
Abstract
The high-pressure freezing/freeze substitution technique followed by Lowi-cryl K4M embedding provided an excellent ultrastructure and retention of antigenicity of rat gastric glands as well as the intraluminal fluid contents. By taking this advantage, we histochemically investigated the excretory flow of the zymogenic and mucin contents in rat gastric glandular lumen at the ultrastructural level. The combination of KMnO4-UA/Pb staining for zymogenic contents and Griffonia simplicifolia agglutinin-II (GSA-II) labeling for mucous neck cell (MNC) mucin distinguished the exocytosed zymogenic contents from the MNC mucin in the glandular lumen. Interestingly, at the base and neck regions, the zymogenic contents showed a droplet-like appearance, forming a distinct interface with the MNC mucin. At the pit region, the GSA-II labeling demonstrated restricted paths, designated as MNC mucous channels, which flowed into the surface mucous gel layer. It should be noted that the interface between exocytosed zymogenic contents and MNC mucin disappeared, and that the zymogenic contents merged into the MNC mucous channels. At the top pit region, the surface mucous gel layer showed laminated arrays of three types of gastric mucins. On the basis of these ultrastructural findings, we propose a model of the excretory flow in rat gastric gland.
Keywords
A CONTINUOUS VISCOELASTIC MUCOUS LAYER, secreted by the surface epithelial and the mucous neck cells, covers the gastric mucosa. It is primarily responsible for the protective properties and critical in the stomach. Zymogenic contents and acid secreted into the lumen of the gastric gland have to traverse the mucous layer to contribute to the digestion of food. Thus far, how they reach the gastric lumen is not fully understood. In in vitro studies, Bhaskar et al. (1992) demonstrated that the injection of HCl through solutions of pig gastric mucin produced viscous fingering patterns above pH 4. Recently, Johansson et al. (2000) revealed the existence of the in vivo acid transport through restricted channels in the mucous layer of rat stomach using Congo red staining at the light microscopic level. However, histological and histochemical visualizations of the excretory flow of zymogenic and mucin contents have been hindered by the difficulty of preserving them in conventional tissue preparations (Allen et al. 1993).
The high-pressure freezing (HPF) technique is capable of freezing up to 0.2-mm-thick biological specimens in a vitreous or microcrystalline state suitable for high-resolution structural analysis (Moor 1987; Studer et al. 1995). Thus far, many publications state that HPF processing is much superior to conventional processing in terms of excellent preservation of some protein epitopes, such as the distinct protein components of cuticlin of nematodes, CUT-1 and CUT-2 (Favre et al. 1995), the major cell cycle regulator complex cyclin B in the yeast Schizosaccharomyces pombe (Audit et al. 1996), the worm cuticular collagen and the vertebrate Type I fish scale collagen (Nicolas et al. 1997), the organic matrix components in the adult echinoid skeleton and tooth (Ameye et al. 1999) and filaggrin of human epidermis (Pfeiffer et al. 2000). In addition, Bordat et al. (1998) succeeded in demonstrating a precise calcium distribution in HPF bone cells by electron energy-loss spectroscopy and electron spectroscopic imaging. The application of HPF/freeze substitution to glycoconjugate histochemistry has provided excellent retention of some carbohydrate molecules of not only the epithelial cells of rat gastric glands but also the intraluminal exocytosed contents, suggesting the formation of a seromucous interface between the exocytosed zymogenic and mucin contents without diffusion (Suganuma et al. 1998,2001).
Recently, we have developed the potassium permanganate oxidation-uranyl acetate-lead citrate (KMnO4-UA/Pb) staining, which gives a differential staining of the zymogen granules of rat gastric chief cells on Lowicryl K4M ultrathin sections prepared by HPF/freeze substitution. It has been suggested that the KMnO4 oxidation evokes anionic sites on the zymogen granules which contribute to the binding of uranyl acetate (Sawaguchi et al. 1999). In the gastric glandular lumen, KMnO4-UA/Pb staining demonstrates the exocytosed zymogenic contents as electron-opaque images and allows us to follow the secretory dynamics of the zymogenic contents in rat gastric glands.
The mucous-secreting cells have been mainly classified into two types, i.e., the mucous neck cell (MNC) and the pit cell. The MNC mucin is specifically labeled with Griffonia simplicifolia agglutinin-II (GSA-II) (Ihida et al. 1988). The pit region has been subdivided equally into four segments, i.e., low, mid, high, and top pit (Karam and Leblond 1993), and the characteristics of the pit cells vary with their locations. For example, the low and mid pit cells secrete a sulfated mucin that is specifically labeled with cationic colloidal gold (CCG) at pH 1.0 (Yang et al. 1996). On the other hand, the high pit cell mucin is specifically labeled with monoclonal antibody RGM11, which recognizes a carbohydrate moiety of the mucin (Ishihara et al. 1993).
The aim of this study was to elucidate the in vivo fluid dynamics of the excretory flow in rat gastric gland. From the base to the top pit region, we carried out the combination of KMnO4-UA/Pb staining and colloidal gold labeling for various gastric mucins on Lowicryl K4M ultrathin sections prepared by HPF/freeze substitution. On the basis of our observations, we first propose a model of the excretory flow of zymogenic and mucin contents in rat gastric gland.
Materials and Methods
Reagents
The monoclonal antibody RGM11 was prepared by Dr. Ishihara (Ishihara et al. 1993). The monoclonal anti-phospholipase Cγ1 (PLCγ1) antibody was purchased from Santa Cruz Biotechnology (Santa Cruz, CA). Biotinylated GSA-II, goat anti-mouse IgM, and goat anti-biotin antibody were obtained from Vector Laboratories (Burlingame, CA). Goat anti-mouse IgG was purchased from Jackson ImmunoResearch Laboratories (West Grove, PA). Poly-L-lysine hydrobromide with molecular weight range of >300 kD was purchased from Sigma Chemical (St Louis, MO). N-Acetyl-D-glucosamine was from Nacalai Tesque (Kyoto, Japan).
