
Abstract
Data on the myosin heavy chain (MyHC) composition of human muscle spindles are scarce in spite of the well-known correlation between MyHC composition and functional properties of skeletal muscle fibers. The MyHC composition of intrafusal fibers from 36 spindles of human biceps brachii muscle was studied in detail by immunocytochemistry with a large battery of antibodies. The MyHC content of isolated muscle spindles was assessed with SDS-PAGE and immunoblots. Four major MyHC isoforms (MyHCI, IIa, embryonic, and intrafusal) were detected with SDS-PAGE. Immunocytochemistry revealed very complex staining patterns for each intrafusal fiber type. The bag1 fibers contained slow tonic MyHC along their entire fiber length and MyHCI, α-cardiac, embryonic, and fetal isoforms along a variable part of their length. The bag2 fibers contained MyHC slow tonic, I, α-cardiac, embryonic, and fetal isoforms with regional variations. Chain fibers contained MyHCIIa, embryonic, and fetal isoforms throughout the fiber, and MyHCIIx at least in the juxtaequatorial region. Virtually each muscle spindle had a different allotment of numbers of bag1, bag2 and chain fibers. Taken together, the complexity in intrafusal fiber content and MyHC composition observed indicate that each muscle spindle in the human biceps has a unique identity.
M
Myosin ATPase activity resides in the head region of the myosin heavy chain (MyHC) molecule, and its relative activity determines the shortening velocity of muscle fibers (Barany 1967). Two heavy chains and four light chains make up the myosin molecule, which is the major contractile protein in muscle fibers. MyHC exists in different isoforms encoded by a highly conserved multigene family (for review see Weiss and Leinwand 1996; Schiaffino and Salviati 1997) and it is the major determinant of the force-velocity properties of the muscle fiber (Bottinelli et al. 1996). Therefore, MyHCs are considered as the best markers for the functional properties of human skeletal muscle fibers (Larsson and Moss 1993). To date, at least eight distinct MyHC isoforms have been found in human skeletal muscles, and these MyHC isoforms are typically expressed in a tissue-specific and developmental stage-specific manner (Schiaffino and Reggiani 1996; Weiss et al. 1999).
Several lines of evidence suggest the existence of more than one slow MyHC (MyHCI) isoform, although only one corresponding gene has been identified to date (Schiaffino and Reggiani 1996), and it is generally accepted that each MyHC isoform is the product of one single gene (Weiss and Leinwand 1996). Using three different MAbs (A4.840, A4.951, and N2.261) recognizing distinct MyHCI epitopes, Hughes et al. (1993) have reported the sequential appearance of three MyHCI isofoms during human and rat muscle development. At the mRNA level, two different β-cardiac MyHC cDNAs, pHMC3 and pSM-HCZ, have been found in human muscle (Jandreski et al. 1987). Finally, different phenotypes among My-HCI-containing fibers have been reported in adult rabbit masseter muscle (English et al. 1998).
Although there is an extensive body of knowledge concerning the MyHC composition of human extrafusal muscle fibers, corresponding information on human muscle spindles is rare (Thornell et al. 1988, 1989; Pedrosa-Domellöf et al. 1993; Pedrosa-Domellöf and Thornell 1994; Eriksson et al. 1994). Data from other species have shown that the mammalian nuclear bag and nuclear chain fibers have a complex MyHC composition that differs from that of extrafusal fibers and varies along the fiber length (Kucera and Walro 1988,1989,1990; Kucera et al. 1992; Maier et al. 1988; Rowlerson et al. 1988; Pedrosa et al. 1989,1990; Pedrosa and Thornell 1990; Scapolo et al. 1990; Walro et al. 1997; Wang et al. 1997; for review see Soukup et al. 1995; Walro and Kucera 1999). In rat limb muscle spindles, for example, the bag1 and bag2 fibers preferentially react with antibodies against slow tonic MyHC (MyHCsto), MyHCI, α-cardiac MyHC (MyHCα) and, to a variable extent, with embryonic MyHC (MyHCemb). In addition, the nuclear bag2 fibers react with antibodies against fast-twitch MyHCII and neonatal MyHCneo, whereas the nuclear chain fibers mainly react with antibodies specific for MyHCneo and MyHCemb.
In this study we investigated the composition and distribution of MyHC isoforms in the muscle spindles of adult human biceps brachii muscle. This study is the first part of a large investigation aimed at thoroughly characterizing and comparing muscle spindles in human muscles with distinct architecture and functional demands.
Materials and Methods
Materials
Twelve muscle samples were collected shortly after death from the biceps brachii muscle of four females (age 48, 38, 19, and 15, six specimens) and six males (age 38, 37, 27, 25, 22, and 19) without known muscle-related disorders. The samples were collected according to the ethical recommendation of the Swedish Transplantation Law, with the approval of the Medical Ethical Committee, Umeå University.
The biceps muscle samples were mounted, rapidly frozen in liquid propane chilled with liquid nitrogen, and stored at −81C until sectioned. Frozen specimens were serially sectioned at −25C using a Reichert Jung cryostat (Leica; Nussloch, Germany). Sections to be used for demonstration of myofibrillar ATPase activity after preincubation at pH 10.4, 4.6, and 4.3 (Dubowitz 1985) were 8 (μm thick, those for immunocytochemistry were 5 (μm thick, and those for freeze-drying were 100 (μm thick.
Additional samples were collected shortly after death from adult extraocular, lumbrical, and heart muscles and frozen as above before SDS-PAGE analyses and immunoblotting. A sample of fetal (20 weeks of gestation) biceps brachii muscle (Pedrosa-Domellöf and Thornell 1994) and adult chicken anterior latissimus dorsi (Ald) muscle were also used for biochemical analyses.
