
Abstract
The histological diagnosis of transmissible spongiform encephalopathies (TSEs), such as scrapie and bovine spongiform encephalopathy (BSE), relies on identification in the brain of spongiosis, gliosis, and neuron loss without inflammatory lesions. Because of its sensitivity, immunohistochemistry of abnormal prion protein (PrPsc) is of great help in this diagnosis and can be used on its own or complementary to the biochemical detection of PrPsc. However, in some cases no formalin-fixed material is available, rendering its use as a complementary method impossible. For that purpose, we studied the possibility of detecting PrPsc immunohistochemically in fixed brain samples that had been previously frozen and used for Western blotting analysis. We compared freshly and fixed-frozen brain samples originating from the same sheep, either affected or unaffected with scrapie. We also studied fixed-frozen brain samples from scrapie-affected goats and from cows showing BSE. We showed that in all the species tested, despite damage to the histological structures, PrPsc was still detectable in the fixed-frozen brain sections without unspecific background staining. Notwithstanding the limited number of cases thus far analyzed, we have already demonstrated the possibility of using PrPsc immunohistochemistry on fixed-frozen brain samples with very good efficacy, thus rendering possible its use for diagnostic purposes in TSEs.
S
IHC is actually a more effective tool than conventional histology, as shown for the diagnosis of scrapie in sheep (Miller et al. 1994). It has been reported that the topography of PrPsc immunolabeling corresponds to the distribution of vacuolar changes in BSE brains (Wells and Wilesmith 1995). In addition, IHC analysis of the topographical distribution of PrPsc may play an important role in the recognition of TSE strains (Foster et al. 1996; Wood et al. 1997; Orge et al. 2000; Caramelli et al. 2001).
In addition, IHC offers the possibility of a complementary approach to the diagnosis of TSEs based on the biochemical detection of PrPsc. It is particularly useful when weakly positive cases, as detected by Western blotting, need to be clarified. In these cases, when fixed tissues are not available it is of great interest to know to what extent the application of IHC on frozen brain samples is possible. For that purpose, these frozen brain samples need first to be postfixed and then embedded in paraffin.
In the present study we tested the possibility of applying PrPsc IHC to frozen samples of sheep brains, both scrapie-affected and normal, after formalin post-fixation. We compared PrPsc immunolabeling of freshly fixed obex with adjacent slices of tissue from the same sheep, frozen and then fixed. We also extended the trial to previously frozen brain samples from scrapie-affected goats and from BSE-affected cattle. We showed that, in all the species tested and despite damage to the histological structures, PrPsc was still detectable in the fixed-frozen brain sections without nonspecific background staining.
Materials and Methods
Animals
Sheep. Three sheep collected from the same flock (Drôme, France) were used as follows. Brains from two sheep clinically affected with scrapie and one from a healthy sheep were each cut sagittally into two parts. One half was cut in slices, put in Petri dishes, frozen, and maintained at — 80C until use for biochemical detection of PrPsc. The other half was formalin-fixed (10% in PBS 0.1 M) and maintained in fixative until use for this study. The two scrapie-positive brains were known to give a strong and a moderate PrPsc immunolabeling using Western blotting.
Goats. Four frozen brain samples (cerebellum and cerebral cortex) of scrapie-affected goats were used. These had been collected through the French scrapie surveillance scheme, and were stored at — 80C. Freshly formalin-fixed tissues were not available from these animals.
Cows. Four frozen brain samples (brainstem and spinal cord) acquired through the French BSE surveillance program of cattle at risk (aged 24 months and over, dead on farm or subjected to emergency slaughter due to sickness or accident) were added to the study (Calavas et al. 2001). These frozen samples were maintained at either —20C or —80C. Two brains were BSE-positive using the Prionics test (Schaller et al. 1999) and two were Prionics-negative.
Tissue Preparation
All frozen tissues had been thawed before this study because they had previously been used for biochemical PrPsc detection (Western blotting or Prionics). Slices of 5-mm thickness were cut at the level of the obex from both frozen and fixed brains. Similarly sized slices were cut from the bovine brains (obex and spinal cord) and from the goat tissues (cerebellum and cerebral cortex).
