
Abstract
The alkaline phosphatases are a small family of isozymes. Bovine preattachment embryos transcribe mRNA for two tissue-specific alkaline phosphatases (TSAP2 and TSAP3) beginning at the 4- and 8-cell stages. Whereas no mRNA has been detected in oocytes, there is maternally inherited alkaline phosphatase activity. It is not known which isozyme(s) is responsible for the maternal activity or when TSAP2 and TSAP3 form functional protein. No antibodies are available that recognize the relevant bovine alkaline phosphatases. Therefore, sensitivity to heat and chemical inhibition was used to separate the different isozymes. By screening tissues, it was determined that the bovine tissue-nonspecific alkaline phosphatase (TNAP) is inactivated by low temperatures (65C) and low concentrations of levamisole (<1 mM), whereas bovine tissue-specific isozymes require higher temperatures (90C) and levamisole concentrations (>5 mM). Inhibition by L-homoarginine and L-phenylalanine was less informative. Cumulus cells transcribe two isozymes and the pattern of inhibition suggested heterodimer formation. Inhibition of alkaline phosphatase in bovine embryos before the 8-cell stage indicated the presence of only TNAP. At the 16-cell stage the pattern was consistent with TNAP plus TSAP2 or −3 activity, and in morulae and blastocysts the pattern indicated that the maternal TNAP is fully supplanted by TSAP2 or TSAP3.
The Alkaline Phosphatases are a small family of isozymes expressed in diverse species, including bacteria, mammals, reptiles, amphibians, nematodes, and insects (McComb et al. 1979; Millan 1988,1990). Conservation of alkaline phosphatase expression hints at an important role, but to date there is little biochemical evidence for alkaline phosphatase function except in bone mineralization (Yoon et al. 1989; Beertsen and van den Bos 1992; Fedde et al. 1999) and de-phosphorylation of adenosine nucleotides (Gallo et al. 1997). The alkaline phosphatases have been postulated to be involved in a range of other processes, including cell adhesion (Hui et al. 1993), vitamin B transport (Rindi et al. 1995), metastasis (Manara et al. 2000), and cell signaling (Swarup et al. 1981; Muller et al. 1991). We are interested in their role in the development of mammalian preimplantation embryos.
In mammals, the alkaline phosphatase family consists of two groups, tissue-nonspecific alkaline phosphatase and the tissue-specific alkaline phosphatases. The single tissue-nonspecific alkaline phosphatase (TNAP) gene is expressed in many tissues, including liver, bone, and kidney (Garattini et al. 1987; Hsu et al. 1987; Harris 1989; Hahnel et al. 1990; MacGregor et al. 1995; Hoshi et al. 1997; McDougall et al. 1998). Humans with mutations in the TNAP isozyme have defective bone mineralization (infantile hypophosphatasia; Henthorn et al. 1992), and mice engineered to lack TNAP have poor bone mineralization plus a range of defects that result in postnatal death (Narisawa et al. 1997).
The number of tissue-specific alkaline phosphatases expressed depends on the species. There are two tissue-specific isozymes in mice (Manes et al. 1990): intestinal alkaline phosphatase and embryonic alkaline phosphatase (EAP). The EAP isozyme is the major isozyme expressed by 2-cell to blastocyst stage embryos and M-phase spermatogenic cells in the testis (Hahnel et al. 1990; Manes et al. 1990; Narisawa et al. 1992; MacGregor et al. 1995). Mice homozygous null for the mouse EAP gene (EAP knockout) are viable (Narisawa et al. 1997). However, we found that its absence disadvantages preimplantation embryos (Dehghani et al. 2000). Fewer homozygous null embryos survive than wild-type, particularly under even mild stress, such as culture. The results suggest that, in mice, EAP is part of an essential process in blastocyst formation, although its role appears to be duplicated by another molecule. Whether alkaline phosphatase plays a similar role in preimplantation development in other mammals remains to be determined. There are three human tissue-specific alkaline phosphatases: intestinal, germ cell, and placental (Harris 1989). Ectopic expression of alkaline phosphatases is often found in cancer cells (Millan 1988,1992; Millan and Manes 1988). It is not known whether human preimplantation embryos express a tissue-specific alkaline phosphatase. In cattle, several different cDNA sequences have been cloned from intestine (Weissig et al. 1993; Manes et al. 1998), and we have cloned two tissue-specific cDNAs (TSAP2 and TSAP3) from blastocysts (McDougall et al. 1998). Transcription of TSAP2 is first detected at the 4-cell stage in the in vitro-derived bovine embryo and TSAP3 at the 8-cell stage. This is equivalent to the mouse 2-cell stage relative to the initial burst of transcription from the embryonic genome (Telford et al. 1990). Therefore, one or both of these bovine isozymes may be the functional equivalent of the mouse EAP. TSAP2 also is transcribed in testis, spleen, and cumulus cells, and TSAP3 is transcribed by kidney and oviduct epithelial cells (McDougall et al. 1998). A complication in the comparison to mouse is that alkaline phosphatase activity is detected in all stages from oocyte to blastocyst, although transcription of alkaline phosphatase does not begin until the late 4-cell stage (McDougall et al. 1998). It is assumed that the early activity is due to maternal protein from mRNA transcribed during oogenesis. Which isozyme is responsible for the maternal activity is unknown. This research was designed to determine the isozyme(s) responsible for the protein activity encountered in preattachment bovine embryos at the various stages.