Antibodies used to detect different MyHC isoforms
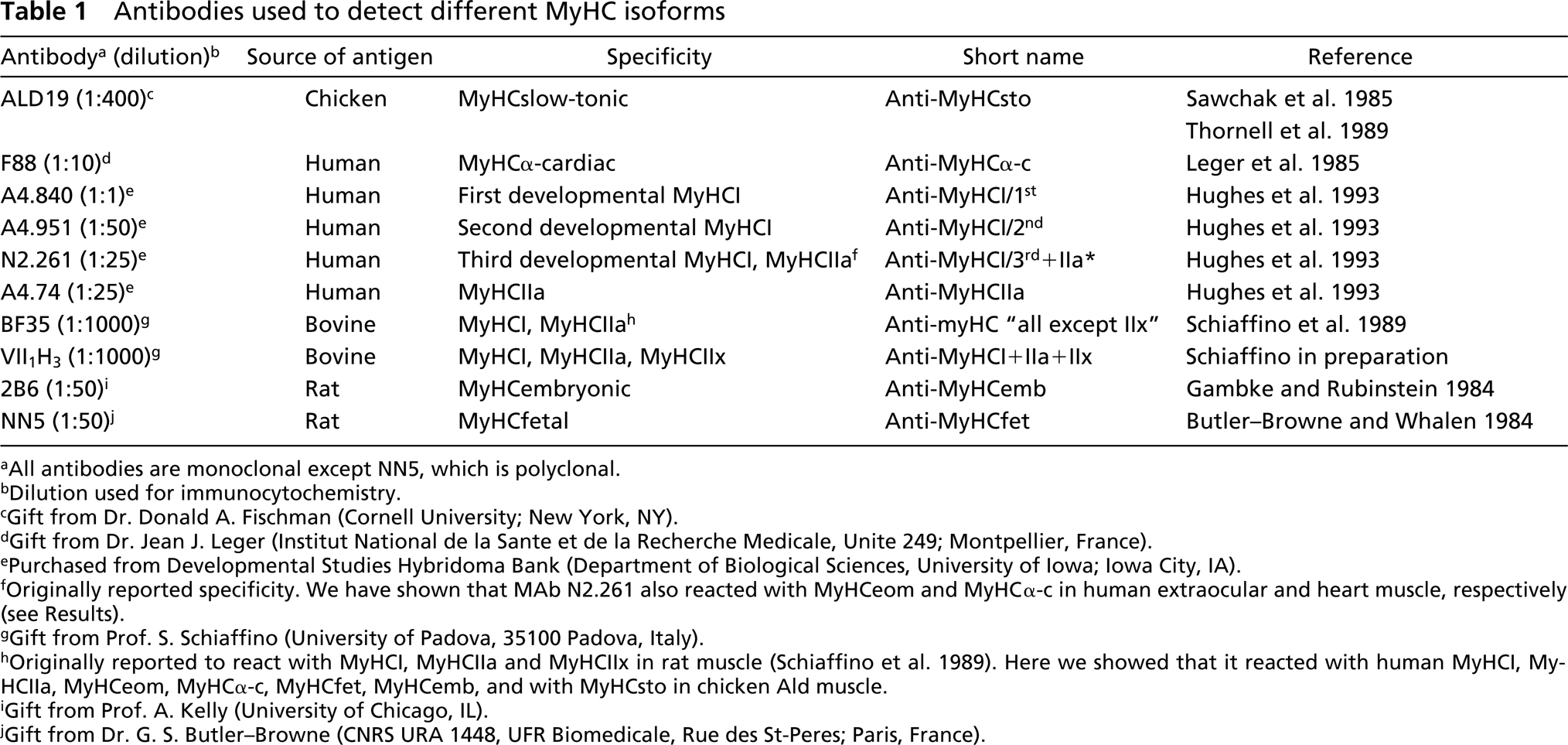
All antibodies are monoclonal except NN5, which is polyclonal.
Dilution used for immunocytochemistry.
Gift from Dr. Donald A. Fischman (Cornell University; New York, NY).
Gift from Dr. Jean J. Leger (Institut National de la Sante et de la Recherche Medicale, Unite 249; Montpellier, France).
Purchased from Developmental Studies Hybridoma Bank (Department of Biological Sciences, University of Iowa; Iowa City, IA).
Originally reported specificity. We have shown that MAb N2.261 also reacted with MyHCeom and MyHCα-c in human extraocular and heart muscle, respectively (see Results).
Gift from Prof. S. Schiaffino (University of Padova, 35100 Padova, Italy).
Originally reported to react with MyHCI, MyHCIIa and MyHCIIx in rat muscle (Schiaffino et al. 1989). Here we showed that it reacted with human MyHCI, MyHCIIa, MyHCeom, MyHCα-c, MyHCfet, MyHCemb, and with MyHCsto in chicken Ald muscle.
Gift from Prof. A. Kelly (University of Chicago, IL).
Gift from Dr. G. S. Butler-Browne (CNRS URA 1448, UFR Biomedicale, Rue des St-Peres; Paris, France).
Antibodies and Labeling
Monoclonal and polyclonal antibodies specific for different MyHC isoforms were used in the present study (Table 1). The specificity of these antibodies was carefully assessed for human muscles.
Immunocytochemistry
Immunocytochemistry was performed using the indirect peroxidase-anti-peroxidase (PAP) technique (Sternberger 1979) with a large battery of antibodies (Table 1). In brief, sections were air-dried for 20–30 min and immersed in 5% normal rabbit serum (for MAbs) (Dakopatts; Glostrup, Denmark) for 15 min. Sections were then incubated with the appropriate primary antibody for 60 min at 37C. After washing in 0.01 M PBS for 15 min, sections were immersed again in normal rabbit serum for 15 min, followed by incubation with 2% rabbit anti-mouse antibody (for MAbs; Dakopatts) for 30 min at room temperature (RT). After washing in 0.01 M PBS for 15 min, the sections were incubated with 1% mouse peroxidase-anti-peroxidase (Dakopatts) for 30 min and then washed in 0.01 M PBS for 15 min. Normal swine serum (5%), swine anti-rabbit antibody (1%), and 1% rabbit peroxidase-anti-peroxidase (Dakopatts) were used on the sections treated with the antibody against MyHCfet. Peroxidase developing was obtained by applying a solution containing diaminobenzidine and hydrogen peroxidase for 5–10 min, followed by rinsing with running water for 5 min. Finally the sections were dehydrated in graded dilutions of ethanol followed by xylene and mounted with DPX (BDH; Poole, UK). Control sections were processed as above, except that the primary antibody was replaced by normal serum from the same species and at the same dilutions. No staining was observed in the control sections.
All sections were examined and photographed with a Zeiss Axiophot microscope (Carl Zeiss; Oberkochen, Germany) equipped with a 3CCD camera (Dage-MTI; Michigan City, MI), using the Adobe Photoshop (version 5.5) software program.
