
Abstract
We studied regulation of the AT2 receptor by investigating the effect of bilateral nephrectomy (bNX) in Sprague–Dawley rats. The expression of aldosterone synthase (CYP11B2) and AT2 receptor mRNA was detected by nonradioactive in situ hybridization. AT2 receptor mRNA was detected in cells of the first two or three subcapsular cell layers of the zona glomerulosa (ZG) and in the medulla of sham-operated animals. After bNX, the number and area of distribution of AT2 receptor-positive cells increased in the ZG. This was associated with an enlargement of the steroidogenic active ZG and with reduced proliferation rate (sham 5.9 ± 0.9%; bNX 2.4 ± 0.2%; p > 0.02). Infusion of angiotensin II (ANG II; 200 ng/kg/min SC for 56 hr) to bNX rats did not reverse the effect of nephrectomy on the distribution of AT2 receptor expression, although mRNA levels per cell were reduced compared to NX alone. ANG II infusion decreased proliferation rate further (0.4 ± 0.07%; p > 0.001). In the adrenal medulla after bNX, decreased expression of the AT2 receptor was associated with increased proliferation (2.6 ± 0.2% vs 6.6 ± 0.5%). These results demonstrate differential regulation of the AT2 receptor in the adrenal gland and suggest that expression of the AT2 receptor is involved in regulating proliferation and differentiation in the ZG and medulla.
(
T
In the rat adrenal gland, the AT2 receptor is expressed mainly in the medulla and to a lesser degree in the zona glomerulosa (ZG). Its physiological function in the ZG is largely unknown. Sodium depletion is associated with increased aldosterone production and growth of the ZG by proliferation. Under this condition, the level of AT2 receptor mRNA is slightly elevated (Lehoux et al. 1997).
We were interested in the regulation of AT2 receptor expression under conditions that increase the number of aldosterone synthase-expressing cells by differentiation processes rather than by proliferation. We chose bilateral nephrectomy (bNX) as an appropriate model. BNX increases aldosterone production and ZG size more prominently than sodium depletion, but we supposed this was unlikely to be accomplished by proliferation because an enlarged ZG is already present within a very short period of time, i.e., 2 days (Peters et al. 1998).
Materials and Methods
Animals, Nephrectomy, and Hemodialysis
Adult male Sprague-Dawley rats weighing 400–520 g were housed under alternating 12-hr light and dark cycles at a constant temperature between 20 and 22C. They were fed a standard laboratory chow [Ssniff (R/M-H); Soest, Germany) and had free access to tapwater. All animal experiments were in accordance with international ethical rules and appreciated by a governmental ethical commitee.
Under ketamine/xylazine anesthesia (75 mg/kg and 6 mg/kg bw), animals were either bilaterally nephrectomized (n = 12) or sham-operated (n = 6). They received chronic indwelling catheters into the femoral artery and vein and an intraperitoneally located osmotic minipump. Hemodialysis was performed every day after surgery as described previously (Peters et al. 1999). The bNX animals received either 0.9% NaCl (n = 6) or ANG II (n = 6) (200 ng/mg/min) via an osmotic minipump for a period of 55 hr, the sham group received 0.9% NaCl (n = 6).
Tissue Preparation
Rats were anesthetized with ketamine/xylazine (75 mg/kg and 6 mg/kg bw, respectively). After cannulation of the abdominal aorta just below the renal arteries, a retrograde perfusion was performed with 2% freshly prepared paraformaldehyde in PBS, pH 7.4, for 90 sec at a pressure level of 220 mmHg and for 90 sec at a pressure level of 170 mmHg. This was followed by a perfusion with 18% sucrose in PBS, adjusted to 800 mOsm/kg for another 3 min at the same pressure level. Adrenal glands were removed, gently sliced, mounted on small pieces of styrofoam, and quickly snapfrozen in liquid nitrogen-cooled isopentane.
Preparation of Riboprobes and In Situ Hybridization (ISH)
Full-length rat AT2 receptor cDNA (2.8 kb) was initially adapted and subcloned into the pcDNAI vector as described in the original report (Kambayashi et al. 1993). To generate antisense and sense RNA probes for ISH experiments, plasmids were linearized with Hind III and Not I, respectively. In vitro transcription with digoxigenin-labeled UTP (Roche Diagnostics; Mannheim, Germany) was performed using SP6 RNA polymerase (antisense) and T7 RNA polymerase (sense). To improve penetration into the sections during hybridization, transcripts were finally shortened by alkaline hydrolysis to a calculated average length of 250 bases.