Preparation of Colloidal Gold (CG), CCG, and Immunoglobulin-Gold Complex
Monodisperse CG of 8 nm in diameter was prepared by the method of Slot and Geuze (1985), and that of 14 nm in diameter was prepared by the modified method of Frens (1973). The conjugation of goat anti-biotin, goat anti-mouse IgM, or goat anti-mouse IgG antibody and CG were performed by the modified method of De Mey et al. (1981). Conjugation of poly-L-lysine and CG was prepared by the modified method of Goode et al. (1992) and Kashio et al. (1992) using bovine serum albumin (BSA) as a stabilizer (Torihara et al. 1995).
High-pressure Freezing
Male Wistar rats, 8–10 weeks old, were deeply anesthetized with diethylether and sodium pentobarbital and small fragments of the stomach were excised. They were promptly cut into 0.2–0.3-mm slices and sandwiched in the cavity of a supporting aluminum plate (Sawaguchi et al. 2001b). The space surrounding the specimen was filled with 1-hexadecene. This assembly was immediately frozen at 2100 bar pressure using a high-pressure freezing machine (BAL-TEC HPM 010; Liechtenstein) and rapidly transferred to liquid nitrogen for storage until required for further processing.
Freeze Substitution and Embedding
Freeze substitution was carried out using a Reichert AFS system (Leica; Vienna, Austria). After programmed warming to −90C at 3C/hr, samples were freeze-substituted at −90C for 20 hr in acetone containing 0.5% glutaraldehyde in which a molecular sieve had been added before use. After programmed warming to −30C at 10C/hr, the substitution medium was replaced with pure ethanol (three changes each of 10-min duration) and then gradually raised to 18C and left for 2 hr to remove the remaining hydration shell of a protein (Kellenberger 1991). After complete substitution, the temperature was gradually lowered to −35C and infiltration with Lowicryl K4M was performed in a mixture of 1:1 (v/v) 100% ethanol:Lowicryl K4M and 1:2 for 60 min each and in pure Lowicryl K4M overnight at −35C. Polymerization was performed under indirect, diffuse ultraviolet light (TTP 010/BAL-TEC) for 24 hr at −35C and for a further 24 hr at 18C.
Chemical Fixation
As a control experiment for cryoimmobilization, we prepared specimens by conventional processing. Rats were deeply anesthetized with diethylether and sodium pentobarbital and then perfused with 2% paraformaldehyde-2.5% glutaraldehyde in 0.1 M phosphate buffer (PB). The stomach was removed and further fixed by immersion in the above fixative at 4C overnight. After washing in PBS for 2 hr, specimens (about 1 mm3 in size) were dehydrated in a graded series of ethanol in water at progressively lower temperature: 30% for 30 min at 0C; 50% for 60 min at −20C; 70, 90, and 100% for 60 min at −35C. Infiltration with Lowicryl K4M and polymerization were performed as described above.
KMnO4-UA/Pb Staining
The KMnO4-UA/Pb staining was performed as described in the previous publication (Sawaguchi et al. 1999), with some modifications. Briefly, ultrathin sections (80–90 nm) of the specimens processed by HPF mounted on gold grids were oxidized with 0.1% KMnO4 in 0.1 N H2SO4 for 1 min at room temperature (RT). After washing with distilled water (DW), sections were stained with a saturated uranyl acetate aqueous solution (UA) and Reynolds' lead citrate (Pb) for 3 min and 40 sec, respectively (designated KMnO4-UA/Pb staining).
Combination of PLCγ1 Immunolabeling and KMnO4-UA/Pb Staining
Ultrathin sections of the cryoimmobilized specimens were treated with 50 mM Tris-HCl buffer, pH 7.2, containing 5% normal goat serum (NGS), 0.2% BSA, 0.3 M NaCl, and 0.1% Tween-20 for 30 min to block nonspecific binding and then incubated with anti-PLCγ1 antibody (diluted 1:30 with 1% BSA in PBS) at 4C overnight as a specific marker for the zymogen granules of rat gastric chief cells (Wang et al. 2000). After washing with PBS, the sections were incubated for 30 min with goat anti-mouse IgG antibody-CG (14 nm) conjugate (diluted with 1% BSA in PBS). After washing with DW and drying, the KMnO4-UA/Pb staining was performed as mentioned above. For controls, anti-PLCγ1 antibody was replaced by normal mouse serum or omitted from the procedure.
Combination of GSA-II Labeling and KMnO4-UA/Pb Staining
Ultrathin sections of the cryofixed specimens were treated with 1% BSA in PBS for 10 min to block nonspecific binding and then incubated with 5 μg/ml biotinylated GSA-II (diluted with 1% BSA in PBS) for 40 min at RT. After washing with PBS, the sections were incubated for 30 min with goat anti-biotin-IgG-CG (8 nm) conjugate (diluted with 1% BSA in PBS). After washing with DW and drying, the KMnO4-UA/Pb staining was performed as mentioned above. As a control experiment, GSA-II was pre-incubated with 0.2 M N-acetyl-D-glucosamine for 30 min and then applied to sections.
Combination of CCG Labeling at pH 1.0 and KMnO4-UA/Pb Staining
After blocking with 1% BSA in PBS for 10 min, the sections were passed through 50 mM Tris-HCl buffer containing 0.2% BSA (Tris-HCl/BSA) at pH 1.0 and then incubated with CCG (8 nm) at pH 1.0 for 40 min at RT. After brief incubation with Tris-HCl/BSA at pH 1.0, the sections were washed with DW. Subsequently, the KMnO4-UA/Pb staining was performed. As a control experiment, sections were pre-incubated in 0.1 mg/ml non-labeled poly-L-lysine in Tris-HCl/BSA at pH 1.0 for 30 min before incubation with CCG at pH 1.0.