Thawed cuts were quickly transferred to a bath of buffered 10% formalin and fixed for 48 hr. Then all the samples were placed in tissue-embedding cassettes, put in a bath of formic acid (98–100%; Merk Eurolab, Darmstadt, Germany) for 1 hr at room temperature (RT) to reduce infectivity (Taylor et al. 1997), and then automatically processed through graded alcohols to chloroform and embedded in paraffin (Bayer; Cergy-Pontoise, France). Brain sections of 5-μm thickness were collected on treated glass slides (Star-frost; Medite Histotechnic, Burgdorf, Germany) and kept overnight in an oven at 55C. The brain slices were then de-waxed and rehydrated in graded alcohols.
Immunohistochemistry
Special care was taken because of resistance of brain slices previously weakened by freezing to the drastic pretreatments essential for PrPsc IHC (Bell et al. 1997; Taylor 2000). In this study we used a combination of pretreatments that enhance antigen retrieval and, at the same time, suppress the recognition of normal cellular PrP (PrPc) (Haritani et al. 1994; Van Everbroeck et al. 1999). The brain sections were first treated with 98–100% formic acid for 10 min at RT, followed by hydrated autoclaving for 20 min at 121C in water (Prestige Medical; Blackburn Lane, UK). Proteinase K at 20 μg/ml (Roche Diagnostics; Meylan, France), was then applied for 15 min at 37C. PrPsc immunostaining using SAF84 monoclonal primary antibody (0.5 μg/ml in PBS 0.1 M, pH 7.4/0.1% Triton X-100) was performed on the sheep samples using an automated immunostainer according to the recommendations of the manufacturers (NexES; Ventana Medical Systems, Tucson, AZ). Final revelation was achieved using a diaminobenzidine kit (DAB; Ventana, Illkirch, France). Goat and bovine brain slices were immunostained for PrPsc using the SAF84 antibody as previously described without any modification (Bencsik et al. 2001; Debeer et al. 2001).
Finally, the sections were dehydrated, mounted using Eukitt, and observed under a microscope (Zeiss; Oberkochen, Germany) coupled to an image analysis workstation (Biocom; Les Ulis, France).
Omission of primary antibodies was used to check the nonspecific background staining in scrapie and BSE cases. The specificity of positive PrPsc immunolabeling was also assessed using the scrapie- and BSE-negative brains.
Results
In the freshly fixed obex of one of the scrapie sheep (Figure 1B), intense PrPsc immunolabeling was observed in gray matter nuclei, such as the dorsal nucleus of the vagus nerve (DNV), the nucleus of the solitary tract (NST), the olivary nucleus (ON), the spinal tract of the trigeminal nerve (STN), and the reticular formation (RF). The cuneate (CN) and hypoglossal (HN) nuclei were stained to a lesser extent and white matter was devoid of any immunostaining. Nonspecific background was absent. Neither tears nor retractions were visible at higher magnification but vacuolar lesions were identifiable, i.e., in DNV (Figure 1D). In the previously frozen half obex of the same sheep (Figure 1A), intense PrPsc immunolabeling was also observed macroscopically in the DNV, NST, CN, and RF, whereas the ON and HN were not accessible. The tissue had a marked streaked aspect, showing tears and retractions in the parenchyma surrounding shrunken neurons (Figure 1C).