A specific probe of CYP11B2 and CYP11B1 was generated by PCR from a full-length cDNA, which had been prepared from the rat adrenal cDNA.
The sense (5′-GGGATGTCATCTCTGAGTATG-3′) and antisense (5′-GATTGCTGTCGTGTCAAC-3′) primers used give rise to a fragment of 182 bp, corresponding to nucleotides 779–961 of CYP11B1, and a fragment of 185 bp, corresponding to nucleotides 779–964 of CYP11B2 (Mukai et al. 1993). Within this region of the sequence the highest dissimilarity to CYP11B1 and to CYP11B3 is found, with a homology of less than 70%. Amplification was performed with Pwo DNA polymerase (Roche Diagnostics) and the obtained fragment was cloned into pBluescript and sequenced. DIG1–11-UTP-labeled sense and antisense riboprobes were synthesized from the linearized plasmids using T3 and T7 RNA polymerases, respectively.
ISH has been described in detail previously (Obermüller et al. 1998a). Briefly, cryostat sections (6 μm thick) were transferred onto silane-coated glass slides and postfixed in 4% paraformaldehyde (in PBS, pH 7.4) for 20 min. After several washing steps and dehydration, sections were incubated with prehybridization solution (50% deionized formamide, 50 mM Tris-HCl, pH 7.6, 25 mM EDTA, 20 mM NaCl, 0.25 mg/ml tRNA from yeast, 2.5 × Denhardt's solution) at 45C for 2 hr, followed by incubation for 16 hr at 45C in a moist chamber with 25 μl hybridization mixture (final concentrations were 50% deionized formamide, 20 mM Tris-HCl, pH 7.6, 1 mM EDTA, 0.33 M NaCl, 0.2 M DTT, 0.5 mg/ml tRNA, 0.1 mg/ml sonicated, denatured DNA from fish sperm, 1 × Denhardt's solution, 10% dextran sulfate, and denatured riboprobe 10 ng/μl), covered with a siliconized coverslip. The post-hybridization washes included solutions of various degrees of stringency (from 1 × SSC with 50% formamide at 48C to 0.1 × SSC at room temperature). Incubation of slides with alkaline phosphatase-conjugated anti-digoxigenin antibody (dilution 1:500) and color reactions were performed according to the manufacturer's protocol (Roche Diagnostics). Slides were kept in a moist chamber in the dark. Color reaction was controlled under the microscope and terminated (usually after 12–48 hr) by immersing the slides for 5 min in 100 mM Tris-HCl, 1 mM EDTA, pH 8.0. After rinsing in PBS, sections were mounted in bicarbonate-buffered glycerol, pH 8.6.
The specificity of the ISH signal obtained was verified by parallel incubation with antisense and sense riboprobes on alternate sections. Throughout all experiments, sense probes did not produce any detectable signal. Antisense riboprobes for CYP11B2 and CYP11B1 were used on alternate sections as single probes and were applied simultaneously for double hybridization.
Immunohistochemistry
Immunohistochemistry was performed (n=3 per group) according to the method described previously (Bachmann et al. 1992). Presence of Ki-67 was determined using a highly specific antibody (Gerdes et al. 1997).The antibody was a kind gift of Dianova (rabbit anti-mouse Ki-67, polyclonal; Dianova, Hamburg; Germany). In corresponding cell sections, no difference was observed between the polyclonal antibody and the MIB-5 antibody.
Fresh frozen sections 7 μm thick were transfered onto silane-coated glass slides and air-dried for 20 min. All incubation steps were performed at room temperature. Sections were incubated with blocking solution (2% BSA, 0.15% Triton in PBS, pH 7.4) for 1 hr. Then the slides were incubated for 3 hr either with the first antibody, Ki-67 diluted 1:100 in 2% BSA, PBS, pH 7.4 or with normal rabbit serum diluted 1:100, as a negative control on alternate sections. They were washed three times with PBS for 5 min and then incubated in the dark for 50 min with Cy3-conjugated AffiniPure donkey antirabbit IgG (Dianova) diluted 1:600 in PBS. After three washing steps in PBS for 5 min, slides were covered with bicarbonate-buffered 66% glycine, pH 8.6. Throughout all experiments, no staining of nuclei was observed with the control normal rabbit serum.