Dual Labeling of CCG at pH 1.0 and GSA-II Followed by KMnO4-UA/Pb Staining
Ultrathin sections were first labeled with CCG (8 nm) at pH 1.0 and then labeled with the biotinylated GSA-II followed by goat anti-biotin-IgG-CG (14 nm) conjugate, according to each protocol mentioned above. Subsequently, the KMnO4-UA/Pb staining was performed.
Dual Labeling of RGM11 Antibody and GSA-II Followed by KMnO4-UA/Pb Staining
Ultrathin sections were first labeled with GSA-II as mentioned above. After washing with PBS, the sections were incubated with hybridoma medium of RGM11 antibody overnight at 4C. After washing with PBS, the sections were incubated with a mixture of goat anti-biotin-IgG-CG (14 nm) conjugate and goat anti-mouse IgM antibody-CG (8 nm) conjugate for 30 min. Subsequently, the KMnO4-UA/Pb staining was performed. For controls, RGM11 antibody was replaced by normal mouse serum or omitted from the procedure.
GSA-II Labeling for Conventionally Fixed Specimens
To compare with the cryoimmobilized specimens, ultrathin sections of the chemically fixed ones were stained with the biotinylated GSA-II followed by goat anti-biotin-IgG-CG (8 nm) conjugate described above. Then the sections were contrasted with UA/Pb staining.
Results
Base Region
The PLCγ1 immunolabeling was clearly co-localized on the zymogen granules as well as the exocytosed zymogenic contents, which intensively reacted to KMnO4-UA/Pb staining (Figure 1A). The combination of GSA-II labeling for MNC mucin and KMnO4-UA/Pb staining distinguished the zymogenic contents from the MNC mucin in the gastric glandular lumen (Figure 1B). The electron-opaque exocytosed zymogenic contents showed a droplet-like appearance forming the distinct seromucous interface. The intense GSA-II labeling was seen exclusively on the electron lucent area of the glandular lumen as well as the bipartite granules of intermediate cell (Suzuki et al. 1983; Suganuma et al. 1985; Ihida et al. 1988).
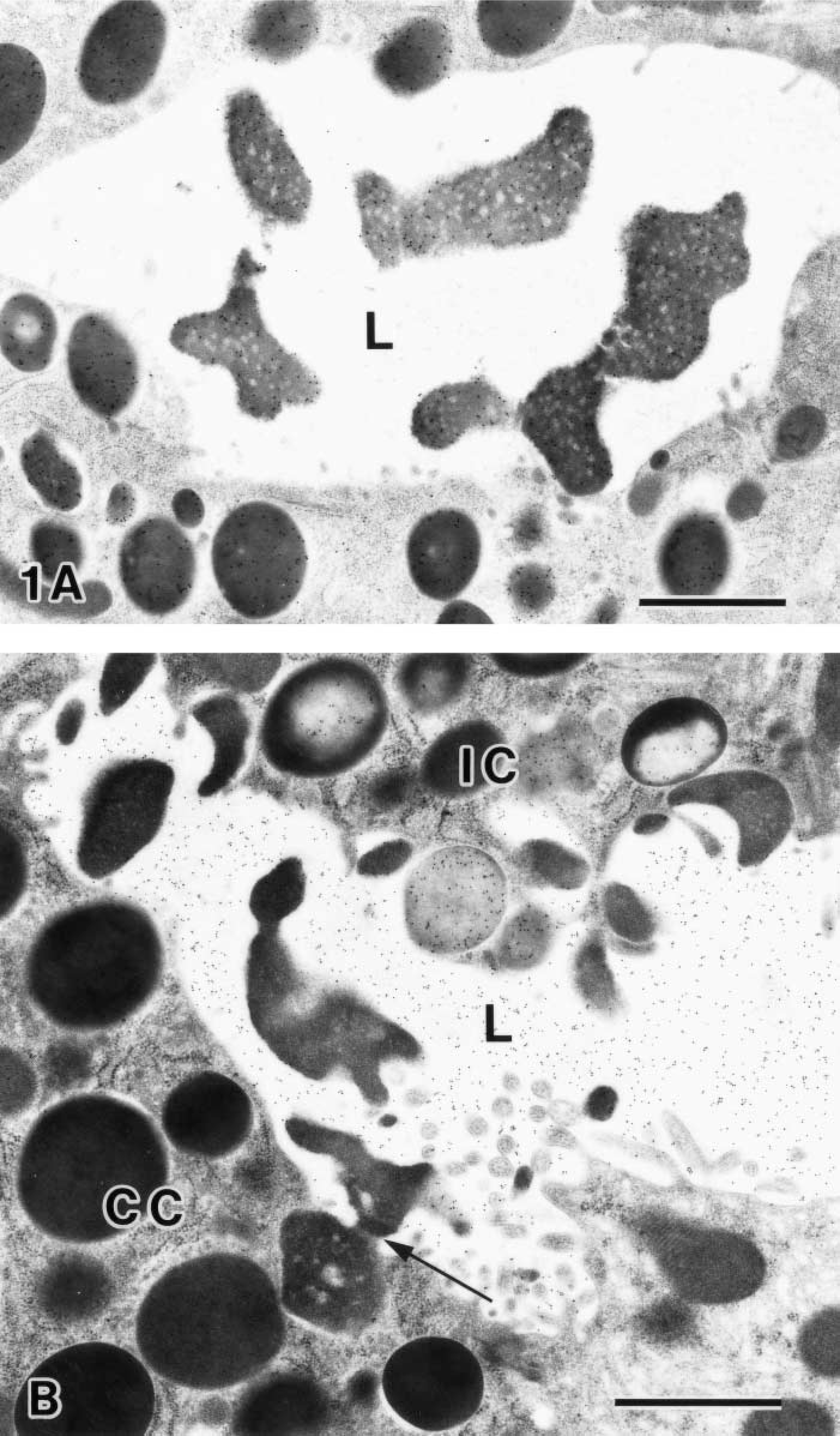
Base region of rat gastric gland. (
Neck Region
The combination of GSA-II labeling and KMnO4-UA/Pb staining revealed that the exocytosed zymogenic contents, ascending to the pit region, still kept the seromucous interface and remained the droplet-like appearance at the neck region (Figure 2A). It should be noted that the GSA-II labeling was seen on the electron-lucent area but was absent on the exocytosed zymogenic contents in the glandular lumen. For the chemically fixed samples, the intense GSA-II labeling was confined to the mucous granules and the apical membrane of MNCs. In contrast, only a few gold particles were found in the glandular lumen (Figure 2B). This indicated poor preservation of the intraluminal fluid contents processed by conventional fixation. It was noteworthy that the PLC71 immunolabeling still remained within the exocytosed zymogenic contents (Figure 2C.