Biochemistry
SDS-PAGE. Individual muscle spindles were dissected from freeze-dried sections under a stereomicroscope (Wild; Heerbrugg, Switzerland) and lysed in Laemmli sample buffer (Bio-Rad Laboratories; Hercules, CA) containing 5% β-mercaptoethanol. Whole muscle extracts were prepared as previously described (Kadi and Thornell 1999). SDS-PAGE (Talmadge and Roy 1993) was run (Mini Protean II; Bio-Rad, Glattbrug, Switzerland) at 75 V for 22 hr, with the lower two thirds of the gel unit surrounded by a 7C water bath. The gels were then silver-stained (Oakley et al. 1980) and photographed.

Electrophoretically separated MyHC content of extracts from human biceps muscle (Bic), chicken anterior latissimus dorsi muscle (Ald), human biceps muscle at 20 weeks of gestation (20wg), human extraocular muscles (Eoms), human heart muscle (Heart), and extracts of isolated muscle spindles from human lumbrical (MS1) and biceps (MS2) muscles.
Western Blotting Analysis
After SDS-PAGE, proteins were transferred to 0.45-μm nitrocellulose membrane (Bio-Rad) in a Tris-glycine buffer system using a Mini Trans-Blot apparatus (Bio-Rad) surrounded by a 15C water bath unit for 17 hr at 30-V constant voltage. The membranes were washed twice for 5 min with 20 ml distilled water to remove gel and transfer buffer components and weakly bound proteins. Immmunostaining was performed with WesternBreeze kit (Novex; Carlsbad, CA) according to the manufacturer's instructions (Western-Breeze Chromogenic Immunodetection Protocol) with the following primary antibodies (diluted in blocking solution): A4.840 (1:20), A4.951 (1:100), A4.74 (1:250), 2B6 (1:500), VII1H3 (1:10,000), ALD19 (1:500), N2.261 (1:250), and BF35 (1:3000).
Survey
A total of 36 muscle spindles (25 from females, 11 from males) were examined, 20 of them in consecutive serial transverse sections covering 1–3 mm of the length of the fibers. The spindles were divided into three regions: the A region, including equator (EQ) and juxtaequatorial parts (Aj), containing the periaxial fluid space; the B region, extending from the end of the periaxial fluid space to the end of the capsule; and the extracapsular C region (Barker and Banks 1994). Each region was further subdivided into an inner (proximal to the equator) and outer (distal to the equator) portion.
Statistical Analyses
The StatView software (Abacus Concepts; Berkeley, CA) was used for statistical analysis. Statistical comparison between males and females for the fiber numbers was performed with an unpaired t-test. Differences were considered significant at p<0.05. Group (gender) data were presented as mean and standard deviations (SD).
Results
Biochemistry
SDS-PAGE revealed the presence of four MyHC bands in muscle spindles (Figure 1) similar to our previous findings (Pedrosa-Domellöf et al. 1993). Identical results were obtained with muscle spindles isolated from the biceps and from the lumbrical muscles (Figure 1, lanes MS2, MS1). The latter were used for immunoblotting because they were more easily available. By immunoblotting and comparison to the whole muscle extracts from different muscles (Figure 2), it was determined that the most rapidly migrating band in the muscle spindle extracts corresponded to MyHCI and the slowest to MyHCemb. The third band, which displayed a slightly higher mobility than MyHCemb, was MyHCIIa. Finally, the prominent band migrating between MyHCI and MyHCIIa, tentatively named MyHCif (if = intrafusal; see Pedrosa-Domellöf et al. 1993), was found only in intrafusal fibers and was not recognized by MAbs N2.261 and BF35 (Figure 2) in the present study or by anti-MyHCsto, anti-MyHCemb, and anti-MyHCII in a previous study (Pedrosa-Domellöf et al. 1993).
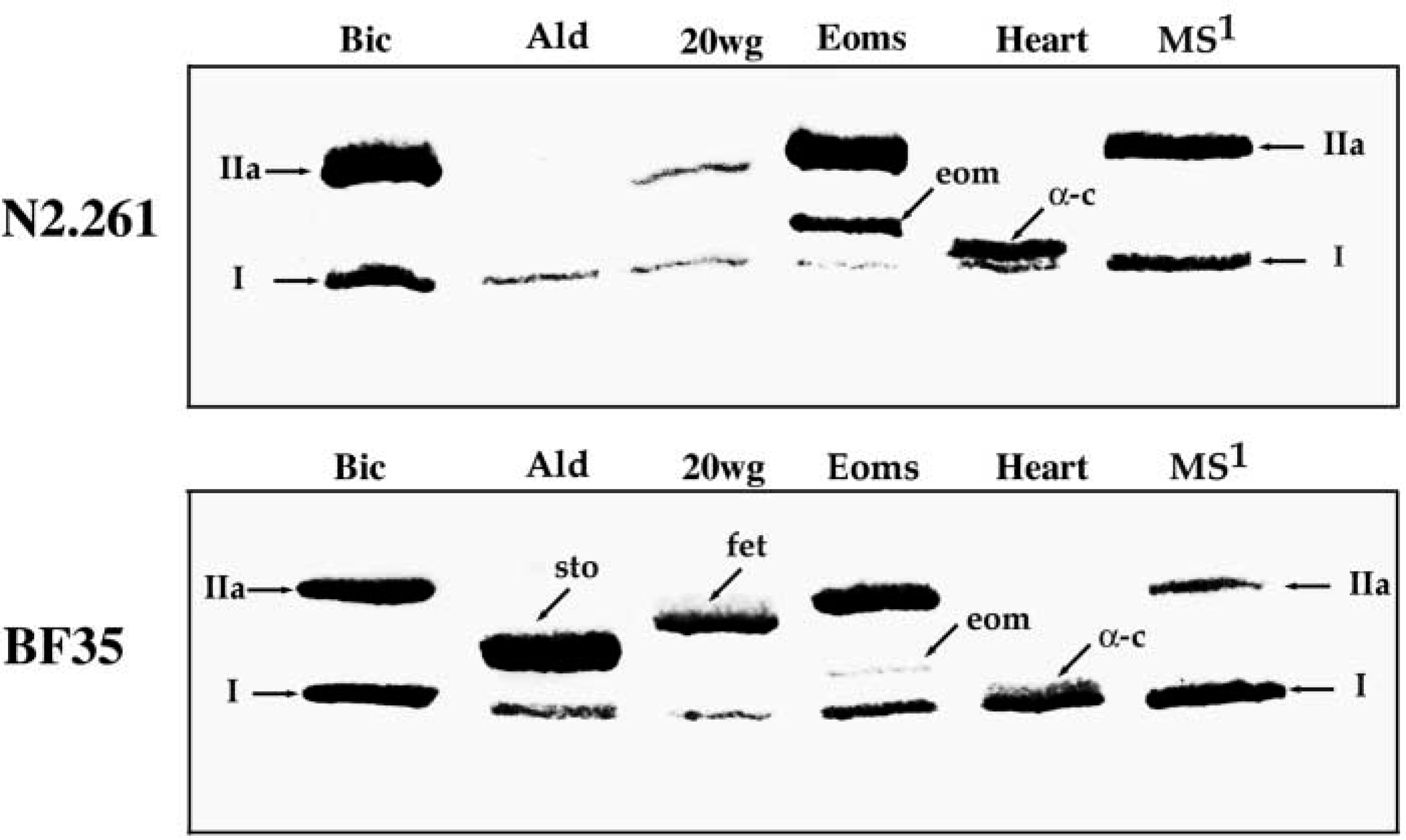
Immunoblots of MyHC isoforms from human biceps muscle (Bic), chicken Ald muscle (Ald), human biceps muscle at 20 weeks of gestation (20wg), extraocular muscles (Eoms), heart muscle (Heart), and muscle spindles of human lumbrical muscle (MS1), labeled with MAbs N2.261 (top) and BF35 (bottom). Both MAbs recognize MyHCI, MyHCIIa, MyHCeom, and MyHCα-c. In addition, BF35 recognizes MyHCsto and MyHCfet. The failure of MAb BF35 to detect MyHCemb could be due to the insufficient amounts of this isoform in the 20wg muscle sample. Note that none of the two MAbs reacted with MyHCif present in muscle spindles extracts. I, MyHCI; IIa, MyHCIIa; sto, MyHCsto; eom, MyHCeom; α-c, MyHCα-c.