Representative sections were photographed with a Polivar 2 (Firma Reichert Jung; Vienna, Austria), visualized with Richardson staining solution, and photographed again in the same position to identify the zonal localization of positive cells. A ratio of positively stained proliferating cells to the total number of cells was obtained by counting each 900–1000 cells of the ZG/ZI and 788–800 cells of the medulla per rat.
Results
AT2 Receptor Expression
AT2 receptor mRNA was observed in the adrenal gland of sham-operated animals mainly in the medulla (Figure 1A). The staining in the medulla was not homogeneous, although almost all cells in the medulla were positive with various degree of intensity. The intensity of the signal in the ZG was far weaker than in the medulla and was restricted to the cells of the first two or three cell layers subcapsularly. Staining was observed neither in the adrenal capsule, demonstrating specificity of the reaction, nor in the zona fasciculata or zona reticularis. CYP11B2 mRNA was detected in the first two or three subcapsular cell layers of the ZG only (Figure 1B).
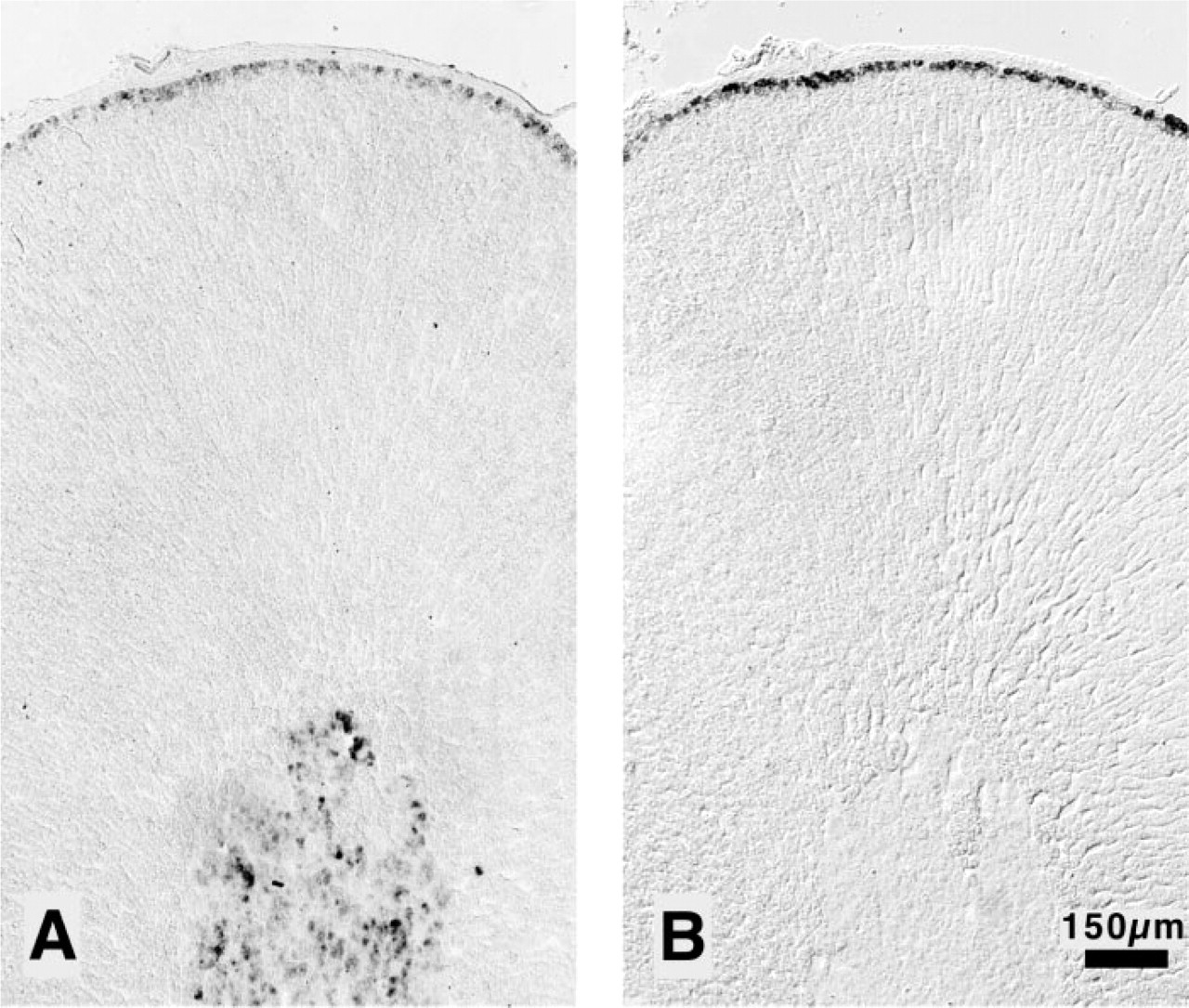
Localization of the AT2 receptor mRNA and aldosterone synthase mRNA by nonradioactive ISH. Representative cryostat sections of adrenal glands of sham-operated rats incubated with AT2 receptor antisense probe (
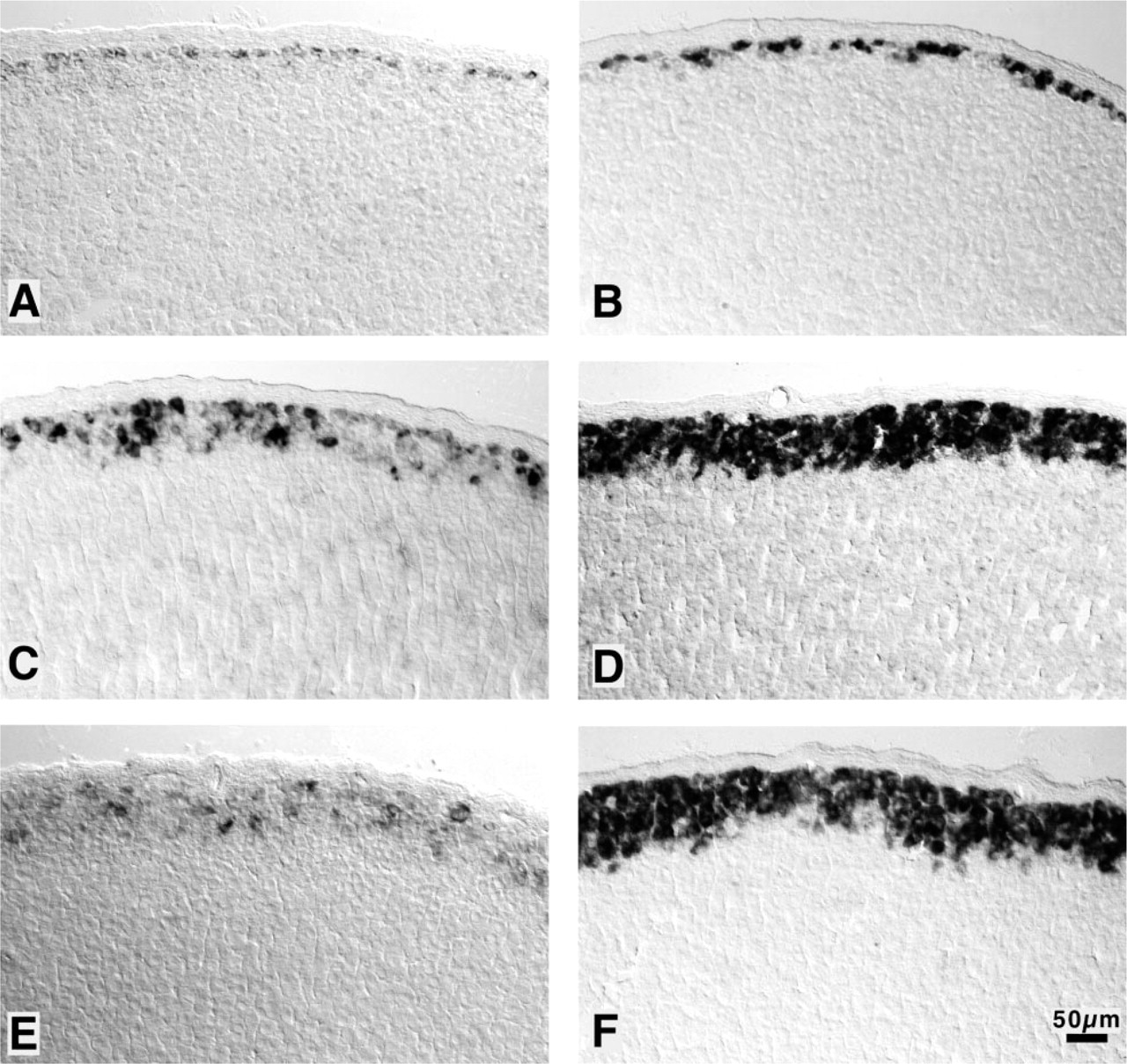