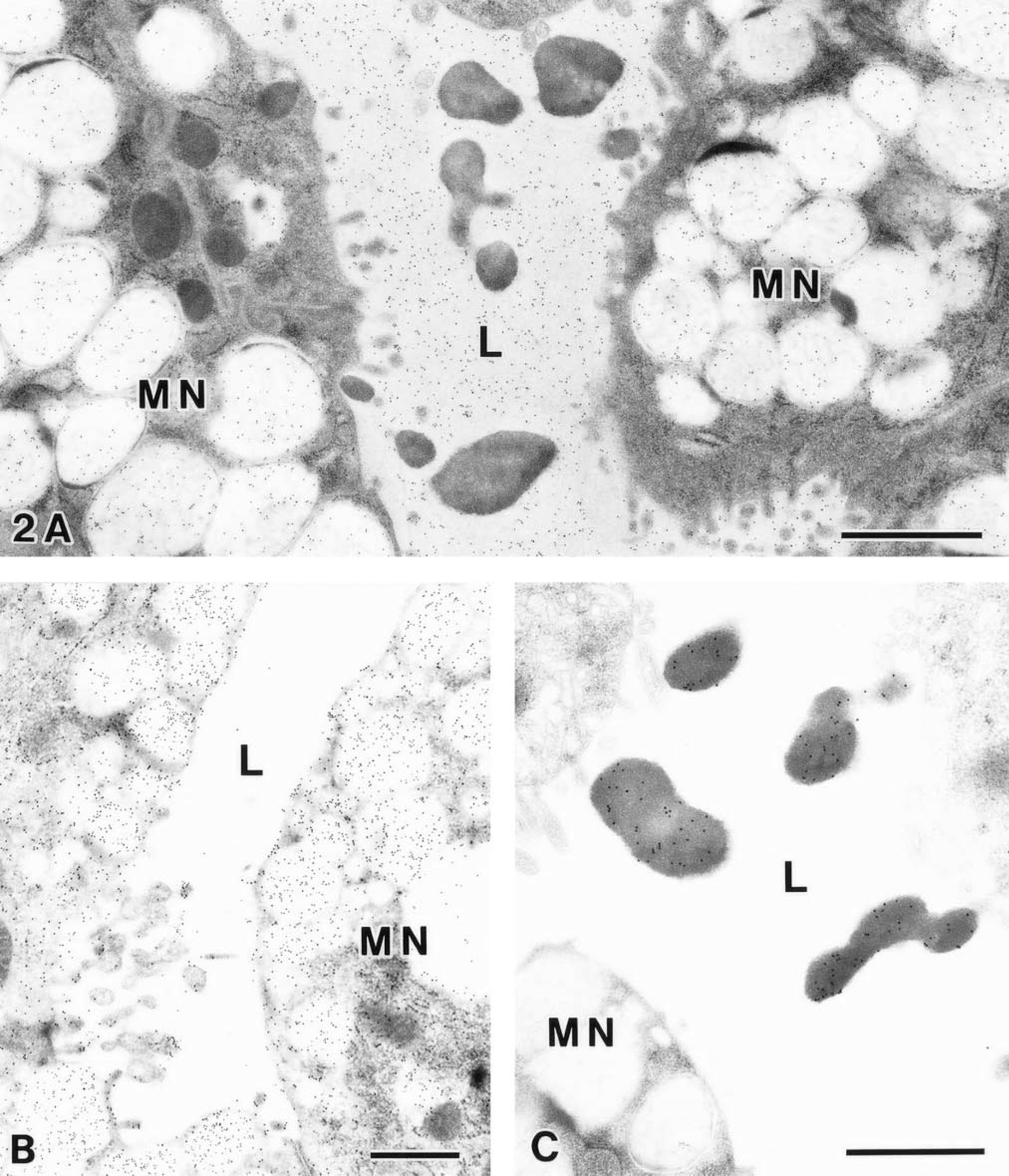
Neck region of rat gastric glands processed by HPF/freeze substitution (
Dual labeling of CCG at pH 1.0 (8-nm CG) and GSA-II (14-nm CG) followed by KMnO4-UA/Pb staining on a cross-section of the low pit region (
Isthmus Region
Undifferentiated, granule-free stem cells predominated in this region. The appearance of excretory flow in the glandular lumen was almost the same as that of the neck region (not shown).