The specificity of A4.840 and A4.951 to MyHCI, A4.74 to MyHCIIa, 2B6 to MyHCemb, and VII1H3 to MyHCI+IIa+IIx was confirmed for human muscles (data not shown). In addition, MAb ALD19 showed strong affinity to MyHCsto in chicken muscle (data not shown). MAb N2.261 (Figure 2) recognized MyHCI and MyHCIIa as previously described (Hughes et al. 1993), and also labeled the MyHCα-c band and the MyHCeom (eom = extraocular muscle), a band unique to the extraocular muscles (unpublished results). The specificity of MAb BF35 was further assessed for human muscle. It labeled MyHCI, MyHCIIa, MyHCeom, MyHCα-c, and MyHCfet (fet = fetal). This MAb also recognized the chicken slow-tonic MyHC, MyHCsto.
Intrafusal Fiber Type Composition
All the spindles investigated in the present study occurred as a single isolated receptor. In the 36 spindles, a total of 235 intrafusal fibers (183 from females, 52 from males) were examined and classified as 59 bag1 fibers, 46 bag2 fibers, and 127 chain fibers on the basis of their ATPase activity. The remaining three intrafusal fibers did not fit into any of these categories because of their atypical mATPase activity (see below). The fiber type composition of the muscle spindles varied considerably (Table 2). On average, there were 9.1 (range 4–16) intrafusal fibers per spindle, 2.2 (range 1–4) nuclear bag1, 1.3 (range 0–4) nuclear bag2, and 5.7 (range 2–12) nuclear chain fibers in each spindle. Intrafusal fibers occurring only in the C region were not counted. Strikingly, among 22 muscle spindles examined in the A or B regions, where the full complement of intrafusal fibers is present, only two spindles had exactly the same fiber composition (Table 2). There were no statistically significant differences either in the total number of intrafusal fibers (p = 0.20) per spindle or the number of bag1 (p=0.54), bag2 (p = 0.84), or chain (p=0.24) fibers per spindle between females and males. Furthermore, there were no distinct differences in staining patterns between males and females, so they will be described together.
ATPase Activity
In general, the myofibrillar ATPase activities of the bag fibers but not of chain fibers showed regional variation (Figure 3). Nuclear bag1 fibers displayed low alkaline ATPase activity over their entire fiber length, low acid ATPase activity along the A and B regions, and an intermediate acid ATPase activity in the C region. Nuclear bag2 fibers showed very weak acid ATPase activity in the equator, moderate in the remaining A region, and tended to become strong along the B and C regions. The alkaline ATPase activity of the nuclear bag2 fibers was very low in the equator but moderate along the remaining A region and most of the B region. Towards the end of the B region and in the C region, the activity was weak to moderate. In general, nuclear chain fibers showed high alkaline and low acid ATPase activity along the entire fiber length. Differentiation between bag2 and bag1 fibers in the C region could be carried out easily and precisely at pH 4.6, since very high activity appeared on bag2 fibers and moderate or low on bag1 fibers (see Figure 7B).