Effect of nephrectomy and ANG II resupplementation to bNX rats on the expression of the AT2 receptor and CYP11B2 in the adrenal cortex revealed by nonradioactive ISH. Left panels: AT2 receptor antisense probe; right panels, CYP11B2 antisense probe. Representative sections of adrenal glands of the three groups: (
Compared to sham-operated rats (Figure 2A), bNX led to a marked increase in the expression of AT2 receptor mRNA in the ZG (Figure 2C). The staining in the ZG became so intense that maximal staining was accomplished after only 6 hr of color reaction, whereas in sections of control rats the signal was even less intense after 48 hr. There was not only an increase per cell but also a change in the pattern of distribution. Concomitantly with the enlargement of the steroidogenic active ZG (from two or three to five or six cell layers), as defined by the expression of CYP11B2 mRNA (Figures 2B and 2D), the area of cells expressing AT2 receptor mRNA increased similarly from two or three to five to seven layers of cells. However, in contrast to the homogeneous distribution of CYP11B2 in bNX animals, distribution of AT2 receptor mRNA was not homogeneous. Only a few clusters of cells were found to express the AT2 receptor gene. ANG II infusion to bNX rats, although suppressing much of the increase of AT2 receptor mRNA, did not prevent the spread of AT2 receptor expression over an area of five to seven cell layers (Figure 2E).
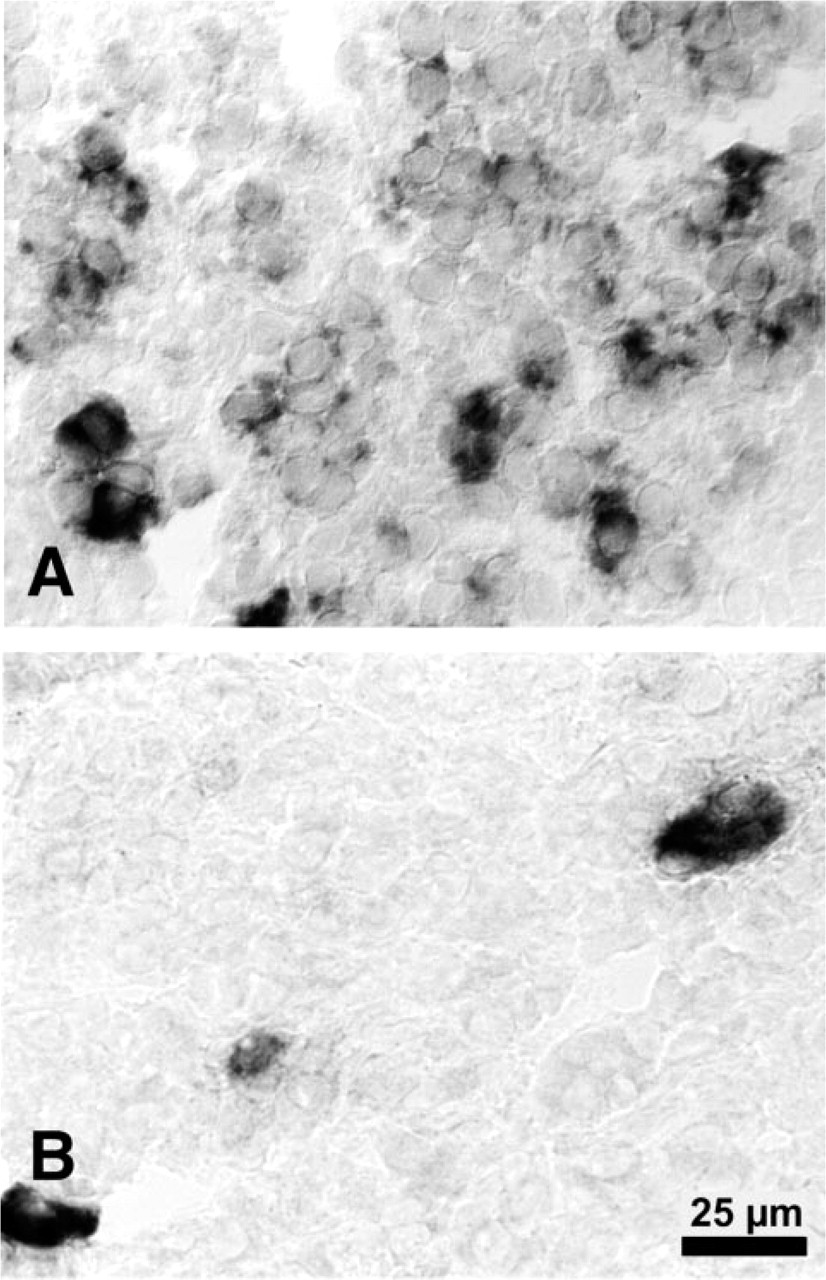
