
Abstract
Taurine exerts a number of actions in mammalian cells, including regulation of ion transport and osmoregulation. The production and secretion of saliva involve transepithelial ion transport, thereby making the plasma-like primary saliva hypotonic before secretion. Therefore, it is plausible to suggest modulation of salivary taurine by muscarinic agents that affect salivary gland function. One of the objectives of this study was to determine tissue content and localization of taurine in the submandibular gland of the rat. Further, we determined whether treatment with muscarinic drugs that either increase (e.g., pilocarpine) or decrease (e.g., propantheline) saliva secretion affects the submandibular gland taurine content. The results indicate that the submandibular gland contains an appreciable amount of taurine (8.9 ± 0.3 moles/g wet wt). Further, acute treatment of the rats with either of the muscarinic drugs did not significantly affect tissue taurine content compared to the control group. By contrast, chronic treatment with propantheline, but not pilocarpine, reduced the tissue content of taurine compared to the control rats (p<0.05). Utilizing light microscopic immunohistochemical techniques, intense immunoreactivity was found primarily in the striated ducts of the submandibular gland. Neither pilocarpine nor propantheline treatment led to differential distribution of immunoreactivity in this tissue. In conclusion, the submandibular gland contains an appreciable amount of taurine, primarily in the striated ducts, that can be decreased by chronic muscarinic receptor blockade.
T
The production and secretion of saliva involve transepithelial transport of ions, thereby regulating saliva tonicity (Bradley 1995; Hedemark-Poulsen 1998). The primary saliva that is produced by the acinar cells is plasma-like with respect to the concentration of sodium, chloride, and bicarbonate. This saliva becomes hypotonic as it flows down the duct system before secretion. The reduction in saliva tonicity is caused primarily by reabsorption of sodium and chloride essentially without further change in water secretion or reabsorption. This process is affected by flow rate which, in turn, is regulated by several mechanisms, including the autonomic nervous system. At low flow rates, the sodium reabsorption mechanism is sufficient for sodium removal so that marked reduction in the sodium concentration of saliva occurs before secretion into the oral cavity. By contrast, at high flow rates, the reabsorption mechanism is insufficient for sodium removal so that an increasing amount of sodium appears in the final saliva (Thaysen et al. 1954; Bradley 1995; Hedemark-Poulsen 1998).
The present investigation was prompted for several reasons. First, taurine is an important player in the adaptive responses of cells to osmotic stress (see Schaffer et al. 2000). This contention is based on reports indicating that exposure of a variety of cells to a hypotonic medium results in cellular extrusion of taurine, a process that contributes to regulatory volume decrease. Conversely, exposure of cells to hypertonic media results in cellular uptake of taurine, an important mechanism in regulatory volume increase (Schaffer et al. 2000). Second, an interaction between sodium and taurine movement is described whereby an increase in intracellular sodium concentration is associated with enhanced taurine efflux from the cell. By contrast, taurine influx is promoted by an increase in extracellular sodium concentration (Suleiman et al. 1992). This relationship has been largely attributed to cotransport of taurine and sodium via the β-amino acid transporter (Wolff and Kinne 1988; Jones et al. 1990,1993; Schaffer et al. 2000). Therefore, given the role of taurine in regulatory volume changes, the association of taurine flux with sodium transport, and the importance of sodium flux in the final composition of saliva, we conjectured that the salivary gland taurine is modulated by autonomic drugs that affect salivary gland function. However, to our knowledge, the tissue content and localization of taurine in salivary glands are not known. Therefore, one objective of our study was to determine tissue taurine content of the submandibular gland of the rat in reference to that of the kidney. The kidney contains large amounts of taurine that is maintained, in part, by the taurine co-transporter (Wolff and Kinne 1988; Jones et al. 1990, 1993). In addition, utilizing immunohistochemical labeling techniques with a polyclonal antibody directed against the taurine molecule, we determined the tissue localization of taurine within the gland. Furthermore, because saliva flow and secretion are stimulated by muscarinic receptor activation but are inhibited by muscarinic receptor blockade (Thaysen et al. 1954; Bradley 1995), we tested the hypothesis that treatment with pilocarpine (muscarinic receptor agonist) and propantheline (muscarinic receptor antagonist) causes differential effects on tissue content and/or localization of taurine.