Intrafusal fiber type composition of individual spindles of adult human biceps brachii a
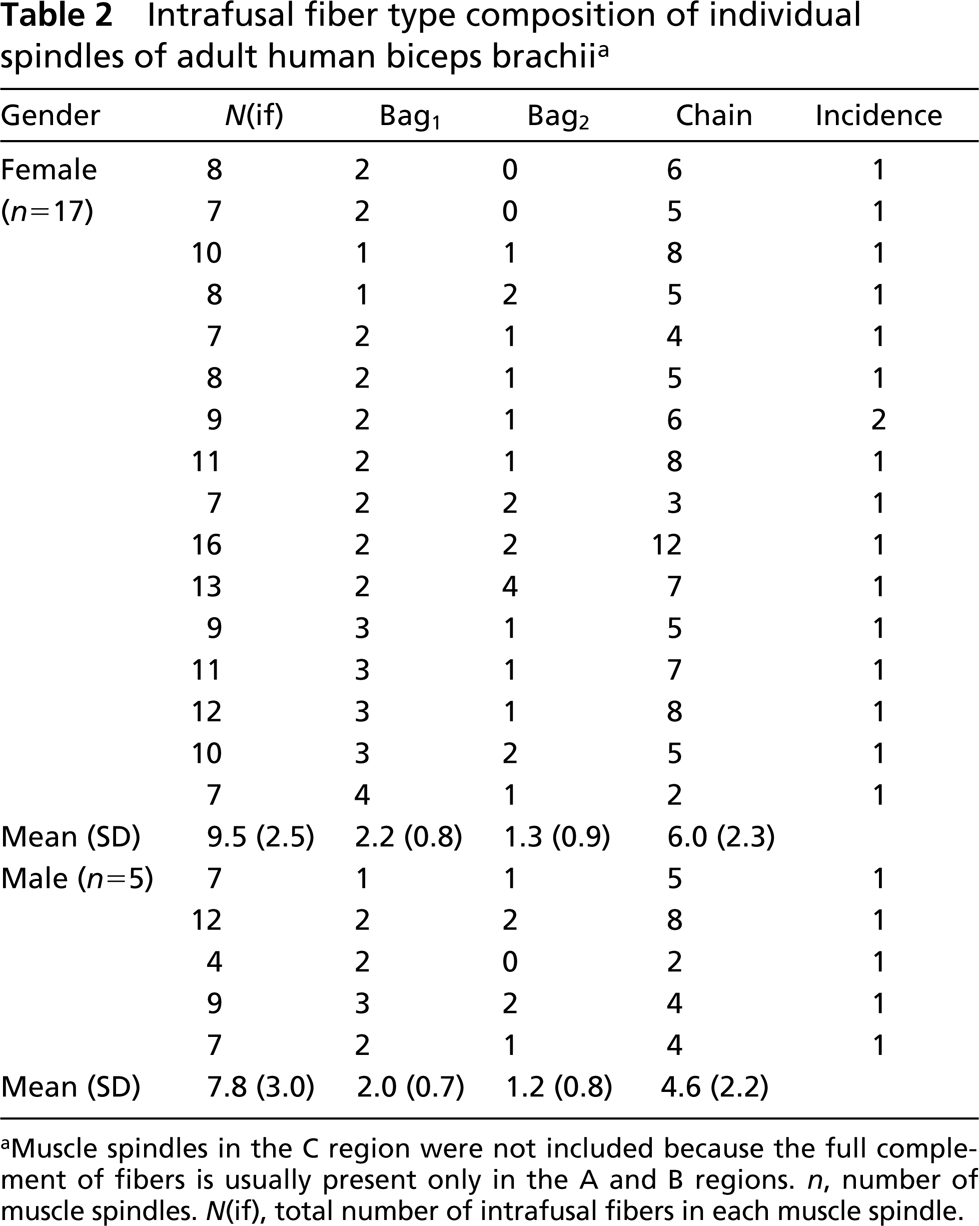
Muscle spindles in the C region were not included because the full complement of fibers is usually present only in the A and B regions. n, number of muscle spindles. N(if), total number of intrafusal fibers in each muscle spindle.
Although most intrafusal fibers conformed to the general ATPase activity described above, exceptions were observed. Three fibers exhibited atypical ATPase activity, showing strong or moderate alkaline but no acid ATPase activity. All these fibers were strongly and consistently stained with anti-MyHCsto.
Immunocytochemistry
Three intrafusal fiber types exhibited complex staining patterns with the different antibodies used (Figure 4).
Nuclear Bag1 Fibers. All the nuclear bag1 fibers were strongly and evenly stained with anti-MyHCsto along their entire fiber length (Figures 5B, 6B, and 7C). Bag1 fibers showed two different staining patterns with two different antibodies against MyHCI. They were weakly stained with anti-MyHCI/1st in the outer encapsulated pole and C region and were unstained along the remaining of their length (Figures 5C and 6C). However, bag1 fibers were moderately stained with anti-MyHCI/2 in the C and the outer B region, tended to become less stained from the inner B to the outer A region, and lost their staining towards the equator (Figures 5D, 6D, and 7D). MAb N2.261 labeled bag1 fibers in the C region only (Figures 5E, 6E, and 7E) and so did MAb BF35 (Figures 5H, 6H, and 7G), except that in one spindle two fibers showed low reactivity for a short distance in the A region. In general, the bag1 fibers were moderately stained with MAb VII1H3 in the extracapsular region, weakly in the B region, and unstained in the A region (Figures 5G and 6G). All bag1 fibers stained with MAb VII1H3 were also stained with anti-MyHCI/2 in the same region. The bag1 fibers showed weak staining with anti-MyHCemb in the A and B regions (Figures 5I and 6I). In the C region, more than half of the fibers were weakly to moderately stained (Figure 7H); the remainder were unstained with this antibody (not shown). Although the bag1 fibers were generally weakly stained by anti-MyHCfet, fibers that were unstained or moderately stained for a short distance in the A or B region were also encountered (Figure 8). The staining intensity of bag1 fibers with anti-MyHCα-c varied among different fibers and different spindles. At the equator, bag1 fibers showed a wide variation of staining intensity, from strong to weak. Bag1 fibers were often strongly to moderately stained in the juxtaequatorial region, less stained along the inner B region, and became negative in the remaining B and C regions (Figures 5J and 6J). However, the staining patterns of four bag1 fibers from one spindle were rather peculiar. These fibers were strongly stained in the juxtaequatorial region and unstained both in the B region and the equator. There were no fibers extending into the C region in this particular muscle spindle. None of the bag1 fibers reacted with anti-MyHCIIa (Figures 5F, 6F, and 7F).