Low and Mid Pit Regions
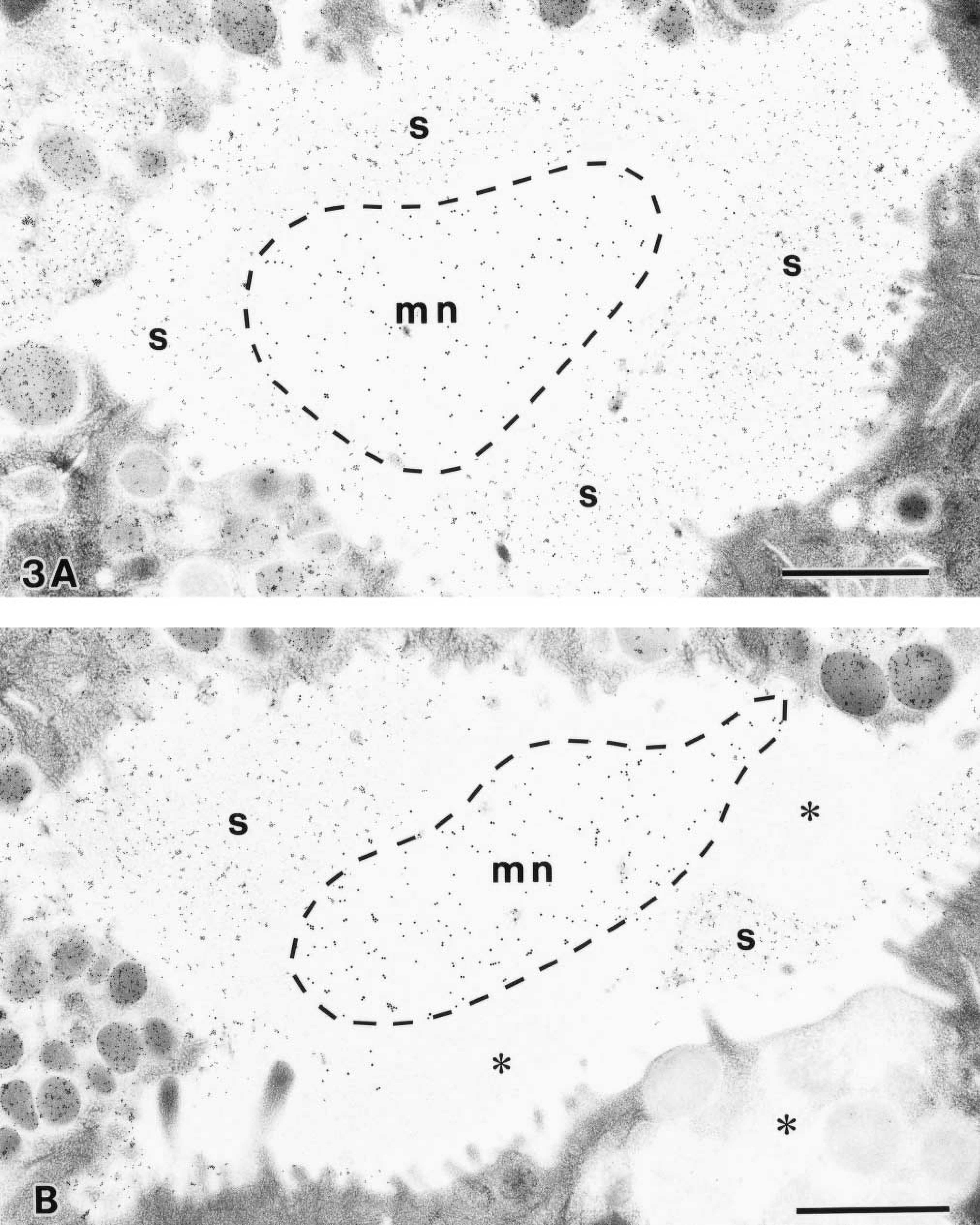
Another type of mucin, sulfated mucin, which was specifically labeled with CCG at pH 1.0, was secreted from the low and mid pit cells. In the glandular lumen, dual labeling of CCG at pH 1.0 and GSA-II differentially stained the sulfated mucin and the MNC mucin, respectively (Figure 3). On a cross-section at the low pit region, the MNC mucin was confined to the central portion of the glandular lumen, surrounded by the sulfated mucin secreted from the low pit cells (Figure 3A). It should be noted that a distinct interface was formed between the two types of mucins. At the mid pit region, the MNC mucin, the sulfated mucin, and the non-sulfated pit cell mucin faced each other, spatially occupying the distinct territories in the glandular lumen (Figure 3B).
High Pit Region
The combination of GSA-II labeling and KMnO4-UA/Pb staining demonstrated restricted paths, designated as channels, in the glandular lumen (Figures 4A and 4B). It should be noted that the appearance of exocytosed zymogenic contents changed into several electron-dense streaks with KMnO4-UA/Pb staining (Figure 4A). Close to the top pit region, the streaks of zymogenic contents were vividly labeled with GSA-II. These results imply that the zymogenic contents merge into the MNC mucin, forming several streaks. With dual labeling of RGM11 antibody and GSA-II, the MNC mucous channels were topologically surrounded by RGM11-positive mucin in the glandular lumen (Figure 4C). On the other hand, CCG labeling at pH 1.0 clearly demonstrated the sulfated mucous channels in the glandular lumen (Figure 4D). Furthermore, the dual labeling of CCG at pH 1.0 and GSA-II verified that both types of mucous channels merged with each other, forming topological compartments at the high pit region (Figure 4E).
Top Pit Region
The combination of GSA-II labeling and KMnO4-UA/Pb staining demonstrated the laminated arrays of the MNC mucin in the surface mucous gel layer (Figure 5A). The MNC mucin laminae showed high electron density with KMnO4-UA/Pb staining, indicating the presence of the zymogenic content. The CCG labeling at pH 1.0 also demonstrated the sulfated mucin laminae in the surface mucous gel layer (Figure 5B). Furthermore, dual labeling with the RGM11 antibody and GSA-II demonstrated an alternating laminated array consisting of MNC mucin and RGM11-positive mucin (Figure 5C).