Materials and Methods
Chronic Treatment with Muscarinic Drugs
Six-week-old male Wistar-Kyoto rats were obtained from Harlan Laboratories (Indianapolis, Indiana). All rats were maintained at constant humidity (60 ± 5%), temperature (24 ± 1C), and light cycle (0600–1800 hr.). Two days after arrival the animals were randomly assigned to three groups: (a) the control group (n=5) was given tapwater, (b) the pilocarpine-treated group (n=5) received tapwater containing pilocarpine (5 mg/kg/day), and (c) the propantheline-treated group (n=5) was provided with tapwater containing propantheline bromide (20 mg/kg/day). To achieve these dosages, the concentration of each drug in the drinking fluid was adjusted on the basis of daily fluid intake and body weight before and after administration of each drug. Food and drinking fluid were available ad libitum throughout the study.
Six days after initiation of drug therapy the animals were anesthetized with sodium pentobarbital (60 mg/kg, IP). The left submandibular gland and the left kidney were removed from each rat, weighed, and homogenized in 2% chilled perchloric acid (1:10 w/v). The homogenates were centrifuged (4C, 10,000 rpm) and the supernatants used for the taurine assay (Shaffer and Kocsis 1981).
Immunohistochemical Labeling
The right submandibular glands from the above animals were placed in formalin solution and used to establish tissue localization of taurine utilizing a light microscopic immunohistochemical technique (Borke et al. 1995; Hsu et al. 1981).
Formalin-fixed submandibular glands used for the localization of taurine were dehydrated through a graded series of ethanols and xylene before embedding in paraffin. Five-μm paraffin-embedded tissue sections were cut and collected on glass slides for immunohistochemical labeling.
The polyclonal antibody AB137, used to localize taurine, was purchased from Chemicon International (Temecula, CA). This antibody was created using a taurine-glutaralde-hyde-polylysine immunogen and shows minimal crossreactivity with other amino acids.
Sections of the submandibular glands were processed for immunohistochemical localization of taurine using the avidin-biotin-peroxidase technique of Hsu et al. (1981), with minor modification. By this technique, the tissue sections on glass slides were deparaffinized with
After immunohistochemical processing, the sections were counterstained with Mayer's hematoxylin (Sigma) to aid in morphological identification. Serial sections were also processed as negative controls without incubation in the primary antibody AB137.
Finally, as an index of the effectiveness of drug treatments, 5-μm paraffin-embedded tissue sections were cut and stained with periodic acid-Schiff (PAS) reagent. The PAS stain was deemed appropriate because the submandibular gland contains both mucous and serous secretory cells and the PAS reagent labels both the mucous secretory component and other secretory granules of the serous component.
Acute Treatment with Muscarinic Drugs
Male Wistar-Kyoto rats (170–190 g) were anesthetized with sodium pentobarbital (60 mg/kg, IP) and implanted with a femoral venous catheter (PE 50 fused to PE 10; Mozaffari and Schaffer 2001). Thereafter, the animals were infused with isotonic saline at the rate of 20 μl/min for 10 min (e.g., stabilization period). This was followed by infusion of either isotonic saline (time-controlled; n = 6), or isotonic saline containing pilocarpine (n = 6) or propantheline (n = 6) for an additional 10 min (20 μl/min). The concentration of each drug in the infusate was adjusted to deliver 50 μg/kg/min of pilocarpine (Nagasako et al. 1993) or 200 μg/kg/min of propantheline. During the stabilization period the right and left submandibular glands (including the sublingual gland) were dissected out and a silk suture placed around the neurovascular bundle of the right salivary glands. Five minutes after administration of either isotonic saline (e.g., time-controlled) or pilocarpine or propantheline, the right submandibular gland was procured after ligating the neurovascular bundle and removing the sublingual gland. The left submandibular gland, on the other hand, was removed 10 min after infusion of each drug or isotonic saline (e.g., time-controlled). Each submandibular gland was weighed and immediately homogenized in 2% chilled perchloric acid (1:10 w/v). The homogenates were centrifuged (4C, 10,000 rpm) and the supernatants used for the taurine assay (Shaffer and Kocsis 1981).