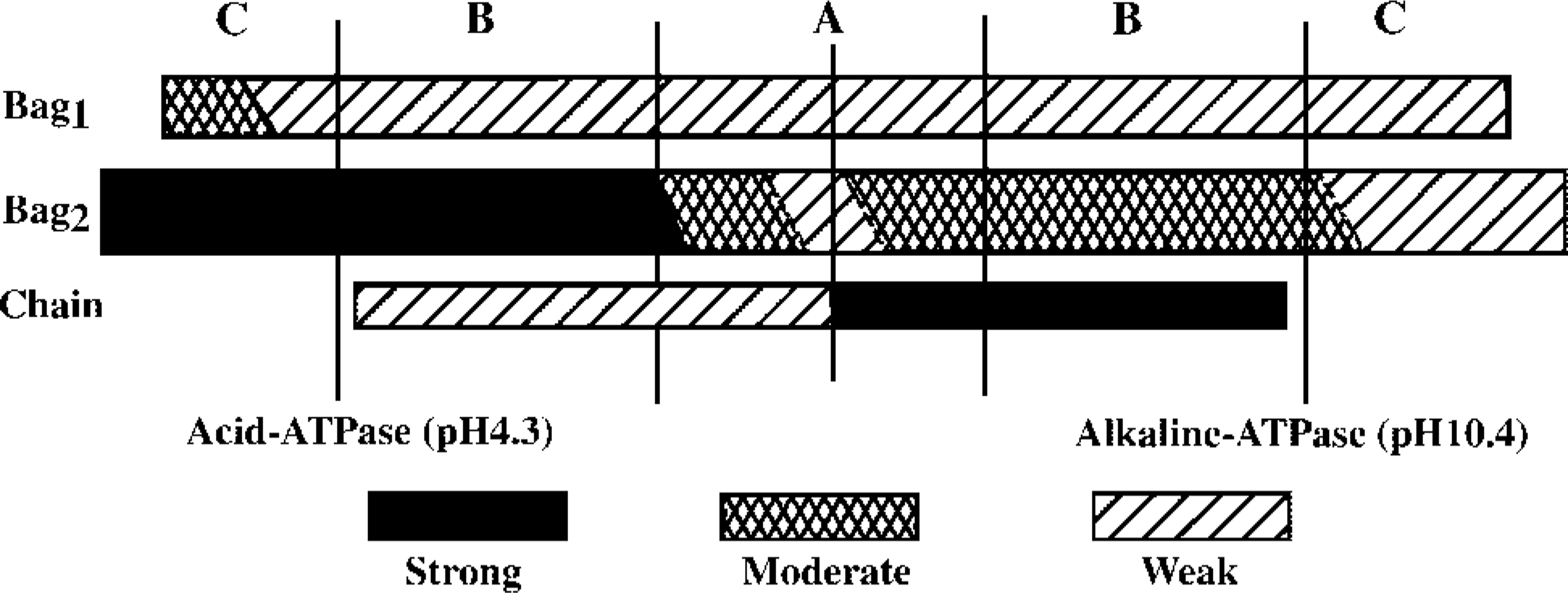
Schematic representation of the mATPase activity after acid (pH 4.3) and alkaline (pH 10.4) preincubations along the length of three types of intrafusal fibers from human biceps. Three different levels of staining intensity are distinguished.
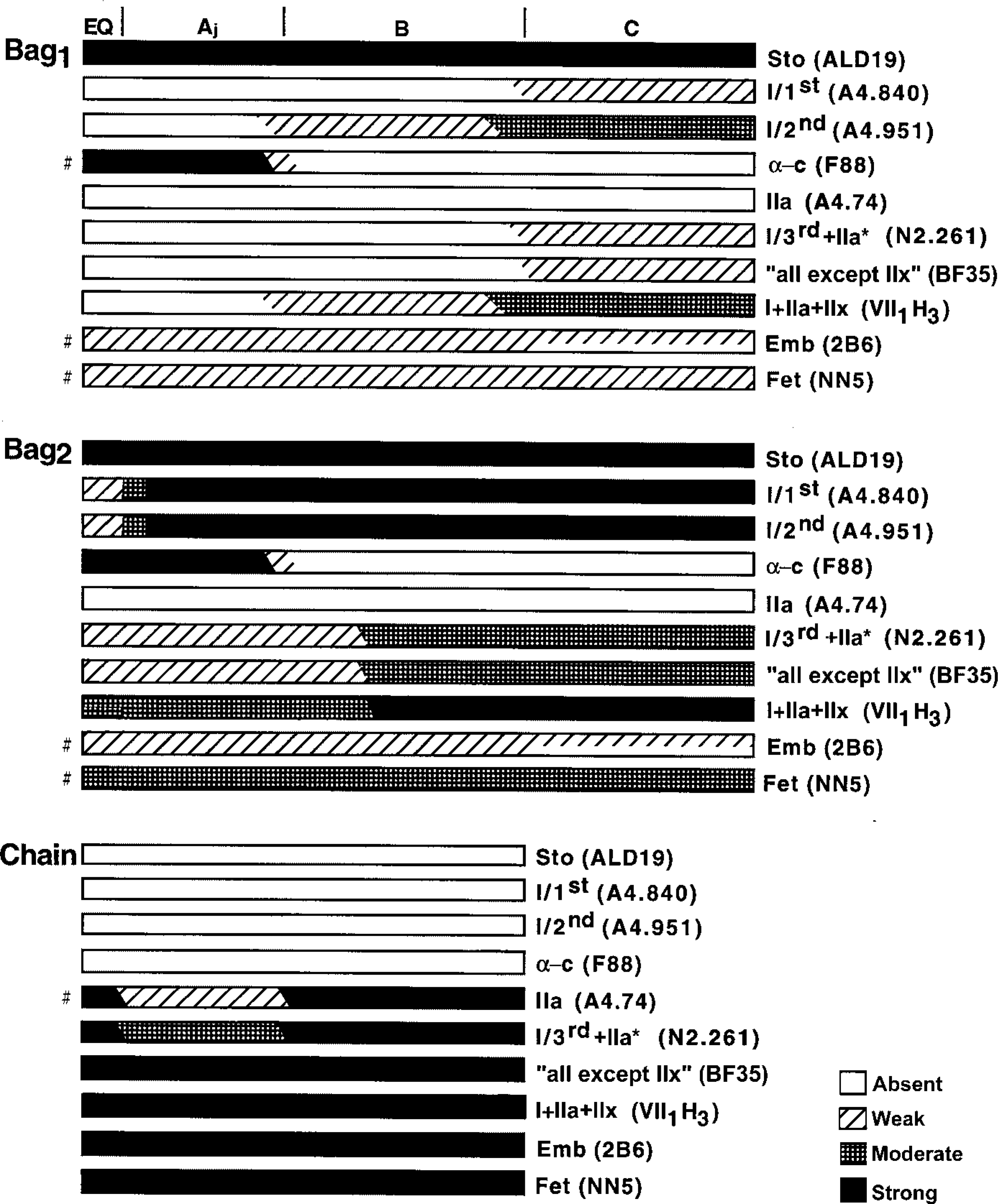
Schematic representation of the staining profiles of nuclear bag1, bag2, and chain fibers from adult human biceps muscle with the different MyHC antibodies. The A, B, and C regions are marked at the top of the fibers and the vertical bars represent, from left to right, the end of the periaxial space and the end of the capsule. The representation of staining profiles of chain fibers is lacking in the C region because few chain fibers extended into this region. The symbols “#” on the left denote some transient variations with regard to the general staining profile, and which are described in detail in the Results. Not drawn to scale.