Discussion
The HPF/freeze substitution method provided the deep vitreous freezing that permitted the cryo-immobilization of fluid contents in rat gastric glandular lumen. As shown in Figures 2A and 2B, the glandular lumen of the cryofixed gastric glands was filled with GSA-II-reactive mucin content and exocytosed zymogenic content, whereas that of the chemically fixed specimens was almost negative for GSA-II labeling. These results imply that conventional processing is inadequate to examine the intraluminal fluid contents and that HPF processing therefore appears to be superior in terms of preservation of extracellular fluid content close to the living state. By means of the combination of KMnO4-UA/Pb staining and colloidal gold labeling for various gastric mucins, we demonstrated the excretory flow in rat gastric fundic gland, from the base to the pit region, as summarized in Figure 6. As previously reported, the KMnO4-UA/Pb staining enhanced the overall contrast, without remarkable reduction of immuno-gold labeling, which allowed us to define the antigen localization with high accuracy (Sawaguchi et al. 2001a).
The combination of PLCγ1 immunolabeling and KMnO4-UA/Pb staining demonstrated that most of the exocytosed PLCγ1 molecules were co-localized to the electron-opaque areas, reactive with KMnO4-UA/Pb staining, in the glandular lumens. These results reconfirmed that KMnO4-UA/Pb sequence produced adequate differential staining not only of zymogen granules but also of exocytosed zymogenic contents in the glandular lumen (Sawaguchi et al. 1999; Wang et al. 2000).
The combination of GSA-II labeling and KMnO4-UA/Pb staining spatially distinguished the exocytosed zymogenic contents from the MNC mucin in the glandular lumen. Interestingly, at the base and neck regions, the exocytosed zymogenic contents showed a droplet-like appearance forming the distinct seromucous interface between the MNC mucin and the zymogenic contents. Previously, we applied periodic acid-thiocarbohydrazide-silver proteinate (PA-TCH- SP) staining on rat gastric glands processed by HPF/freeze substitution (Suganuma et al. 1998,2001). PA-TCH-SP staining produced negative images on the exocytosed zymogenic contents that were surrounded by the positively stained mucin contents forming the distinct interface in gastric glandular lumen. Taken together, these observations indicate the existence of a seromucous interface between the exocytosed zymogenic contents and the mucin contents in rat gastric glandular lumen.