Antibodies are currently available for only bovine intestinal alkaline phosphatase. In other species, however, TNAP and the tissue-specific isozymes have different sensitivities to temperature and chemical inhibition by various L-form amino acids and levamisole. Both mouse and human TNAP are particularly sensitive to inhibition with levamisole and L-homoarginine, and are comparatively insensitive to L-phenylalanine (Harris 1989; Lepire and Ziomek 1989; Millan 1990). The tissue-specific isozymes are comparatively insensitive to levamisole and L-homoarginine and sensitive to L-phenylalanine inhibition (Harris 1989; Lepire and Ziomek 1989; Millan 1990). The human and murine TNAPs are inactivated by temperatures less than 60C, whereas the tissue-specific isozymes typically withstand higher temperatures (up to 90C; Harris 1989; Lepire and Ziomek 1989). Therefore, we began this study with examination of the response of bovine tissues transcribing various alkaline phosphatase isozymes to heat and chemical inhibition and subsequently used the information to assay the AP isozymes expressed in preattachment bovine embryos. Because of the difficulty of obtaining sufficient material for biochemical tests of alkaline phosphatase from preattachment bovine embryos, a histochemical assay was used throughout.
Materials and Methods
Tissue Preparation
Adult bovine tissues (liver, kidney, intestine, spleen, testis) were recovered at slaughter, flash-frozen in OCT embedding compound (Immucore Canada; Edmonton, Alberta, Canada) in liquid nitrogen, and stored at — 80C. Sections (7 μm) were cut using a cryostat, mounted on slides, fixed for 20 min with 4% paraformaldehyde (Sigma; St Louis, MO), washed three times in PBS, and held briefly in PBS before treatment.
Cumulus cells and bovine oviduct epithelial cells were prepared during the culture of embryos. Cumulus cells were recovered during the stripping of presumptive zygotes by gentle pipetting (see below), washed three times in PBS containing 0.1% polyvinyl pyrolidone (PBS+PVP; Fisher Scientific, Unionville, Ontario, Canada), and the cells were smeared on slides. Bovine oviduct epithelial cells were recovered from co-culture preparation (see below), washed three times in PBS + PVP, and smeared on slides. Cells were allowed to dry overnight, then fixed for 20 min in 4% paraformaldehyde and held in PBS until treatment.
Embryo Preparation
Embryos were produced by in vitro maturation, fertilization, and culture as described previously (Xu et al. 1992), with the following modifications. Cumulus-oocyte complexes were aspirated from visible follicles less than 1 cm in diameter using an 18-gauge needle into modified Ham's F-10 medium (Gibco BRL; Burlington, Ontario, Canada) containing 10 mM HEPES (Gibco BRL), 2 U/ml heparin (Hepalean; Organon Teknica, Toronto, Ontario, Canada), and 2% heat-inactivated steer serum. They were matured for 22–24 hr in TCM-199 medium (HEPES buffered medium 199 with Earle's salts; Gibco BRL) containing 55 mg/ml sodium pyruvate and 29 μg/ml L-glutamine (Sigma) and 1.2% heat-inactivated steer serum. After maturation, oocytes were washed twice in sperm-TALP (Greve et al. 1987) buffered with 10 mM HEPES (IVF medium) and placed in this medium for fertilization. Sperm were prepared by thawing two 0.5-ml straws at 38C for 1 hr in IVF medium, subjecting to swim-up for 1 hr in IVF medium, and pelleting by centrifugation. Sperm were added to the matured oocytes at a concentration of 1.0 × 106 sperm per ml, and incubated for 18 hr at 38C in 5% CO2 in humidified air. Presumptive zygotes were stripped of cumulus by gentle vortexing and co-cultured with oviduct epithelial cells in Medium 199 with Earle's salts to which was added 55 mg/ml pyruvate, 29 μg/ml glutamine, 0.35% bovine serum albumin (fraction V; Sigma) and 1.2% steer serum (IVC medium). Embryos were removed from culture at specific stages of development (1-cell, 2-cell, 4-cell, 8-cell, 16-cell, morula, and blastocyst) to assay for AP activity. They were rinsed three times in PBS+PVP, fixed in 4% paraformaldehyde for 1 hr, washed three times in PBS+PVP, and held in PBS+PVP.