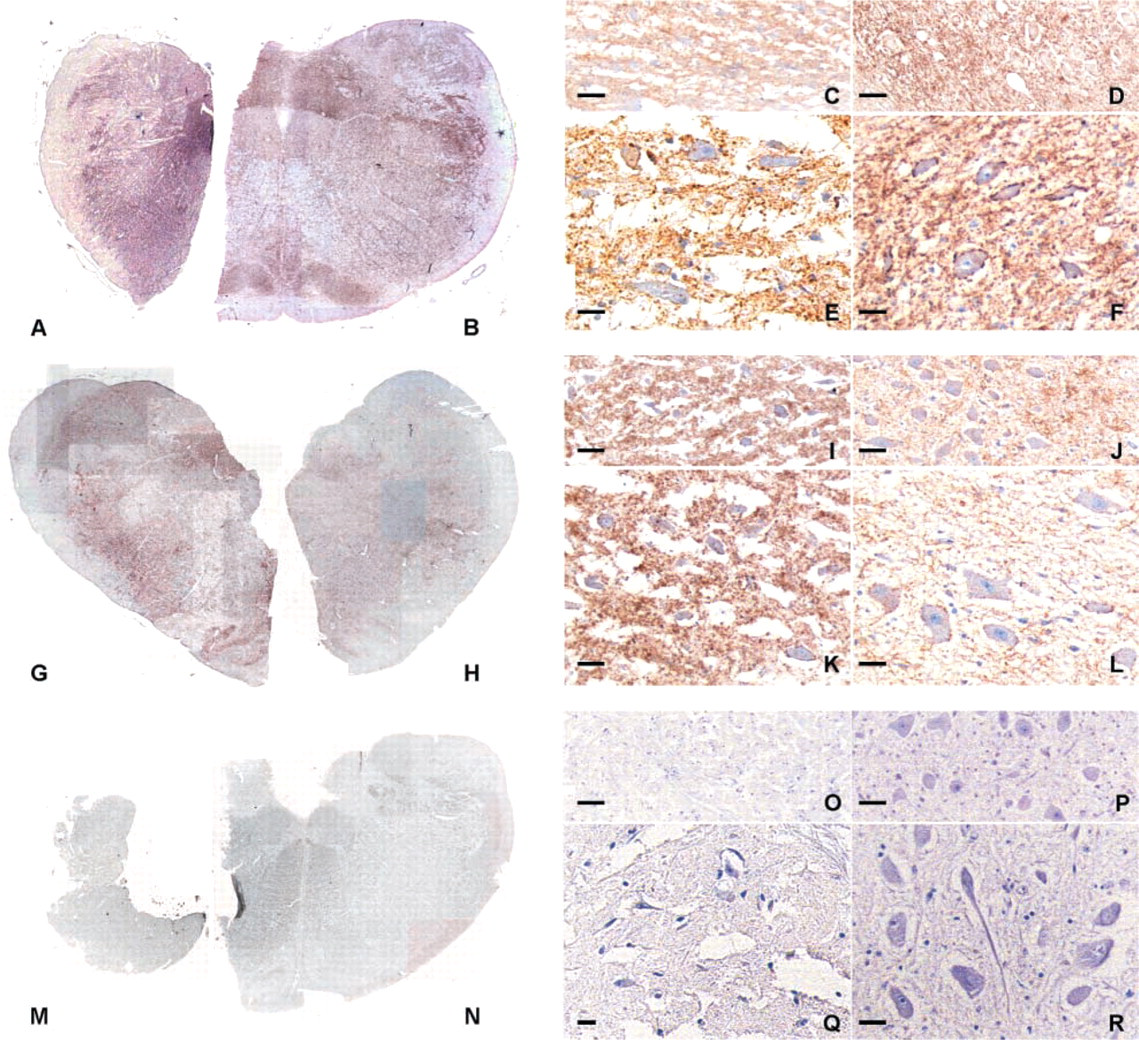
PrPsc immunoreactivity (brown) in fixed-frozen (
In the other scrapie sheep brains, the intensity of PrPsc immunostaining was very faint in the freshly fixed part (Figure 1H) but PrPsc deposits were distinguished at a higher magnification in the nuclei cited above, i.e., DNV, NST, CN, ON, and RF. In the previously frozen part corresponding to the same animal, PrPsc immunolabeling was more intense than in the freshly fixed part, making the DNV, NST, CN, RF and ON clearly identifiable macroscopically (Figure 1G), and tissues were streaked (Figure 1I) compared to the freshly fixed side (Figure 1J).
At the cellular level, PrPsc deposits were laid out as fine granules either scattered in the neuropil, in perikarya, or distributed perineuronally as a ring of pearls. This pattern was observed in scrapie-positive obex both freshly fixed (Figures 1F and 1L) and previously frozen (Figures 1E and 1K).
No PrPsc deposits or nonspecific background were seen in the obex of healthy sheep either fixed (Figures 1N, 1P, and 1R) or previously frozen (Figures 1M, 1O, and 1Q).
In the previously frozen cerebellum of scrapie-affected goats, PrPsc immunostaining was seen in the central white matter and in the granular and molecular layers of cerebellar gray matter (Figure 2A). Among retractions, granular PrPsc deposits appeared as a delineation of neuronal processes scattered within the neuropil of the medial cerebellar nucleus (Figure 2B) or within the cytoplasm of neurons (Figure 2C). In the cerebral cortex, the distribution and intensity of PrPsc immunostaining were irregular throughout the section (Figure 2D). In the middle of important lacerations, particulate and stellate PrPsc deposits were observed in the parenchyma (Figures 2E and 2F).