Statistics
All quantitative comparisons between groups were analyzed by ANOVA and Duncan's test was employed for comparison of mean values (significance of criteria of p<0.05). Data are reported as means ± SEM.
Results
Chronic Treatment with Muscarinic Drugs
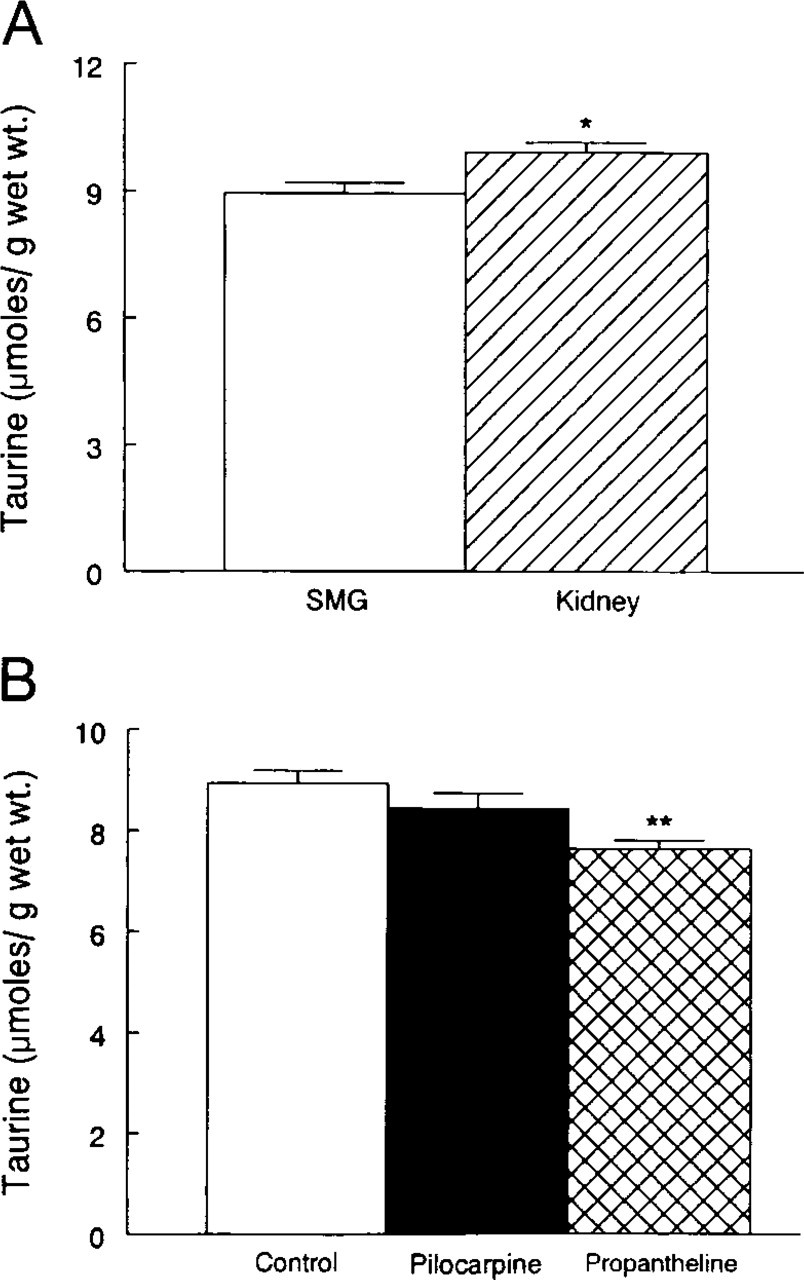
Body weight was similar among the groups before initiation of drug therapy and was not affected by either pilocarpine or propantheline treatment [173 ± 5 g (control), 171 ± 3 g (pilocarpine-treated), and 171 ± 2 (propantheline-treated)]. Furthermore, no differences were noted among the groups in tissue weights of the left submandibular gland [197 ± 6 mg (control), 200 ± 11 mg (pilocarpine-treated), and 190 ± 9 mg (propantheline-treated)] and the left kidney [698 ± 14 mg (control), 704 ± 26 mg (pilocarpine-treated), and 695 ± 16 mg (propantheline-treated)].