Bovine oviduct epithelial cells for co-culture were prepared as follows. Oviducts were trimmed and rinsed and the epithelium stripped by squeezing the oviduct with fine forceps, forcing sheets of epithelial cells from the oviduct into IVC medium. Large sheets of cells were broken by aspiration through a 25-gauge needle and cultured in 1.5 ml IVC medium for 24 hr. In vitro culture medium for embryos (see above) was conditioned for 24 hr by the addition of the oviduct epithelial cells to culture wells on the day of fertilization, 18–24 hr before adding embryos (Xu et al. 1992).
Heat Inactivation
There were three independent trials of tissues and embryos. Each trial used tissue from a different individual or a different batch of in vitro-produced bovine embryos. Tissue sections were incubated at temperatures ranging from 25C to 100C (in 5C increments) for 1 hr in PBS in an oven (for temperatures less than 80C) or in a water bath (for temperatures over 80C). Sections were removed after 1 hr and rinsed in PBS. Tissue sections were equilibrated for 10 min in 0.1 M Tris, pH 10.0 (Sigma). They were then incubated for 20 min in the same buffer containing NBT/BCIP substrate [200 μl substrate per 10 ml 0.1 Tris, pH 10.0, NBT (nitroblue tetrazolium chloride), BCIP (5-bromo-4-chloro-3-indolyl phosphate; Boehringer Mannheim, Montreal, Quebec, Canada)] in the dark at room temperature. Sections were rinsed three times with PBS, counterstained for 5 sec with methyl green (0.1% w/v solution; Allied Chemical, New York, NY), rinsed another three times in PBS, and coverslips mounted with Aqua-Polymount (Polyscience; Warrington, PA).
Using bovine tissue sections, incubation at 70C was found to distinguish TNAP from tissue-specific alkaline phosphatase activities, so embryos were incubated for 1 hr at 70C in PBS + PVP in capped 200 μl Eppendorf microcentrifuge tubes (to prevent loss of volume due to evaporation). Embryos were recovered from tubes, equilibrated for 10 min in 0.1 M Tris, pH 10.0, and subjected to the AP protein activity assay as previously described, except that embryos were not counterstained with methyl green. Embryos were mounted in PBS under coverslips sealed with petroleum jelly to prevent evaporation and were scored immediately.
Chemical Inactivation
As for heat inactivation, tissues from different individuals or different batches of embryos were used in each of three trials. Inhibitors were prepared as 500 mM stock solutions in PBS, then further diluted to working solutions in 0.1 M Tris, pH 10.0, directly before use. Working inhibitor concentrations were 50 mM, 25 mM, 10 mM, 5 mM, and 1 mM for levamisole (Sigma), L-phenylalanine (Sigma), and L-homoarginine (ICN Biomedicals; Costa Mesa, CA). Tissue sections on slides were incubated for 1 hr in 0.1 M Tris, pH 10.0, containing an inhibitor. Alkaline phosphatase enzyme activity was then determined by incubation for 20 min in the same inhibitor as the previous incubation, containing NBT/BCIP substrate as above, at room temperature in the dark. Sections were rinsed three times in PBS and coverslips were mounted with Aqua-Polymount and observed under a light microscope.
Results from adult bovine tissues showed that incubation in 50, 25, or 10 mM L-phenylalanine and L-homoarginine, and 10, 5, or 1 mM levamisole, in 0.1 M Tris, pH 10.0, for 1 hr would provide a characteristic pattern of alkaline phosphatase inactivation. Embryos were incubated in inhibitor in 0.1 M Tris, pH 10.0, for 1 hr, then for 20 min in 0.1 M Tris, pH 10.0, plus inhibitor containing NBT/BCIP substrate as described above. Embryos were washed three times in PBS + PVP, mounted, and scored as above.
Results
Heat Inactivation
Bovine tissues known to transcribe only tissue-specific alkaline phosphatase mRNA (McDougall et al. 1998) were found to require a higher temperature (90C) to fully eradicate alkaline phosphatase activity than bovine liver (65C), which transcribes only tissue-nonspecific alkaline phosphatase (Table 1). Three trials gave identical results. Figure 1 shows examples of intestinal tissue incubated at room temperature, 80C, and 90C. Bovine spleen, which transcribes mRNA for TSAP2 and TSAP3, also required incubation for 1 hr at 90C to fully eradicate activity (Figures 1I-1J). Kidney, bovine oviduct epithelial cells, and cumulus cells transcribe both TNAP and a tissue-specific isozyme, as determined by RT-PCR (McDougall et al. 1998), and therefore could, in theory, express a combination of alkaline phosphatase proteins. These tissues required a temperature similar to that of liver TNAP for inactivation (Table 1). As shown for kidney in Figures 1E-1G, all positive cells were inactivated at the same temperature (60C); there were no cells with residual thermotolerant activity.
