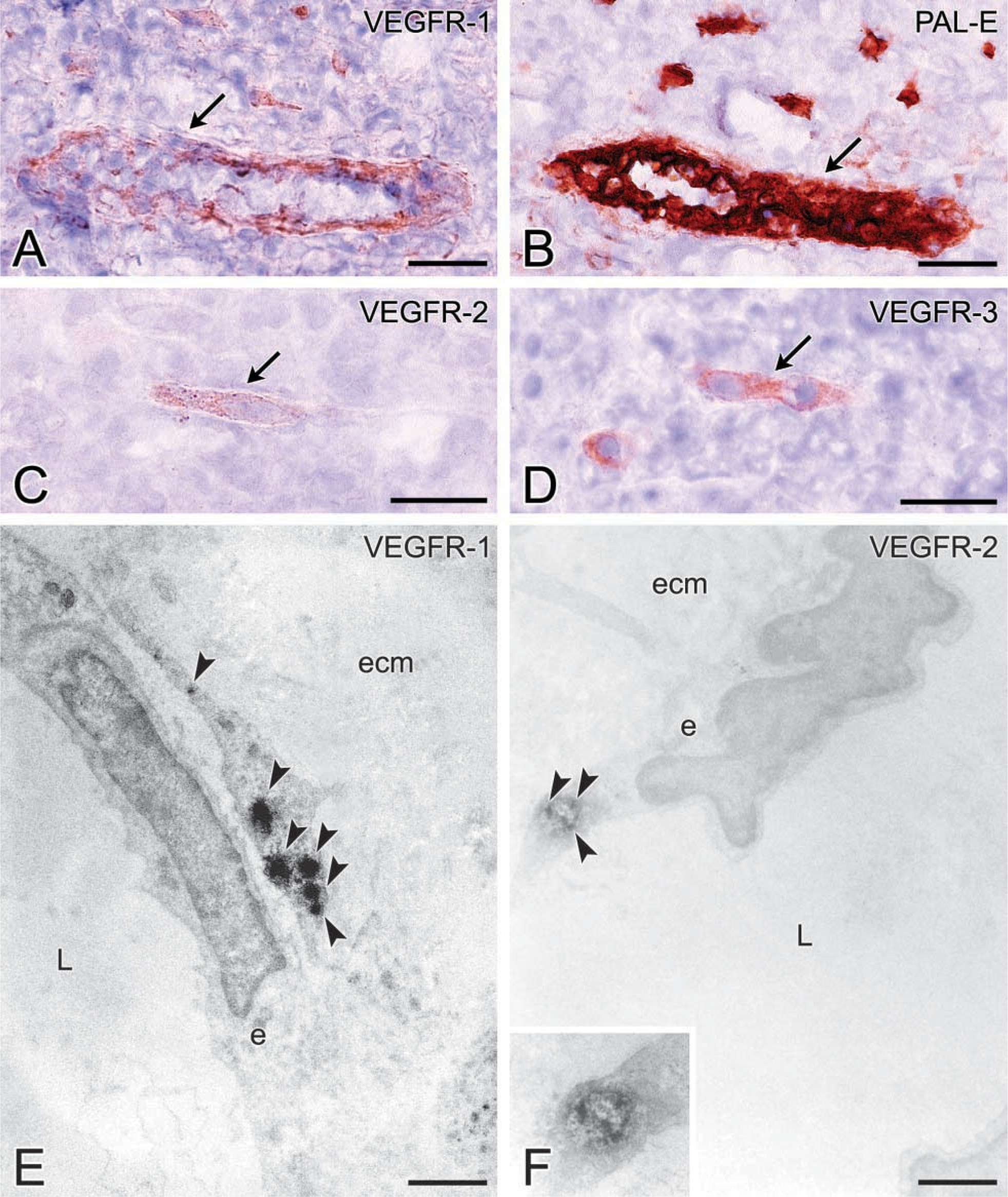
VEGFR-1. Weak to intense granular staining of VEGFR-1 was found in capillaries adjacent to epithelia such as the choriocapillaris, capillaries of ciliary processes in the eye, capillaries in the lamina propria of gastrointestinal mucosa (Figure 1), kidney glomeruli, capillaries surrounding hair follicles and within the papilla of hair follicles (Figure 2), capillaries in portal tracts close to bile ducts in liver, and capillaries of choroid plexus (Table 2). In addition, capillaries in the CNS showed intense staining (Figure 3). Capillaries not adjacent to epithelia, and larger vessels, such as arterioles, venules, arteries, and veins, were negative or only weakly stained for VEGFR-1. In lymphoid tissues, intense staining of VEGFR-1 was observed in capillaries, high endothelial venules, and sinusoids (Figure 4). The distribution pattern of the VEGFR-1 staining product in this tissue gave the impression of a localization of this receptor in microvascular pericytes and/or at the abluminal side of endothelial cells of blood vessels. In these cases, unstained endothelial cells were found at the luminal side of small and high endothelial venules (Figure 4). Furthermore, non-vascular VEGFR-1 staining was observed in basement membranes surrounding kidney tubuli, bile duct epithelium in portal tracts of the liver, in glassy membranes of hair follicles (Figure 2), which is the basement membrane separating epithelium from connective tissue of the follicle, and in the inner limiting membrane of the retina (Witmer et al. 2002).
Staining patterns of VEGFs in normal human and monkey tissues


Tested only in human tissue.
Witmer et al. in press.
Tested only in monkey tissue.
VEGFR-2. Similar to VEGFR-1, larger vessels such as arterioles, venules, arteries, and veins were negative or weakly stained for VEGFR-2. Granular staining of VEGFR-2 was always found in VEGFR-1-positive capillaries adjacent to epithelia and lymphoid tissues (Figures 1, 2, and 4; Table 2). In lymphoid tissues, staining of VEGFR-2 was weak in arterioles and venules and distinct in sinusoidal endothelium. In the CNS, staining of VEGFR-2 was absent in blood vessels, in contrast to VEGFR-1 (Figure 3).