Suppression of AT2 receptor expression in the adrenal medulla after nephrectomy. Representative section of the adrenal medulla of nephrectomized rats (
In contrast to the increase of AT2 receptor expression in the ZG by bNX, this condition led to a suppression of the receptor in the medulla (Figures 3A and 3B). Only a few single cells still expressed AT2 receptor mRNA to a significant level after bNX. No marked changes were observed between ANG II- or NaCl-infused nephrectomized rats with regard to AT2 receptor expression in the medulla (data not shown).
Zonal Changes and Proliferation Indices
In sham-operated rats, CYP11B2 mRNA was detected in cells of the first two or three subcapsular cell layers only, with no crossreaction to CYP11B1, which was detected only in the ZF and ZR, decreasing gradually centripetally. A few single cells in the medulla were positive for CYP11B1 as well, reflecting the presence of steroidogenic cells in the medulla. The steroidogenic undifferentiated zone remained free of staining in the double ISH of CYP11B1 and CYP11B2 of control adrenals (Figure 4A).
After bNX, the cells of the ZG were markedly hypertrophic. Between ZG and ZF, a zone with cells free of staining could no longer be detected (Figure 4B. Only single cells remained negative for both probes, but unstained cells were also found deeper within the zona fasciculata. No difference was observed between bNX- and bNX/ANG II-treated groups (Figures 4B and 4C). The capsule remained free of staining.