Nuclear Bag2 Fibers. Nuclear bag2 fibers showed strong and homogeneous staining with anti-MyHCsto (Figures 5B, 6B, and 7C). They were strongly labeled by both anti-MyHCI/1st and anti-MyHCI/2nd over their entire length except at the inner juxtaequatorial and the equatorial regions, where they were moderately and faintly stained, respectively (Figures 5C, 6C, 5D, 6D, and 7D). Bag2 fibers were stained identically with MAbs N2.261 and BF35, moderately along the C and B regions, weakly in the juxtaequatorial region, and tended to become very weakly stained or unstained at the equator (Figures 5E, 6E, 7E, 5H, 6H, and 7G). These fibers were strongly stained with MAb VII1H3 in the C and outer B regions, moderately in the inner B and A regions (Figures 5G and 6G). Two bag2 fibers, however, showed weak or negative staining in the equator and the latter fiber was unstained with MAbs A4.951, N2.261, A4.74, and BF35. Bag2 fibers showed the same staining patterns as bag1 fibers did with anti-MyHCemb, although two bag2 fibers in one spindle showed high activity in the A region (Figures 5I, 6I, and 7H). The bag2 fibers showed a heterogeneous staining pattern with anti-MyHCfet. In some of the spindles, the bag2 fibers were moderately to strongly stained (Figure 8), whereas in other spindles they were weakly stained. In general, bag2 fibers were strongly to moderately stained with anti-MyHCα-c in the A region, less stained in the inner B region, and became unstained in the remaining portion (Figures 5J and 6J). Bag2 fibers generally did not react with anti-MyHCIIa (Figures 5F, 6F, and 7F), although two fibers showed weak staining in the equatorial region (not shown).
Nuclear Chain Fibers. Nuclear chain fibers were definitely unstained with anti-MyHCsto, anti-MyHCI/1st, anti-MyHCI/2nd, and anti-MyHCα-c, whereas they were strongly and uniformly stained with MAbs VII1H3 and BF35 and with anti-MyHCemb and anti-MyHCfet. All nuclear chain fibers were strongly stained with MAb N2.261, except that in the juxtaequatorial region almost one third of the fibers were moderately or even weakly stained (Figure 5E). Nuclear chain fibers were generally strongly stained with anti-MyHCIIa in the equatorial region and in the outer B and C regions. The staining intensity varied in the juxtaequatorial region, and variability was observed not only along the individual fibers but also between spindles. Most chain fibers were strongly stained, but some were weakly stained or even unstained (Figure 5F). These MyHCIIa-negative chain fibers, however, were always positive with both VII1H3 and N2.261 MAbs.
Discussion
The present investigation yielded major results on the specificity of MAbs N2.261, BF35, and the MyHC composition of human biceps muscle spindles. Taken together, the data showed that the muscle spindles in the biceps are by far more complex than anticipated.
A thorough characterization of the Abs used was a prerequisite for interpreting the immunocytochemical data. The present study further extended our knowledge of the MyHC affinity for MAbs N2.261 and BF35 in human muscle to include MyHCα-c and MyHCeom (N2.261 and BF35) and MyHCsto (BF35).
Four MyHC isoforms were present in intrafusal fibers in amounts sufficient to be detected by SDS-PAGE. The present study confirmed the presence of MyHCI, MyHCif, and MyHCemb (Pedrosa-Domellöf et al. 1993). The band migrating slightly more rapidly than the band of MyHCemb has now been characterized by immunoblotting as MyHCIIa rather than MyHCfet, as previously assumed (Pedrosa-Domellöf et al. 1993). We could confirm the unique position of the MyHCif band, which was distinct from that of MyHCeom found in extraocular muscle and of MyHCsto found in chicken Ald muscle (Figure 2).
We have performed a number of immunoblots (data not shown) with MAbs known to react specifically with intrafusal fibers, but neither of those MAbs labeled the MyHCif band or any of the other three bands. In particular, MAbs ALD19 and ALD58, specific against chicken MyHCsto, failed to label any muscle spindle band, although they reacted with extracts of chicken Ald muscle. These are intriguing results because MAbs against MyHCsto generally label bag fibers strongly and specifically (Kucera and Walro 1989; Thornell et al. 1988,1989; Pedrosa et al. 1989; Pedrosa-Domellöf and Thornell 1994; Eriksson et al. 1994). One possible explanation is that these antibodies do not detect MyHCsto on the intrafusal fibers but rather crossreact with an epitope that is affected by denaturation and which therefore cannot be demonstrated by SDS-PAGE and immunoblotting. Because the MyHCif is unique to the intrafusal fibers and so is the reactivity with slow-tonic MAbs, it is tempting to speculate that the epitope recognized by slow-tonic antibodies is likely to correspond to the MyHCif; however, further proof is needed. Because of the lack of an Ab that reacts with MyHCif both on a blot and on tissue sections and because the specificity of the ALD19 MAb against chicken MyHCsto could be confirmed, we prefer to continue interpreting the staining pattern of MAb ALD19 on human tissue sections as related to the presence of MyHCsto. Further studies of the MyHCif band will elucidate its true nature.
When sufficient spindles from the biceps or from the first lumbrical muscle were pooled together for SDS-PAGE, similar migration patterns were observed, indicating that MyHCIIa, MyHCemb, MyHCif, and MyHCI are the major isoforms present in human muscle spindles. However, it should be noted that clear differences in the relative amounts of these four isoforms are detectable by SDS-PAGE when distinct single muscle spindles or the different regions of a single muscle spindle are compared (Pedrosa-Domellöf et al. 1993; and the present study). This further supports the detailed data obtained with immunocytochemistry (see below). SDS-PAGE is not sensitive enough to detect other isoforms present in very small amounts, nor does it reflect the full complexity of the distribution of these MyHC isoforms among and along the spindle fibers. The general staining patterns of nuclear bag1, bag2, and chain fibers revealed by immunocytochemistry were very complex and are schematically illustrated in Figure 4.