Inhibitor concentrations (mM) and temperature (C) required to inactivate alkaline phosphatase (AP) enzyme activity in adult bovine tissues with a 1 hr incubationa
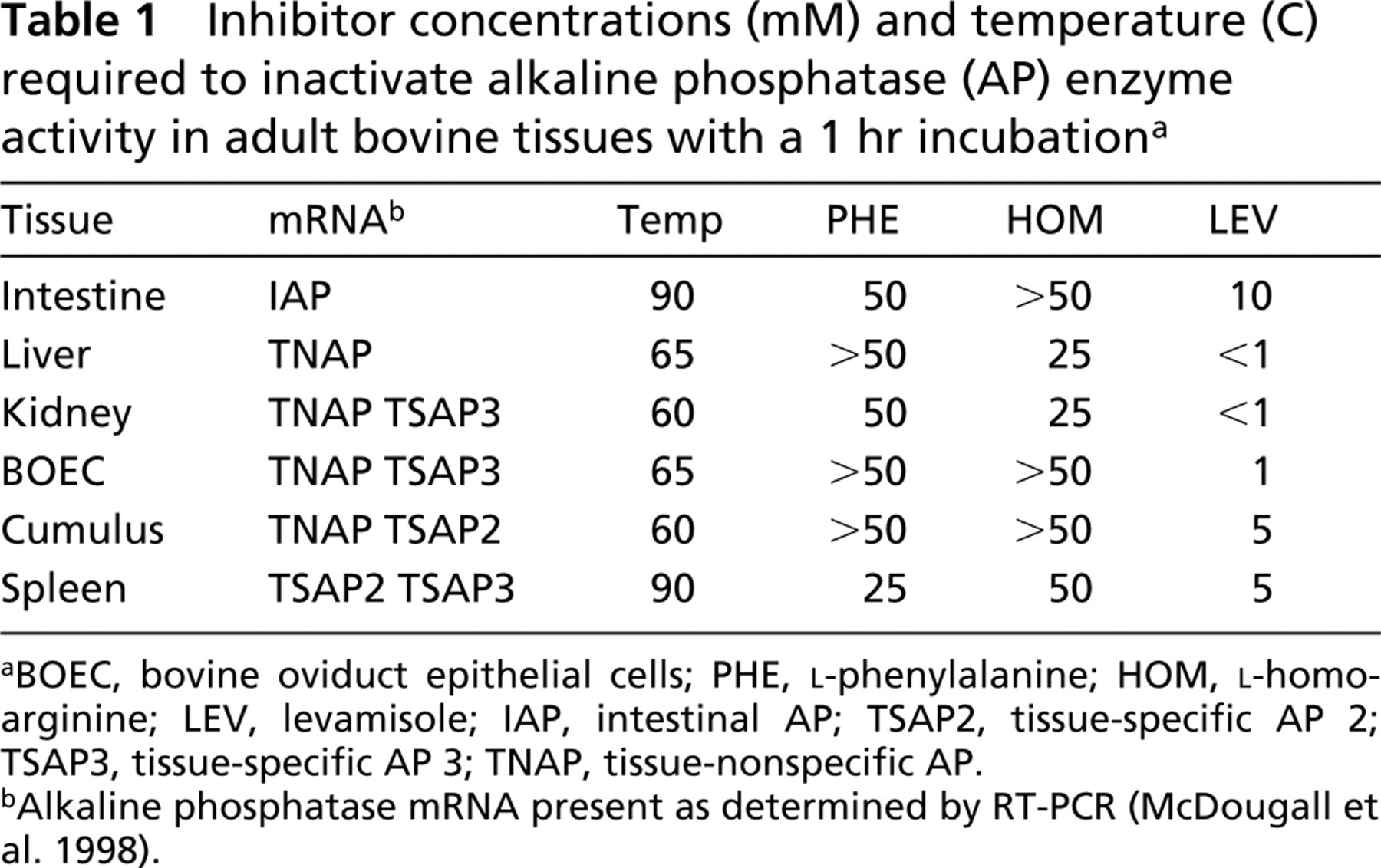
BOEC, bovine oviduct epithelial cells; PHE, L-phenylalanine; HOM, L-homoarginine; LEV, levamisole; IAP, intestinal AP; TSAP2, tissue-specific AP 2; TSAP3, tissue-specific AP 3; TNAP, tissue-nonspecific AP.
Alkaline phosphatase mRNA present as determined by RT-PCR (McDougall et al. 1998).