In the ZG/ZI, the rate of proliferation was higher in sham-operated animals than in bNX animals (5.86 ± 0.93% vs 2.41 ± 0.24%; p > 0.02). The proliferation rate was even lower in bNX/ANG II than in both other groups (0.41 ± 0.07%; p > 0.001).
In contrast, in the adrenal medulla the proliferation rate increased in bNX (from 2.6 ± 0.2% in sham-operated rats to 6.6 ± 0.5% in bNX; p > 0.05). No significant difference was observed between bNX and bNX/ANG II (6.6 ± 0.5% vs 4.7 ± 1.1%; n.s.).
Discussion
Taking advantage of nonradioactive ISH with the use of specific riboprobes, it is possible not only to define the different zones of the adrenal cortex but also to localize the expression of a given gene to single cells. Under control conditions, we found AT2 receptor mRNA predominantly within the medulla, as expected, and to a lesser degree in subcapsular cells of the zona glomerulosa.
Some authors reported the presence of AT2 receptors not only in the zona glomerulosa but also in the zona fasciculata by use of immunhistochemistry (Ozono et al. 1997). Others could not confirm the expression of AT2 receptor in the ZF by radioactive ISH (Shanmugam et al. 1995) or autoradiographic binding studies (Lehoux et al. 1997). Although we can not exclude the possibility that cells of the zona fasciculata express the AT2 receptor at low levels, we observed AT2 receptor mRNA-positive cells exclusively in the ZG.