Basal taurine content of the submandibular gland was slightly lower than that of the kidney (Figure 1A; p<0.05). Whereas treatment of the rats with pilocarpine did not affect the submandibular gland content of taurine, treatment with propantheline caused a significant decrease (∼15%) compared to the control group (Figure 1B).
(
Light microscopic examination revealed significant immunoreactivity with taurine-specific antibody. In sections of the submandibular gland from the control group (Figure 2A and 2B), localization was found mainly in the blood vessels and striated ducts, whereas the acini showed no immunoreactivity. Chronic treatment with either pilocarpine (Figure 2D and 2E) or propantheline (Figure 2G and 2H) did not alter the pattern of localization of immunoreactivity. Immunohistochemical processing of the tissue without the use of the primary anti-taurine antibody showed an absence of staining [Figures 2C (control), 2F (pilocarpine-treated), and 2I (propantheline-treated)].
To illustrate the effectiveness of the muscarinic drug regimens, tissue sections from the control and drug-treated rats were stained with PAS (Figures 2J-2L). The staining intensity was reduced in the submandibular gland sections of the pilocarpine-treated rats compared to the control rats (Figure 2K vs 2J). By comparison, tissue sections from the propantheline-treated rats (Figure 2L) displayed intense staining compared to the tissue sections from either the control or the pilocarpine-treated rats (Figures 2J and 2K, respectively). As a result, the difference in staining was most prominent between the propantheline-treated and the pilocarpine-treated rats (Figure 2L vs 2K). Muscarinic receptor stimulation (e.g., pilocarpine) increases but muscarinic receptor blockade (e.g., propantheline) reduces salivary gland secretion (Thaysen et al. 1954; Bradley 1995). Therefore, the difference in staining intensity is likely due to the reduction and accumulation of salivary secretory products with pilocarpine and propantheline, respectively.
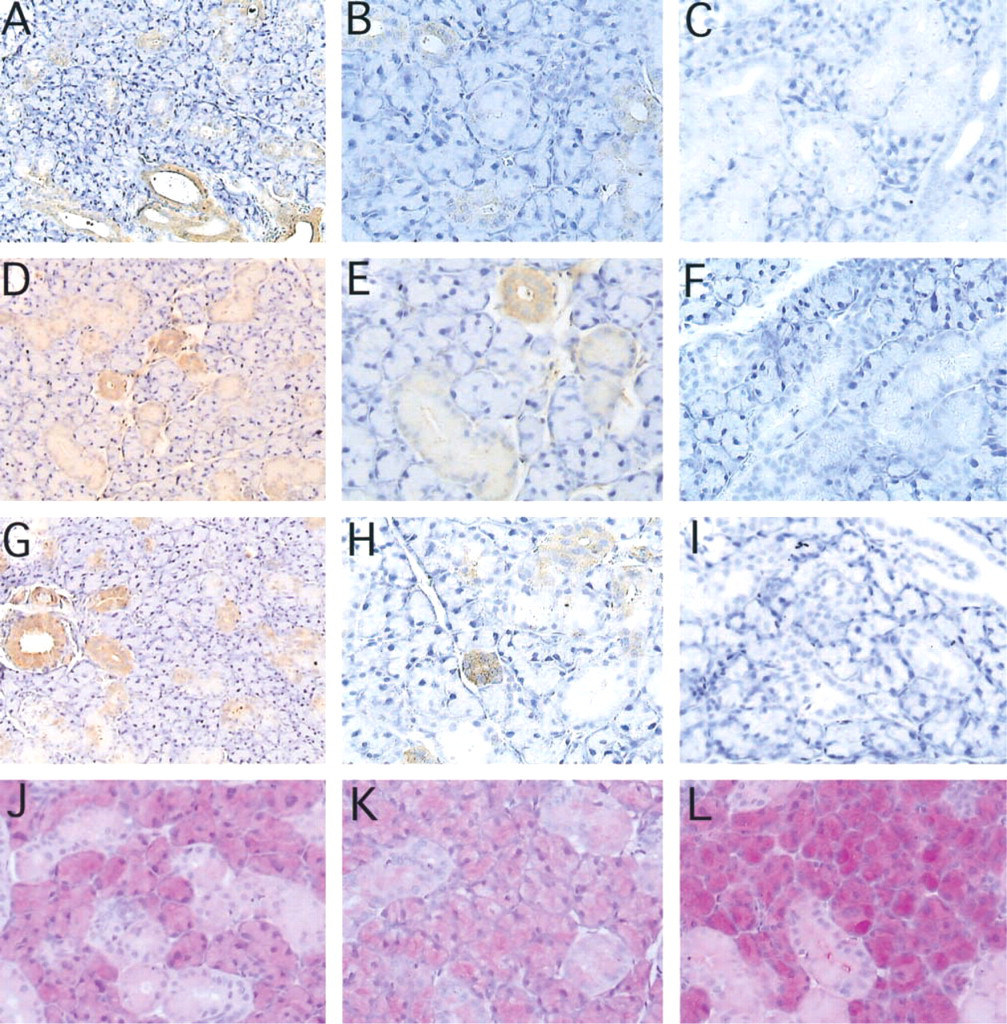
Light microscopic examination reveals intense immunoreactivity with taurine-specific antibody in the striated ducts and blood vessels of the submandibular gland in the vehicle-treated control rat [
Acute Treatment with Muscarinic Drugs