Inhibitor concentrations (mM) and temperature (C) that inactivate alkaline phosphatase activity in preattachment bovine embryosa
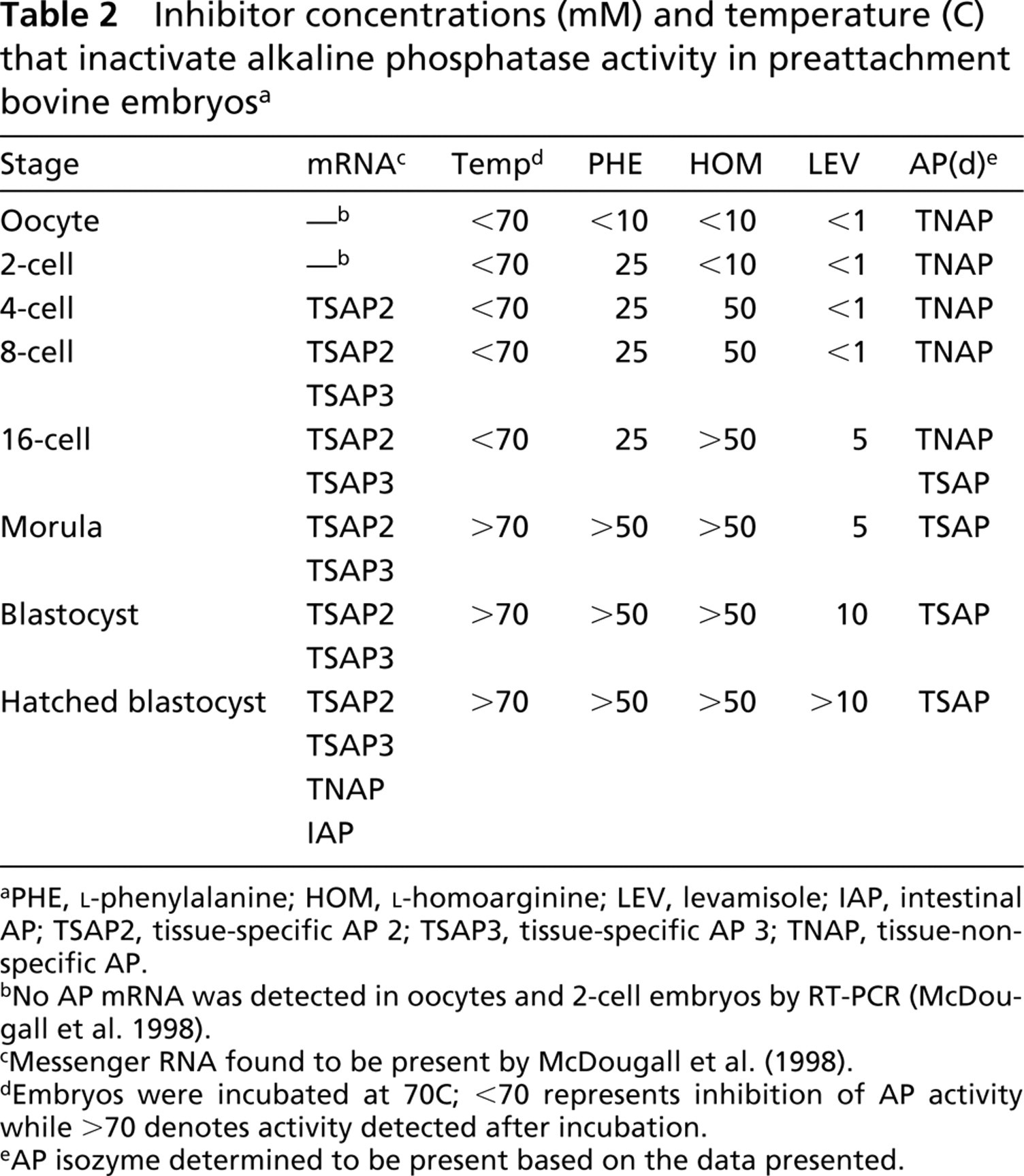
PHE, L-phenylalanine; HOM, L-homoarginine; LEV, levamisole; IAP, intestinal AP; TSAP2, tissue-specific AP 2; TSAP3, tissue-specific AP 3; TNAP, tissue-nonspecific AP.
No AP mRNA was detected in oocytes and 2-cell embryos by RT-PCR (McDougall et al. 1998).
Messenger RNA found to be present by McDougall et al. (1998).
Embryos were incubated at 70C; <70 represents inhibition of AP activity while >70 denotes activity detected after incubation.
AP isozyme determined to be present based on the data presented.
Embryos were incubated at 70C for 1 hr, because we found that this temperature distinguished TNAP, or a combination of isozymes containing TNAP, from the tissue-specific isozymes in adult tissues. Oocytes and embryos, up to and including the 16-cell stage, were negative for alkaline phosphatase activity when incubated for 1 hr at 70C. Morulae, blastocysts, and hatched blastocysts still were positive after 1 hr of incubation (Table 2), showing a change from TNAP to TSAP2 or 3-expression after the 16-cell stage. Figures 2A, 2B, 2E, and 2F compare alkaline phosphatase activity of 8-cell embryos and early blastocysts incubated at room temperature and 70C.