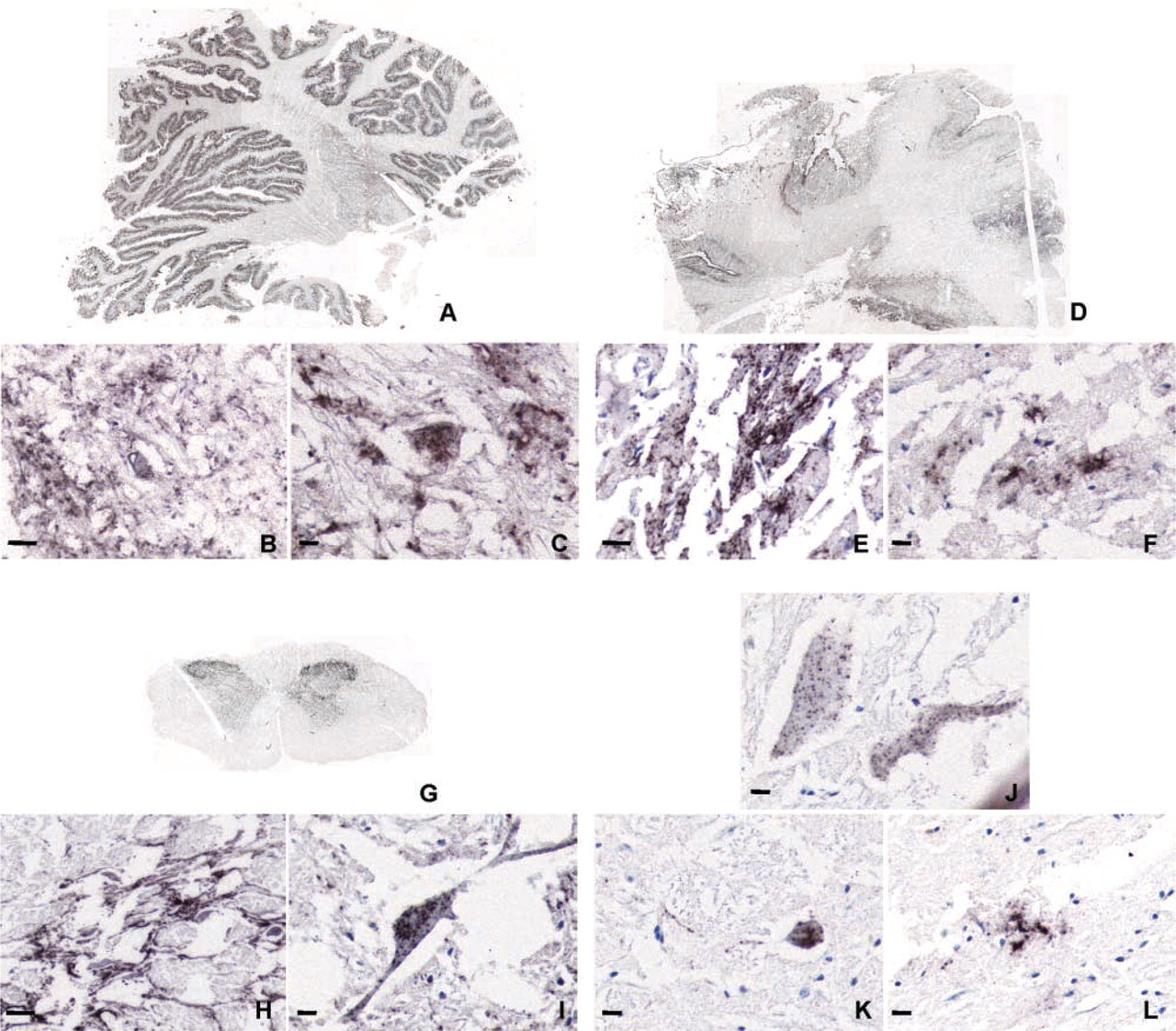
PrPsc immunohistochemistry in samples frozen before fixation from cerebellum (
In the previously frozen spinal cord of BSE-positive cattle, PrPsc deposits were restricted to the gray matter (Figure 2G). At a higher magnification, significant cytological damage was apparent. PrPsc granular delineation of neuronal processes (Figure 2H) and PrPsc deposition in neuronal perikarya were seen (Figure 2I). This pattern was also observed in a case in which the brainstem was weakly positive in the Prionics assay (Figures 2J–2L).