VEGFR-3. Co-localization of VEGFR-3 with VEGFR-1 and VEGFR-2 was found in capillaries adjacent to epithelia and capillaries of lymphoid tissues (Figures 1, 2, and 4; Table 2). In lymphoid tissues, staining of VEGFR-3 was weak in arterioles, distinct in venules and veins, and intense in sinusoidal endothelium. Staining of VEGFR-3 in blood vessels displayed a fine granular intracellular pattern. VEGFR-3 staining was absent in blood vessels in the CNS (Figure 3).
Very intense staining of VEGFR-3 was observed in thin-walled lymphatic vessels in portal tracts of the liver, gastrointestinal villi (Figure 1), kidney, skin, and lymphoid tissues. These vessels were recognized in serial sections on the basis of their PAL-E negativity.
Non-vascular staining of VEGFR-3 was present in cerebral parenchyma (Figure 4), consistent with diffuse VEGFR-3 staining in neural elements of the retina (Witmer et al. 2002).
Electron Microscopy
In tonsil tissue, staining of both VEGFR-1 and VEGFR-2 was found in capillaries. Staining of VEGFR-1 was found in pericytes (Figure 4E), whereas staining of VEGFR-2 was restricted to endothelial cells (Figure 4). VEGFR-1 and VEGFR-2 staining was found in a dot-like configuration in the cytoplasm (Figures 4E and 4F), which is in agreement with the granular staining pattern of VEGFRs at the light microscopic level (Figures 4A and 4C). In addition, a more diffuse precipitation in the cytoplasm was observed.
Photomicrographs of cryostat sections of human duodenum stained for VEGFR-1 (
Discussion
The present study demonstrates (a) co-localization of VEGFR-1, −2, and −3 in capillaries adjacent to epithelia in various normal human and monkey tissues, in addition to VEGFR-3 expression in lymphatic vessels, (b) expression of VEGFR-1, −2, and −3 in microvessels in lymphoid tissues, (c) expression of VEGFR-1, but not of VEGFR-2 and VEGFR-3, in capillaries in brain and retina, (d) non-vascular soluble VEGFR-1 expression in basement membranes of bile ducts and hair follicles, and (e) at the ultrastructural level, expression of VEGFR-1 in pericytes and expression of VEGFR-2 in endothelial cells in capillaries of tonsils.