Acute IV administration of either pilocarpine or propantheline to rats for either 5 (right gland) or 10 (left gland) min did not significantly affect tissue taurine content in the pilocarpine-treated [9.6 ± 0.4 (right gland) vs 9.7 ± 0.6 (left gland) μmoles/g wet wt] or propantheline-treated [9.0 ± 0.3 (right gland) vs 8.6 ± 0.1 (left gland) μmoles/g wet wt] rats compared to the control group [8.9 ± 0.4 (right) vs 8.8 ± 0.3 (left) μmoles/g wet wt]. Interestingly, however, the submandibular gland taurine content tended to be lower in the propantheline-treated rats compared to the pilocarpine-treated rats 10 min after initiation of drug administration (8.6 ± 0.1 vs 9.7 ± 0.6 μmoles/g wet wt).
Discussion
The effects of taurine on cell function have received considerable attention in many tissues such as the kidney, heart, and brain (Wolff and Kinne 1988; Jones et al. 1990,1993; Suleiman et al. 1992; Trachtman et al. 1993; Nakanishi et al. 1994; Saransaari and Oja 2000; Schaffer et al. 2000; Mozaffari and Schaffer 2001). To our knowledge, the present investigation is the first report demonstrating that the rat submandibular gland contains an appreciable amount of taurine, suggestive of its potential involvement in salivary gland function because of the following observations. First, the submandibular gland content of taurine is closely similar to that of the kidney, an organ intimately involved in the regulation of whole-body taurine homeostasis and in which taurine has been shown to contribute to cell volume regulation and modulation of renal excretory function (Jones et al. 1990,1993; Nakanishi et al. 1994; Mozaffari and Schaffer 2001). Second, taurine is preferentially located in the striated ducts of the submandibular gland, a region that contributes importantly to regulation of the ionic composition of saliva (Bundgaard et al. 1977). However, the significance of this finding still remains to be established. Similarly, taurine is preferentially localized in the medullary region of the kidney and is believed to play a role not only in protecting the renal tubule cells from osmotic stress but also in exerting a renoprotective effect in animal models of renal failure (Wolff and Kinne 1988; Trachtman et al. 1993; Nakanishi et al. 1994). Third, chronic but not acute treatment of rats with the muscarinic receptor antagonist decreased submandibular gland taurine content compared to the untreated control rats.