Chemical Inactivation
Table 1 summarizes the concentration of inhibitor that eliminated bovine alkaline phosphatase activity in adult tissues. Again, this occurred at the same incremental step for each tissue in all three trials. Figures 1D and 1H show examples of intestine and kidney incubated with 10 mM and 1 mM levamisole, respectively, and Figures 1K and 1L compare spleen incubated in 10 mM and 25 mM L-homoarginine. Levamisole is a potent bovine TNAP inhibitor, effectively blocking all alkaline phosphatase activity in liver at <1 mM, whereas levamisole did not block the intestinal activity until 10 mM and spleen activity (TSAP2 or −3) until 5 mM. Kidney and bovine oviduct epithelial cells, which transcribe a combination of TNAP and TSAP3, require ≤1 mM LEV, similar to liver TNAP, but cumulus cells, which contain message for TNAP + TSAP2, required 5 mM levamisole for inactivation, like spleen. Bovine liver TNAP was inactivated by 25 mM L-homoarginine, bovine intestinal alkaline phosphatase by >50 mM, and bovine spleen by 50 mM. But kidney again behaved as TNAP, while bovine oviduct epithelial cells and cumulus cells required concentrations like those of the intestinal isozyme (>50 mM L-homoarginine). Liver required >50 mM L-phenylalanine for inactivation, while intestinal tissue required 50 mM and spleen 25 mM. Again, kidney and oviduct epithelial cells did not match, although containing mRNA for the same isozymes, with kidney requiring 50 mM and the oviduct cells needing >50 mM. Cumulus cells also required >50 mM L-phenylalanine for inactivation.