Little information is available about the regulation of the AT2 receptor in the adult rat adrenal gland. After a low-sodium diet, an increase of AT2 receptor mRNA has been observed in the ZG by Northern blotting analysis (Lehoux et al. 1997). Furthermore, it has been noted that bNX increases the total number of ANGII binding sites (Pernollet et al. 1977). With further specification, it has been demonstrated that after bNX AT1A receptor mRNA levels decreased, whereas AT1B mRNA levels increased (Iwai et al. 1992). The expression of the AT2 receptor gene has not been examined under this condition, nor has its expression been localized to single cells of the different cortical zones with respect to steroidogenic activity. We demonstrate here inverse changes of the AT2 receptor expression in the adrenal gland cortex and medulla in response to bNX. Specifically, the AT2 receptor mRNA increased in the glomerulosa and decreased in the medulla. In the ZG, not only did the expression of AT2 receptor mRNA per cell increase but also the number of positive cells. Furthermore, the increase in numbers of AT2 receptor mRNA-positive cells paralleled the increase of the steroidogenic active cell layers, because they were spread over the entire enlarged CYP11B2-positive area. Therefore, during activation of steroidogenesis the AT2 receptor expression is induced.
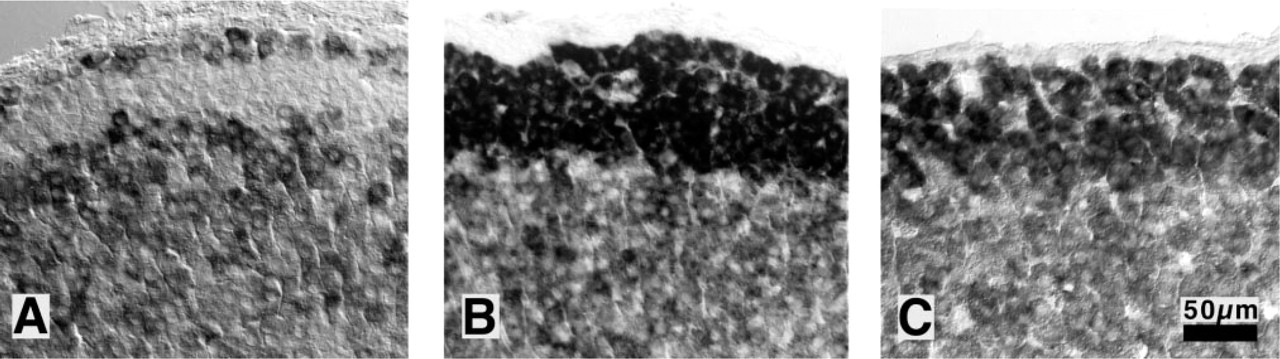
Effect of nephrectomy and ANG II resupplementation on CYP11B2 expression. Representative sections of double ISH with simultaneous use of CYP11B1 and CYP11B2 probes. In sham-operated animals (
It is noteworthy that the appearance of this receptor after bNX is associated with growth processes in the adrenal cortex of adult rats. Thus far, most studies focusing on the AT2 receptor were carried out in vitro using bovine adrenocortical cells or medullary PC12W cells. It has been shown that the AT2 receptor exerts an antiproliferative effect and even promotes apoptosis, depending on the cell type (Tsuzuki et al. 1996; Horiuchi et al. 1997). Although AT2 receptor expression was stimulated after bNX, the glomerulosa cells actively produced aldosterone in large amounts. Therefore, an increased rate of cell death in this area is rather unlikely. Another possible function for the AT2 receptor has been suggested by data from Meffert et al. (1996). These authors observed that AT2 receptor-mediated effects of ANG II induce differentiation processes in neuronal cells by inhibiting proliferation. The rapid increase in size of the glomerulosa and in aldosterone production after bNX in our study is not accomplished by an increased rate of proliferation but rather is a result of hypertrophy and recruitment of previously inactive cells of the ZG and the steroidogenic undifferentiated zone, which is located between ZG and ZF (Mitani et al. 1994). This is supported by the fact that the proliferation rate was low in bNX and the ZG was enlarged at the expense of the steroidogenic undifferentiated zone, which became undetectable.