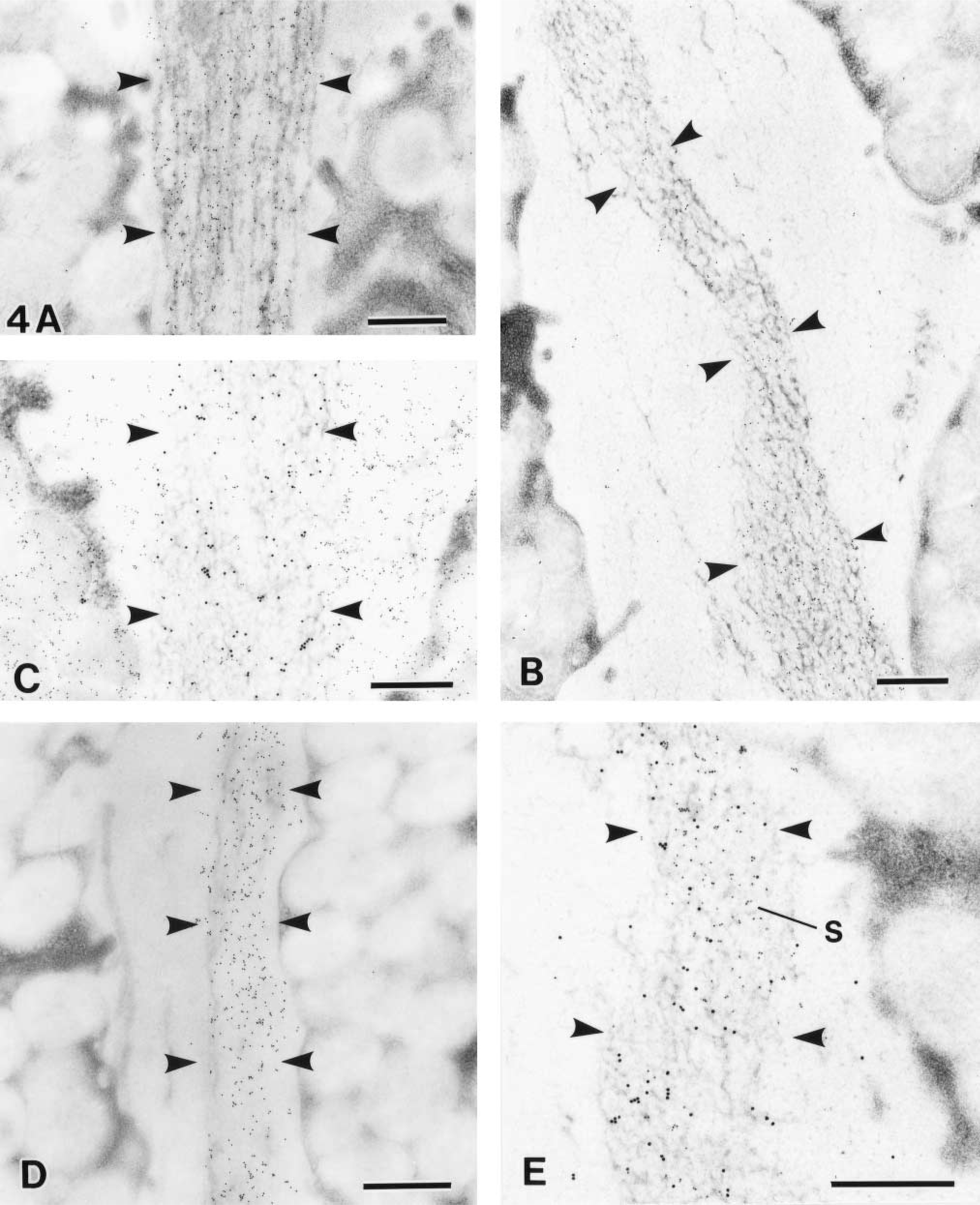
The high pit region of rat gastric glands. Arrowheads demarcate the intraluminal mucus channels. (
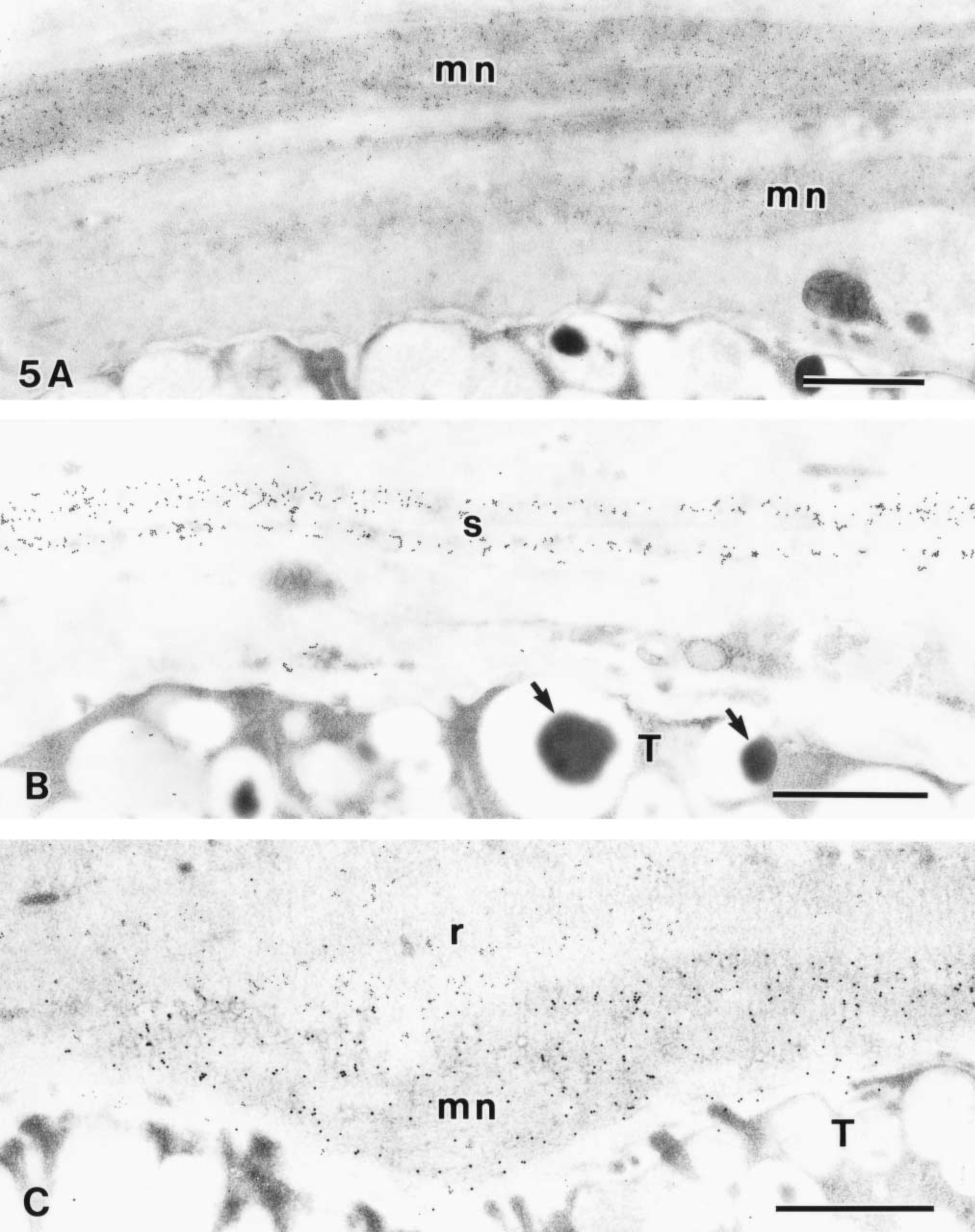
Laminated arrays in the surface mucous gel layer at the top pit region. (
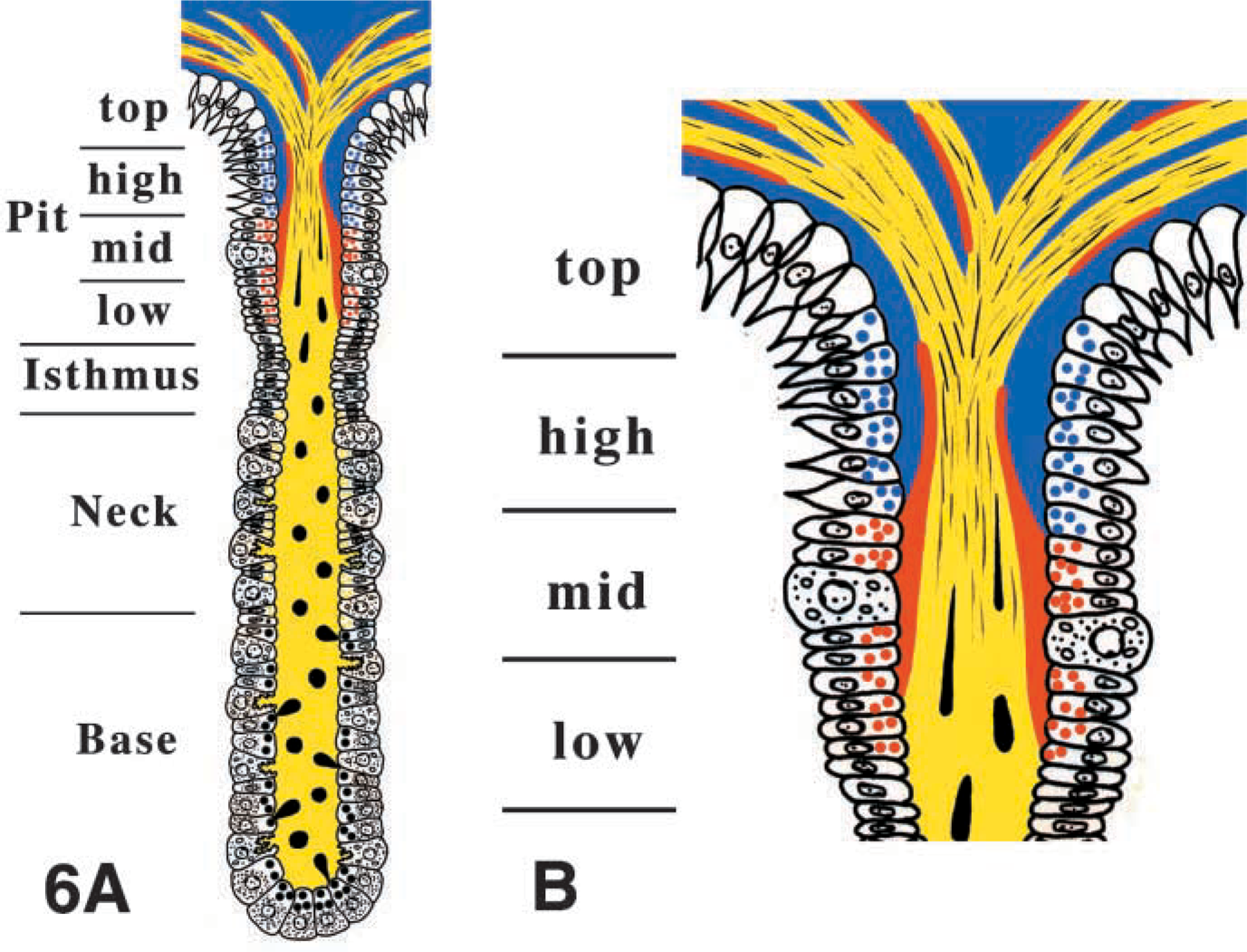
Schematic representation of the excretory flow, consisting of exocytosed zymogenic contents (black), MNC mucin (yellow), sulfated mucin (red), and RGM11 positive mucin (blue) in the glandular lumen of rat gastric gland. (
On reaching the mid or the high pit region, the droplet-like appearance of the zymogenic contents was altered in the bundles of several streaks and merged with the MNC mucous channels. These remarkable changes in rheological property may be explained by the proteolytic degradation of the MNC mucin (Allen 1978; Pearson et al. 1980; Bell et al. 1985). It has been reported that the proteolytic enzymes, such as pepsin, degrade the gastric mucous gel as an ongoing process in vitro (Kerss et al. 1982). In consequence, the properties of the mucous gel changed from water-insoluble to-soluble in the gastric glandular lumen (Allen and Carroll 1985). Because the diffusion constant depends on the relative size of the diffusant and the mesh or pore size of the mucous gel, it can be assumed that the diffusion of zymogenic contents would occur in proportion to the degradation of gastric mucins.
The present study, to the best of our knowledge, provides the first evidence of the in vivo intraluminal mucous channels in rat gastric glandular lumen. Ever since Bhaskar et al. (1992) demonstrated a discrete channel of HCl solution penetrating pig gastric mucin using an in vitro experimental model, the actual existence of intraluminal channel has been postulated in the in vivo gastric gland. Recently, Johansson et al. (2000) demonstrated the acid transport channels in the surface mucous gel layer of rat stomach by using the pH-sensitive dye Congo red. In addition to the acid transport channels, the existence of mucous channels has been postulated in the gastric glandular lumen (Allen 1989; Johansson et al. 2000). However, observation of excretory flow in the gastric glandular lumen has been hindered because of the difficulty in tissue preparation that would preserve the fluid contents in the glandular lumen. The HPF/freeze substitution of the rat gastric glands succeeded in preserving the fine structures of the fluid contents and in demonstrating the intraluminal mucous channels in rat gastric glandular lumen.