In preattachment embryos, the inactivation of alkaline phosphatase by chemical inhibitors changed with each stage examined (Table 2). Examples of early blastocysts and 8-cell embryos incubated in 5 mM levamisole and 50 mM L-phenylalanine are shown in Figures 2C, 2D, 2G, and 2H. Oocytes, 2–, 4–, and 8-cell embryos required <1 mM levamisole to inhibit alkaline phosphatase activity. Sixteen-cell through hatched blastocysts required >1 mM levamisole, 5 mM for 16-cell and morula, 10 mM for blastocysts, and >10 mM for hatched blastocysts. Oocytes and 2-cell embryos required <10 mM L-homoarginine, while 4-and 8-cell embryos required 50 mM and 16-cell embryos required >50 mM for inactivation of alkaline phosphatase activity. Oocyte activity was inhibited by <10 mM L-phenylalanine, 2-cell through 16-cell embryos by 25 mM, morula and all blastocysts by >50 mM. Again, there was no evidence of two-step inactivation, even in embryos known to have mRNA for two isozymes, and the activity on all cells seemed equally sensitive within an embryo.
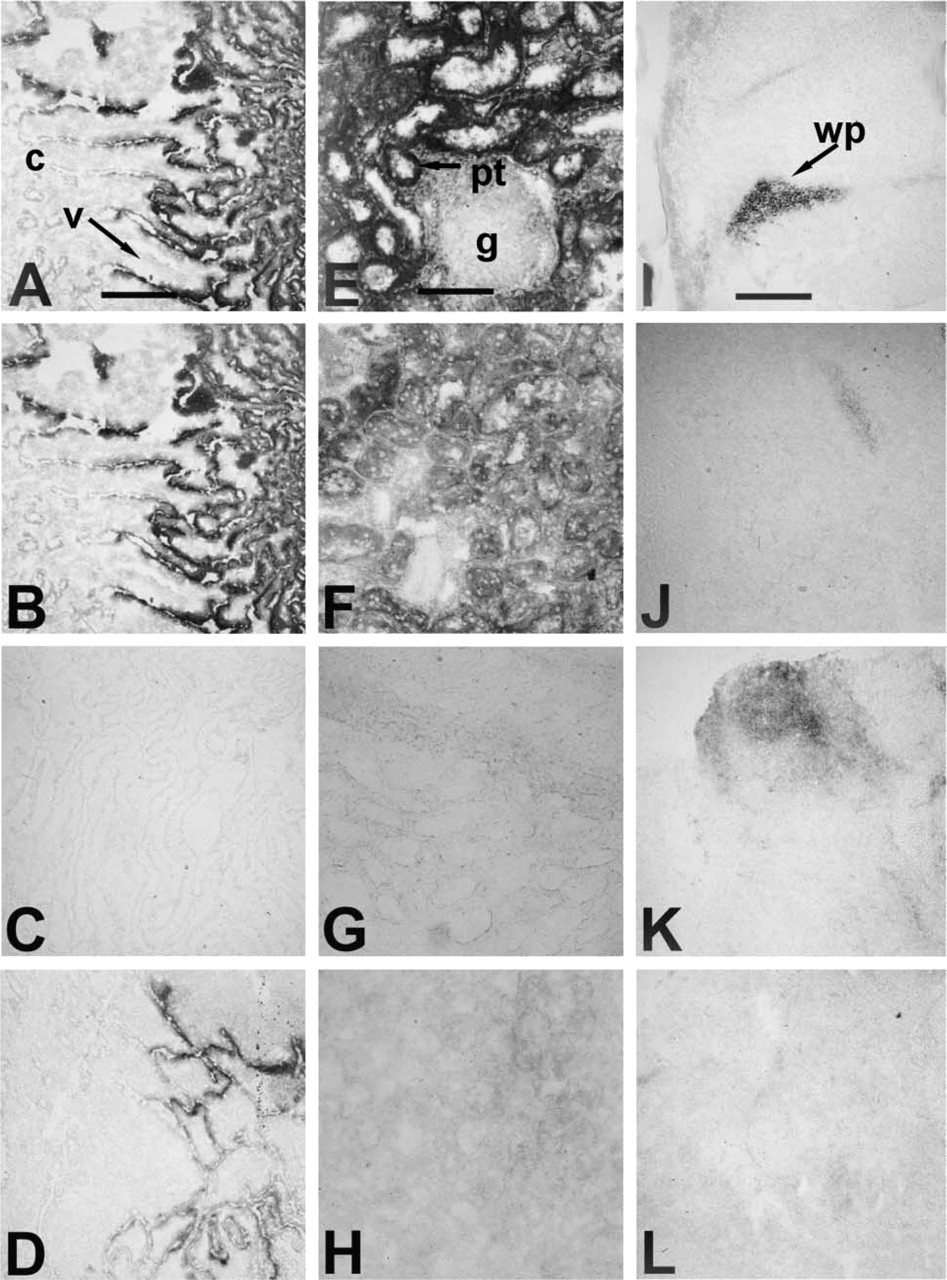
Effects of heat and chemical inhibition on alkaline phosphatase enzyme activity. Cryosections of adult bovine intestine (
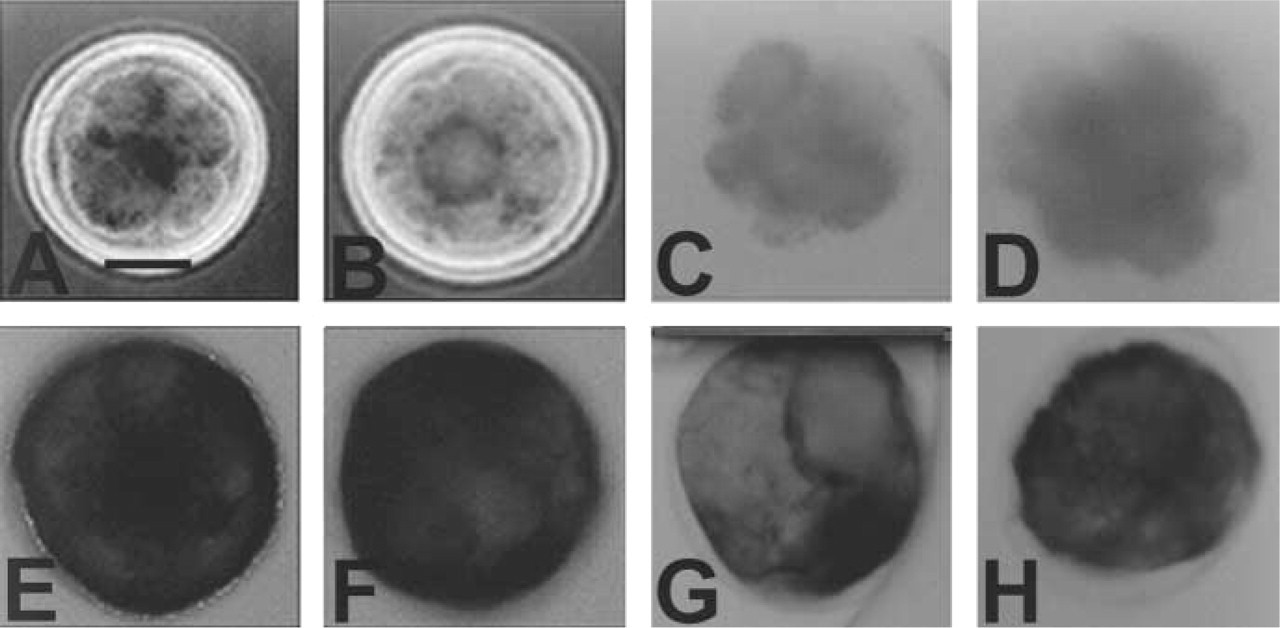
Effects of heat and chemical inhibition on alkaline phosphatase enzyme activity in preattachment bovine embryos. Whole-mount preparations of 8-cell embryos (
Discussion
Inhibition of alkaline phosphatase activity by temperature and levamisole provided the clearest distinctions between bovine isozymes in this histochemical analysis. Adult bovine tissues transcribing only a tissue-specific or combination of tissue-specific mRNA require higher temperatures and higher levamisole concentrations to inhibit alkaline phosphatase activity than tissues expressing TNAP alone (see intestine and spleen vs liver in Table 1). This corresponds with data from mice reported by Lepire and Ziomek (1989) and with data from humans reported by Harris (1989).