In the adrenal cortex, a role for the AT1 receptor in modulating growth processes needs to be considered. The AT1R is known to increase proliferation. Reduced AT1R activation due to reduced levels of circulating ANG II and reduced levels of AT1AR could well account for the decreased proliferation observed. However, any activation of AT1R by additionally infused ANG II to bNX rats should have resulted in higher proliferation rates compared with vehicle-infused nephrectomized rats. Instead, a further inhibition was observed. This rules out the possibility that inhibition of proliferation is a result of diminished number of activated AT1R. In contrast, our data strengthen the hypothesis of an inhibitory effect of the AT2 receptor on proliferation. They do not, however, allow the conclusion that the AT2 receptor causes the NX-induced growth. Certainly, other factors, such as increased potassium levels, should be independently involved (Peters et al. 1999).
The adrenal changes that lead to a rise in aldosterone production observed after bNX are different from those observed with other stimulatory conditions. For instance, the enlargement of the ZG following sodium depletion is mainly due to proliferation and the undifferentiated zone migrates centripetally (Mitani et al. 1994). During sodium depletion, AT2 receptor prevents excess proliferation induced via AT1 receptor by elevated ANG II (Mazzocchi et al. 1997). In the case of bNX, one can consider that AT2 receptor enables differentiation processes by the same mechanism, i.e., by inhibiting proliferation. Pernollet et al. (1977) observed that a nephrectomy-induced increase in total ANG II binding sites in whole adrenal tissue could be abolished by ANG II resupplementation. In the present study, infusion of ANG II to nephrectomized rats decreased the amount of AT2 receptor mRNA in the ZG, compared to nephrectomy alone, but did not prevent the changes in its distribution. It was still spread over an area of five to seven subcapsular cell layers. Therefore, it can be assumed that factors other than withdrawal of ANG II induce the expression at that site in the first line, such as hyperkalemia. Interestingly, in this group the lowest rate of proliferation was observed. Although AT2 receptor mRNA levels per cell were lower in ANG II infused nephrectomized rats compared with vehicle-infused nephrectomized rats, ANG II is known to increase translation of AT2 receptor mRNA (Li et al. 1999), so that the decrease in AT2 receptor mRNA observed may have no functional consequences. Because the AT2 receptors in ANG II-infused bNX rats are also activated by the infused agonist in association with inhibition of proliferation, our data indicate antiproliferative effects of activated AT2 receptors. However, further studies are required to determine whether the AT2 receptor has any particular role in inducing cell arrest and differentiation processes of adrenocytes towards ZG cells.
In contrast to AT2 receptor regulation under a low-sodium diet, which leads to an increased expression in the ZG and the medulla (Lehoux et al. 1997), the AT2 receptor is differentially regulated after bNX. At present it is not known whether or not this is a different response of the zone to a single factor or rather responses to different factors involved in the complex pathophysiological context of bNX.
In contrast to the observations in the outer cortex, in the medulla we found a reduced level of AT2 receptor mRNA after bNX. At first view this was rather unexpected, because bNX is known to increase levels of catecholamines (Hatzinikolaou et al. 1983) and catecholamine release was found to be stimulated by ANG II via the AT2 receptor (Belloni et al. 1998; Martineau et al. 1999). Nevertheless, in the bNX model ANG II is unlikely to be the major factor releasing catecholamines. Interestingly, after bNX the reduced AT2 receptor mRNA in the medulla is associated with increased proliferation. Therefore, the growth of the medulla provides an explanation for the elevated catecholamine levels observed in the bNX model and the clinical state of uremia in humans (Izzo et al. 1987).
The model of bilateral nephrectomy may represent a useful tool to further study the function of the AT2 receptor in the zona glomerulosa and medulla and also to elucidate the derangement in catecholamine release observed in uremic patients.
Footnotes
Acknowledgements
Acknowledgments
Supported by Grant Pe 366/3–3 from the Deutsche Forschungsgemeinschaft and by the Forschungsfond der Fakultät für Klinische Medizin Mannheim of the University of Heidelberg.
We wish to thank Prof Wilhelm Kriz (Department of Anatomy and Cell Biology I, University of Heidelberg) for kindly providing facilities for histological analysis.