The lack of a significant effect of acute exposure to either of the muscarinic drugs corroborates previous reports indicating that significant alterations in the tissue taurine pool occur after chronic but not acute experimental manipulations (Atlas et al. 1984; Schaffer et al. 2000). This may relate, in part, to the slow rate of taurine uptake by the taurine co-transporter as well as transepithelial movement (e.g., influx and efflux) of taurine (Benyajati and Bay 1994; Schaffer et al. 2000). Therefore, the involvement of the taurine co-transporter in modulation of ionic composition and tonicity of saliva after acute manipulation of salivary gland function cannot be ruled out at this time. This contention is partially supported by a tendency for reduction in the submandibular gland taurine content 10 min after IV administration of propantheline compared to pilocarpine.
The reason for the chronic propantheline-induced reduction in submandibular gland taurine is not apparent from this study. One plausible scenario relates to chronic blockade of the muscarinic receptors and the attendant reduction in salivary flow rate, which would promote the formation of a more hypotonic saliva (see Bradley 1995). Taurine efflux is a prominent component of cellular adaptation to a hypotonic environment (see Schaffer et al. 2000). As a result, the duct cells from propantheline-treated rats would be expected to extrude taurine in order to cope with chronic exposure to a more hypotonic ductal fluid. This effect would be much less in pilocarpine-treated rats because the attendant increase in salivary flow rate reduces passage time and subsequently the reabsorption of sodium and chloride. Therefore, increasing amounts of these ions would remain in the final saliva. Thus, the postulated tonicity-mediated taurine efflux from the duct cells may manifest itself primarily when saliva flow rate is reduced (e.g., propantheline treatment) but much less when saliva flow rate is enhanced (e.g., pilocarpine treatment).
The modification of the plasma-like primary saliva occurs via ductal reabsorption of Na+ and Cl−, essentially without water, thereby making the final saliva hypotonic. Several transport processes operate in concert to determine the ionic composition of saliva. These include the basolaterally located Na+-K+-ATP-ase, Cl− and K+ channels, and the Na+/H+ exchanger, together with luminally located Na+ and Cl− channels, K+/H+, Na+/H+, and the Cl−/HCO3 − exchangers (see Hedemark-Poulsen 1998). Because taurine transport is linked to sodium and chloride transport (2–3 Na+; 1 Cl−; 1 taurine) (Wolff and Kinne 1988; Jones et al. 1990,1993), it is likely that the taurine co-transporter participates in determining the final ionic composition of saliva. Although the presence of the taurine co-transporter on the luminal and/or basolateral membrane of the salivary gland duct remains to be established, the following observations point to that possibility. First, after IP administration of [35S]-taurine, the salivary glands accumulate taurine (Shimada et al. 1984). Second, we have demonstrated reduction of approximately 45% in the taurine content of the rat submandibular gland after 3 weeks of treatment with β-alanine (unpublished data). β-Alanine blocks cellular taurine uptake, thereby causing taurine depletion (Shaffer and Kocsis 1981; Jones et al. 1990). In conclusion, the demonstration of an appreciable amount of taurine and its preferential localization in the striated ducts of the submandibular gland, as well as modulation of tissue taurine content by chronic muscarinic receptor blockade, suggest a role for taurine in salivary gland function. In light of these observations, it is plausible to postulate a dual role for taurine in salivary gland function, i.e., regulation of duct cell volume as well as determination of ionic composition and tonicity of saliva. In this context, the potential contributions of taurine efflux vs taurine influx to these processes may represent a fertile area for future investigation.
Footnotes
Acknowledgements
Supported by a grant from Taisho Pharmaceutical Company of Japan.
We are grateful to Drs Worku Abebe and Rafik Abdel-sayed (Department of Oral Biology and Maxillofacial Pathology, MCG School of Dentistry) for their critical review of the manuscript. We also thank Champa Patel and Anita Smith for technical assistance.