The surface mucous gel layer at the top pit region consisted of alternating laminated arrays of MNC mucin, sulfated mucin, and RGM11-positive pit mucin. Ota and Katsuyama (1992) first demonstrated the alternating laminated arrays of MNC mucin and pit cell mucin in the surface mucous gel layer of human gastric gland. It has been proposed that the alternating laminated arrays might play an important role in retarding the diffusion of harmful agents to the epithelial cell lining. It should be emphasized that Helicobacter pylori, a major etiologic factor of chronic gastritis and peptic ulcer, inhabits the surface mucous layer, some of which is attached to the pit cells (Akamatsu et al. 1995; Shimizu et al. 1996). Concomitantly, the alternating laminated arrays in the surface mucous gel layer were disturbed markedly where Helicobacter pylori was found abundantly (Akamatsu et al. 1995). It is highly anticipated that further investigations will disclose not only the physiological but also the clinicopathological significance of the alternating laminated arrays in the surface mucous gel layer.
These results are likely to reflect some physicochemical disparities among the different mucins. The rheological properties of a mucous gel are characterized by the parameters of viscosity and elasticity (Silberberg 1977). It has been postulated that the size and shape of the channel are dependent on the pH of mucins, because the gastric mucins form large aggregates and increase their viscosity at low pH (Bhaskar et al. 1991,1992). Further investigations are required to elucidate the mechanism of the fluid dynamics of the excretory flow, including acid channels, under the influence of pH.
In conclusion, we have found the in vivo paths of the excretory flow of zymogenic and mucin contents in rat gastric glands. The present histochemical findings are capable of providing some clues to the rheological and biochemical analysis of the fluid dynamics of gastric secretions.
Footnotes
Acknowledgements
Supported by grants-in-aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology of Japan (08670024) and from the Japan Society for the Promotion of Science (13670019).
We thank Yoshiteru Goto (Division of Electron Microscopy, Central Research Laboratories, Miyazaki Medical College), Soyuki Ide, and Eiko Matsuura (Department of Anatomy, Miyazaki Medical College) for expert assistance.