Discussion
The diagnosis of animal TSEs is based mainly on the accurate interpretation of lesions such as spongiosis, gliosis, and neuron loss and may be impaired if the brain is not fixed shortly after death. Independently of autolysis, poor-quality fixation and paraffin embedding can also trigger the production of many artifactual vacuoles and retractions of tissues, leading to limited interpretations. Consequently, it is of great interest to promote PrPsc detection, which appears to be more reliable than histopathology as a marker of TSEs (Bolton et al. 1982). Although PrPsc IHC is considered as a highly specific and sensitive tool for PrPsc detection, it is usually performed on fixed materials (Debeer et al. 2001; Miller et al. 1994). The availability of both fixed and frozen halves of the same scrapie-affected sheep brain made it possible to explore its sensitivity when applied to fixed but previously frozen tissue. Indeed, although many antibodies have been characterized using unfixed frozen tissues, only a few, such as antibodies to cytokeratin, have been studied for their reactions on tissue that was frozen and subsequently formalin-fixed and paraffin-embedded (Edgerton et al. 2000).
One of the major constraints analyzed in the present study was the “mechanical” resistance of these fixed-frozen tissues, relative to freshly fixed tissues, to the drastic treatments essential for PrPsc IHC. These treatments were required for both safety and antigen retrieval (Taylor 2000; Taylor et al. 1997). In the present analysis, we showed that all brain pieces retained their integrity during the histological preparation process despite the anticipated cell membrane disruption secondary to freezing and thawing. Similarly, adherence of the brain sections to glass was found to be very satisfactory after all the pretreatments, demonstrating that the physical effects of freezing do not exacerbate this common technical problem in PrPsc IHC. Neither tissue adherence nor integrity was affected by the storage temperature (—20 or — 80C).
We clearly showed that freezing before fixation, compared to freshly-fixed material, resulted in obvious histological damage such as tears following a scarified pattern. It also confirmed the very limited use of that kind of preparation for conventional histology. By contrast, it shows the great value of PrPsc IHC, because PrPsc-positive staining was well preserved in frozen-fixed tissues without generating additional un-specific background staining, as shown by the TSE-negative samples. The finding of an apparent increase of the PrPsc staining in sheep scrapie cases is difficult to interpret as a true enhancement effect of freezing, because in the study there were an insufficient number of comparable cases to indicate whether this would be a frequent or systematic finding. However, it is probably more important to emphasize that PrPsc immuno-labeling patterns, such as those validated in previous studies as specific for TSEs (Miller et al. 1994; Wells et al. 1994; van Keulen et al. 1995; Foster et al. 1996; Debeer et al. 2001; Ryder et al. 2001), were similar to those seen in freshly fixed sections. In fact, a change in those patterns might have been anticipated because freezing leads to cell membrane disruption and, after suspension in a medium during formalin fixation, diffusion of proteins out of cells might have been expected to take place. Surprisingly, this was not observed at all. On the contrary, the cell pattern of PrPsc immunolabeling was either of synapse-like type, as shown by PrPsc deposits surrounding neurons and their processes, or possibly of microglial type, as suggested by a stellate deposition. This was observable even in those BSE cases that were weakly positive using Western blotting. In these, despite the limited number of neurons labeled, the cellular pattern was obviously and strongly PrPsc-positive.
Additional studies may be warranted to test the limits of using that kind of preparation. In particular, we do not know the effect of repeated freezing and thawing nor the effect of long-term freezing. However, the goal of this study was purely to test the applicability of PrPsc IHC on fixed-frozen brain tissue compared to freshly fixed samples. Although the number of cases analyzed in this work was limited, it was clearly demonstrated that PrPsc immunohistochemistry could be applied to fixed-frozen brain samples with very good sensitivity, rendering possible its use in the diagnosis of ruminant TSEs.
Footnotes
Acknowledgements
Supported by grants from European FAIR 98–7021 and French Ministry of Agriculture and Fishery. Sabine Debeer was financially supported by the European FAIR 98–7021.
We thank Dr E. Monks (Veterinary Research Laboratories; Dublin, Ireland) for critical reading of the manuscript and valuable suggestions. We also thank J. Grassi (CEA-Saclay, France) for his courtesy in supplying anti-PrP antibody and the Ventana Company for its technical support with the “NexES” immunostainer.