Photomicrographs of serial cryostat sections of skin of the human scalp stained for VEGFR-1 (
In addition to our previous observations of VEGFR expression in the choriocapillaris in the eye (Blaauwgeers et al. 1999), we found expression of VEGFR-1, −2, and −3 in capillaries adjacent to epithelia throughout the human and monkey body. In tumors in humans and experimental animals, high levels of expression of VEGFRs usually accompany VEGF activity (Plate et al. 1992; Brown et al. 1993a, b, 1995; Hatva et al. 1995, 1996; Takahashi et al. 1995; Warren et al. 1995). Expression of VEGFRs may therefore indicate local VEGF activity. In the present study of expression of VEGFRs under orthophysiological conditions, localization of VEGFR-1 and −2 in endothelia adjacent to epithelia corresponded with expression of VEGF-A mRNA that has been described in some of these epithelia, such as choroid plexus, kidney glomeruli (Breier et al. 1992; Esser et al. 1998), gastrointestinal epithelium (Shifren et al. 1994; Terris et al. 1998), and epithelium of hair follicles (Shifren et al. 1994; Kozlowska et al. 1998; Yano et al. 2001). Localization of VEGFR-3 in endothelium of gastrointestinal mucosa is in line with expression of VEGF-C mRNA that was found in small intestine (Joukov et al. 1996). Our observations are also in line with expression patterns of mRNA of VEGFR-1 and −2 in endothelial cells of adult choroid plexus and kidney glomeruli (Millauer et al. 1993; Simon et al. 1995).
Photomicrographs of cryostat sections of human cerebral cortex stained for VEGFR-1 (
Taken together, our results and data available in the literature suggest that VEGFs have physiological functions in these human tissues. This is important because such functions may be disturbed by therapeutic anti-VEGF strategies in patients with cancer or eye disease. It was recently shown that chronic VEGF inhibition indeed leads to apoptosis of alveolar cells in the lungs of experimental animals (Kasahara et al. 2000). However, other experimental studies of effects of VEGF-A inhibition in rodents were unable to demonstrate gross toxicity, arguing against a physiological role of VEGF-A in adult rodents (Fong et al. 1999; Gerber et al. 1999; Robinson et al. 2001), findings that may, however, not be relevant to humans.
Studies of VEGFR expression patterns at the protein level in normal human tissues are scarce. Simon et al. (1998) demonstrated expression of VEGFR-1 and VEGFR-2 in kidney glomerular capillaries, and expression of VEGFR-3 has been found almost exclusively in lymphatic vessels, with the exception of a few fenestrated endothelia (Partanen et al. 2000; Lymboussaki et al. 1998). Our observations on VEGFR-3 expression are largely in agreement with previous reports, but we found a more widespread distribution pattern of VEGFR-3, both in endothelia adjacent to epithelia and in lymphoid tissues. Interestingly, VEGFR-3 expression in blood vessels always co-localized with VEGFR-2 expression, allowing a synergistic role for different members of the VEGF family in these sites. On the other hand, both receptors are not present in microvessels of brain and retina, whereas VEGFR-1 is constitutively expressed in these capillaries. Other authors have similarly shown that during angiogenesis in the embryonic brain, high initial expression of VEGFR-2 is present, whereas in the adult brain, when angiogenesis has ceased, VEGFR-2 expression is not detectable (Kremer et al. 1997).
Photomicrographs (
In addition to a localization near epithelia, we also observed increased VEGFR expression in lymphoid tissues. VEGF-A mRNA expression was previously reported in normal tonsil by in situ hybridization (Foss et al. 1997). In normal lymph nodes, low expression of VEGFs was also found (Niki et al. 2000; Nishi and Maruyama 2000). Our observation of distinct VEGFR expression in lymph nodes and tonsils suggests that the VEGF/VEGFR signaling system plays a permanent role in these highly vascularized and dynamic tissues.