The alkaline phosphatases usually are described as homodimers. In mouse and humans there is expression of different isozymes (homodimers) by different cell types, e.g., in the testis (Narisawa et al. 1992). However, formation of functional heterodimers has been found in human cell lines (Wray and Harris 1982; Kodama et al. 1994) and in neutrophils of individuals with Down's syndrome (Vergnes et al. 1999). In bovine preattachment embryos there is simultaneous transcription of TSAP2 and TSAP3 by what appears to be a homogeneous population of embryonic cells. Therefore, it is interesting to compare the bovine tissues transcribing a combination of isozymes to tissues transcribing a single class of alkaline phosphatase. The results suggest that TNAP/TSAP2 heterodimers may be formed in cumulus cells. If there were two distinct homodimer populations in cumulus cells, it would be expected that TNAP would be inactivated before TSAP2 with heat and levamisole. This would result in a loss of activity from some cells or a partial loss of activity in all cells at approximately 60C and <1 mM levamisole, with a further loss at 90C and ≥5 mM levamisole. This was not observed. If only one isozyme formed functional protein in cumulus cells (although two are transcribed), then the inhibition profile would probably resemble that of either TSAP2 or TNAP alone. Heat inactivation of cumulus cells was like TNAP, and the levamisole inhibition like that of a tissue-specific alkaline phosphatase. One explanation for the response to the different inhibitors is suggested by the report that functions of human placental alkaline phosphatase subunits are only partly independent of each other (Hoylaerts et al. 1997). In contrast, bovine kidney and oviduct epithelial cells transcribe TNAP and TSAP3. Neither tissue had an inhibition profile that was clearly distinct from liver with TNAP alone. It is possible that TSAP3 is not translated or that it behaves differently from TSAP2 in combination with TNAP. Unfortunately, bovine TSAP2 and TSAP3 messages were not detected alone in a tissue, and therefore their individual temperature and levamisole sensitivities could not be determined.
Inhibition of bovine intestinal alkaline phosphatase and TNAP with L-homoarginine and L-phenylalanine was similar to what has been described for the mouse isozymes (Lepire and Ziomek 1989) and human (Fish-man 1974; Harris 1989; Millan 1990). However, the differences between the classes were smaller in our histochemical assay and were not helpful in further distinguishing the different isozymes.
The inhibition profiles of the bovine alkaline phosphatases in adult tissues proved to be useful for analyzing expression in preattachment bovine embryos. Bovine embryos transcribe mRNA for TSAP2 and TSAP3 starting at the 4-cell and 8-cell stages, respectively (McDougall et al. 1998). However, maternally derived activity is found in the oocyte and is presumably the predominant activity present on at least 1-cell to 4-cell embryos. The inhibition profile of alkaline phosphatase activity on oocytes through 8-cell embryos was distinctly that of TNAP, with inactivation at low temperature and high levamisole. Therefore, TNAP is the maternally-inherited isozyme and the predominate alkaline phosphatase during cleavage. By the 16-cell stage the inhibition was similar to that of cumulus cells, suggesting residual maternal TNAP plus a newly transcribed tissue-specific isozyme (either TSAP2 or TSAP3). After the 16-cell stage the profile was that of a tissue-specific alkaline phosphatase, indicating full replacement of maternal TNAP with embryonic TSAP2 or −3. In hatched blastocysts, TNAP message is again detected but the activity profile still is that of a tissue-specific isozyme, indicating that little or no functional TNAP is synthesized before implantation. Whether both TSAP2 and TSAP3 are translated in morulae and blastocysts could not be determined, but this study shows that TNAP is expressed during early cleavage and is supplanted by TSAP2 or TSAP3 by the morula stage.
Transition from a cleavage-stage embryo to a blastocyst is required for further development and is associated with a number of processes. The first burst of transcription from the embryonic genome in cows occurs at the 8-cell stage (Telford et al. 1990), and an outer epithelium (the trophectoderm) differentiates over the 16- to 32-cell stage (morula). During the equivalent stages in mice, there is evidence for chromatin remodeling and post-translational modifications in the activities of a variety of proteins commonly associated with epithelia (reviewed in Pratt 1989; Collins and Fleming 1995; Thompson 1996). Although there is strong evidence that phosphorylation is important in these events, there has been little research on the enzymes that dephosphorylate proteins in preimplantation embryos. We found that a homozygous null mutation of EAP in preimplantation mouse embryos is associated with loss of 8-cell stage embryos, when epithelialization of the outer cells begins (Dehghani et al. 2000). In the bovine, TSAP2 or TSAP3 protein activity is first detected in 16-cell stage embryos and morulae, as an epithelium begins to form in this species. This lends support to a hypothesis that a tissue-specific alkaline phosphatase is generally expressed by mammalian preimplantation embryos, and that it plays a role in forming the trophectoderm.
Footnotes
Acknowledgements
Supported by grants from NSERC Canada to AH.
We wish to thank Gencor, Inc. (Guelph, Ontario) for donating the bovine sperm used in IVF, the Animal Biotechnology and Embryo Laboratory for providing bovine ovaries, and Kevin O'Reilly for assistance in preparing the figures. KM was supported by an Ontario Government Scholarship.