At the light microscopic level, VEGFR-1 staining of capillaries was found in a tube-like pattern, suggesting staining of pericytes and/or the abluminal side of endothelial cells. At the electron microscopic level, we found staining of VEGFR-1 in tonsil pericytes only and not in endothelial cells, whereas staining of VEGFR-2 was localized in endothelial cells only, as reported by others (Feng et al. 2000). These findings are consistent with observations of VEGFR-1 expression in pericytes in vitro (Takagi et al. 1996) and indicate that, in vivo, VEGFR-1 is predominantly a pericyte receptor for VEGF. Subcellular expression of VEGFR-3 was previously found only in endothelial cells (Witmer et al. 2001). Staining patterns at the electron microscopic level of VEGFR-1 and VEGFR-2 were found to be in a dot-like configuration in the cytoplasm of cells, consistent with the granular staining pattern in capillaries at the light microscopic level. These intracellular structures could not be identified but they may represent storage vesicles containing VEGFR under quiescent conditions, rather than vesiculo-vacuolar organelles (VVOs) (Feng et al. 2000) on the basis of morphology. In the case of pericyte or endothelial cell stimulation, these vesicles may be “shuttled” to the cell membrane where VEGFR is redistributed to be able to bind its ligands (Qu Hong et al. 1995).
In addition to vascular VEGFR-1 expression, non-vascular extracellular VEGFR-1 expression was observed in the basement membranes of kidney tubuli, bile ducts, and hair follicles, and in the inner limiting membrane of the retina, i.e., the basement membrane of Müller cells (Witmer et al. 2002). This may represent the soluble form of VEGFR-1, which was suggested to regulate availability of VEGF (Hornig et al. 2000).
Non-vascular VEGFR-3 expression was observed in cerebral parenchyma. It remains to be elucidated whether this represents a specific localization of this receptor in neural elements or crossreactivity of the antibody used. Similar non-vascular VEGFR-3 expression was found in neural elements of the retina (Wit-mer et al. 2002).
The results of the present study provide important clues with respect to the role of the VEGF family under physiological conditions. In the CNS, initial signaling of VEGF may occur via VEGFR-1 on pericytes and/or endothelial cells, allowing tight control of VEGF activity. Via this signaling pathway, VEGFR-2 and VEGFR-3 may be switched on under pathological conditions to enhance vascular permeability or to induce angiogenesis (Ortega et al. 1997, 1999), as we have recently shown in the retina (Witmer et al. 2002). Epithelia may require permanent VEGF signaling in a paracrine relation to maintain a permeable or fenestrated phenotype in adjacent capillaries, and thus the need of constitutive VEGFR-2 and −3 expression.
Several anti-angiogenic agents, alone or in combination with conventional therapies, are now in clinical trials (see the Internet site http://cancertrials.nci.nih.gov), including drugs that block VEGF and VEGFR signaling. However, long-term side effects are not yet known and it has already been implied that these therapies could affect normal tissues and physiological angiogenesis (Carmeliet and Jain 2000; Oosthuyse et al. 2001; Yano et al. 2001). In the present study, we show that VEGFRs are indeed expressed at specific sites, suggesting orthophysiological functions of VEGFs that may be disturbed by anti-VEGF therapy.
Footnotes
Acknowledgments
Supported by the Haagsch Oogheelkundig Fonds, the Landelijke Stichting voor Blinden en Slechtzienden, the Donders Fonds Utrecht, the Edmond and Marianne Blaauwfonds, and the Diabetes Fonds Nederland (grants 95.103 and 99.050).
We wish to thank Prof Dr K. Alitalo for providing the antibody against VEGFR-3, Prof Dr C.J.F. van Noorden for critically reading the manuscript, the Department of Pathology, Academic Medical Center, Amsterdam, The Netherlands (Head Prof Dr J.J. Weening), and Dr D. Nijdam (Department of Otolaryngology, Academic Medical Center, Amsterdam) for their assistance in obtaining the tissues, and W. Meun and T. Put for preparing the microphotographs.