MyHCsto, as interpreted by staining after incubation with MAb ALD19, was present in both bag1 and bag2 fibers along their entire length. The staining intensity of bag2 fibers with MAb ALD19 was generally as strong as that of bag1 fibers along their entire length. In rat hindlimb muscle spindles, however, bag2 fibers are typically less stained in the inner B region and unstained in the outer B and C regions (Pedrosa-Domellöf et al. 1991; Wang et al. 1997). Even in human masseter muscle spindles (Eriksson et al. 1994), the reactivity and regional variability of bag2 fibers resembles the findings in rat with respect to MyHCsto. MyHCsto was expressed in rat chain fibers for a short distance in the equatorial region (Kucera and Walro 1989; Pedrosa-Domellöf et al. 1991; Walro et al. 1997), whereas it was never found in any of the 127 chain fibers of human biceps investigated here. These findings stress the presence of species differences (Pedrosa-Domellöf et al. 1991) and the need for more data on human muscle spindles.
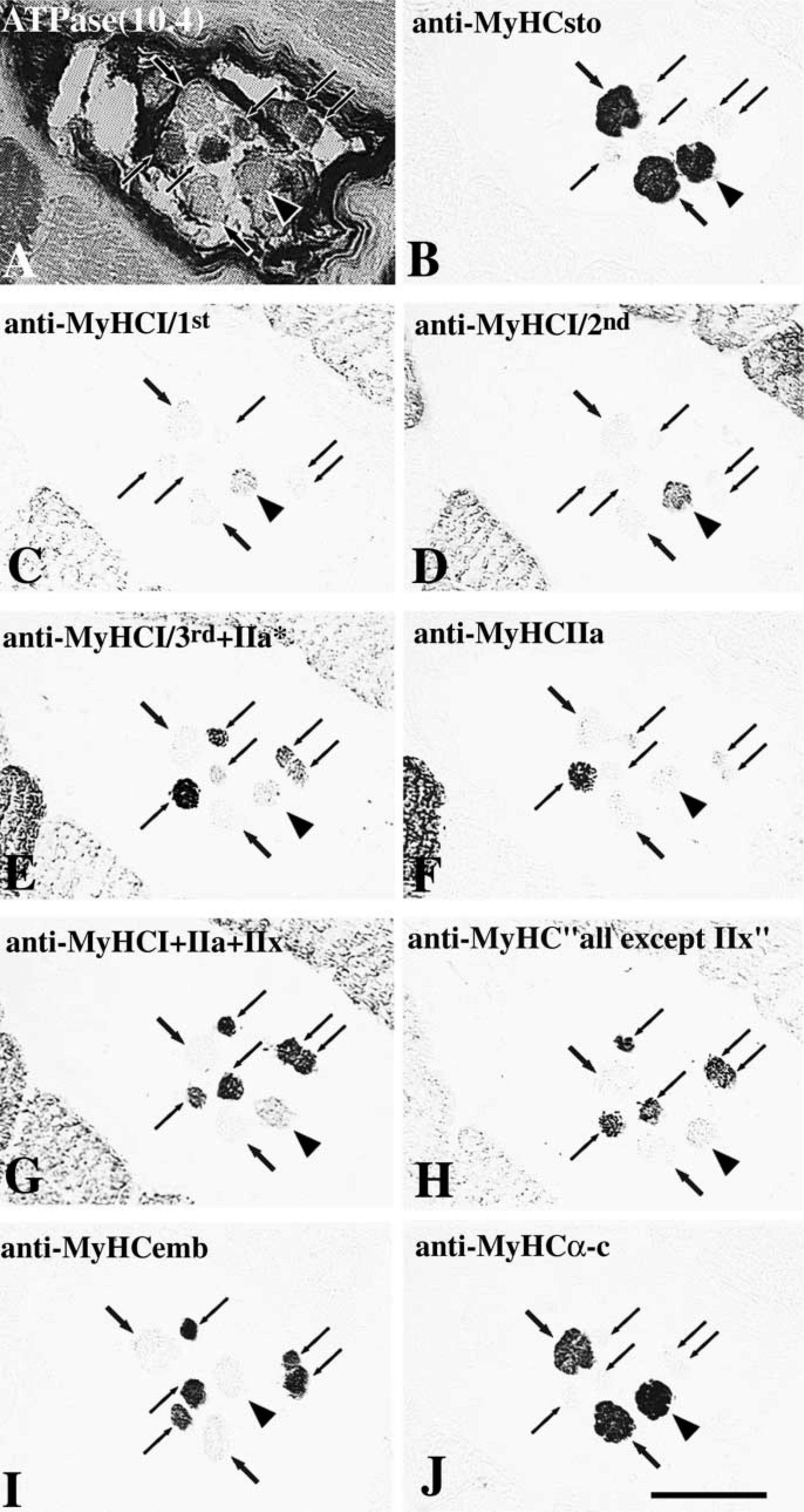
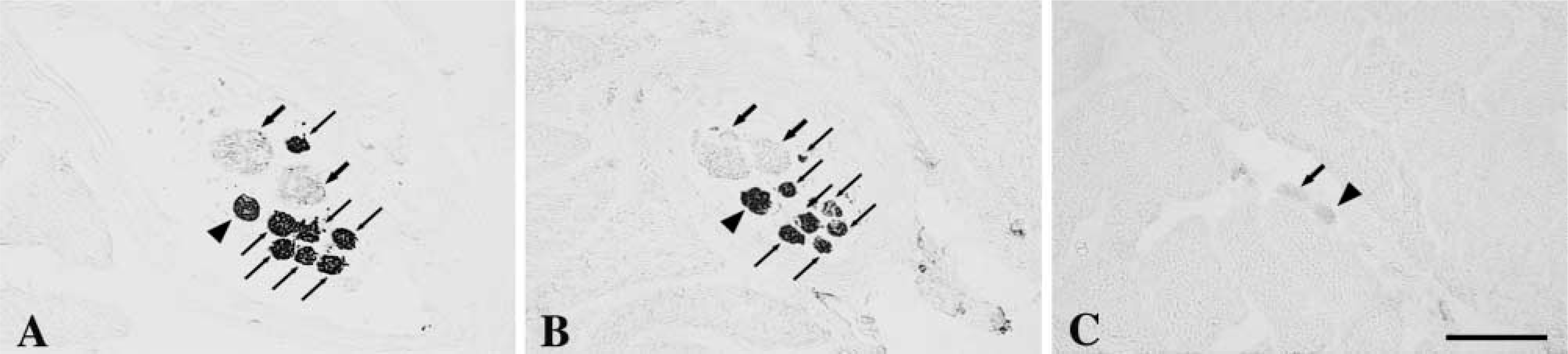
Transverse serial sections of two muscle spindles throughout the A region (
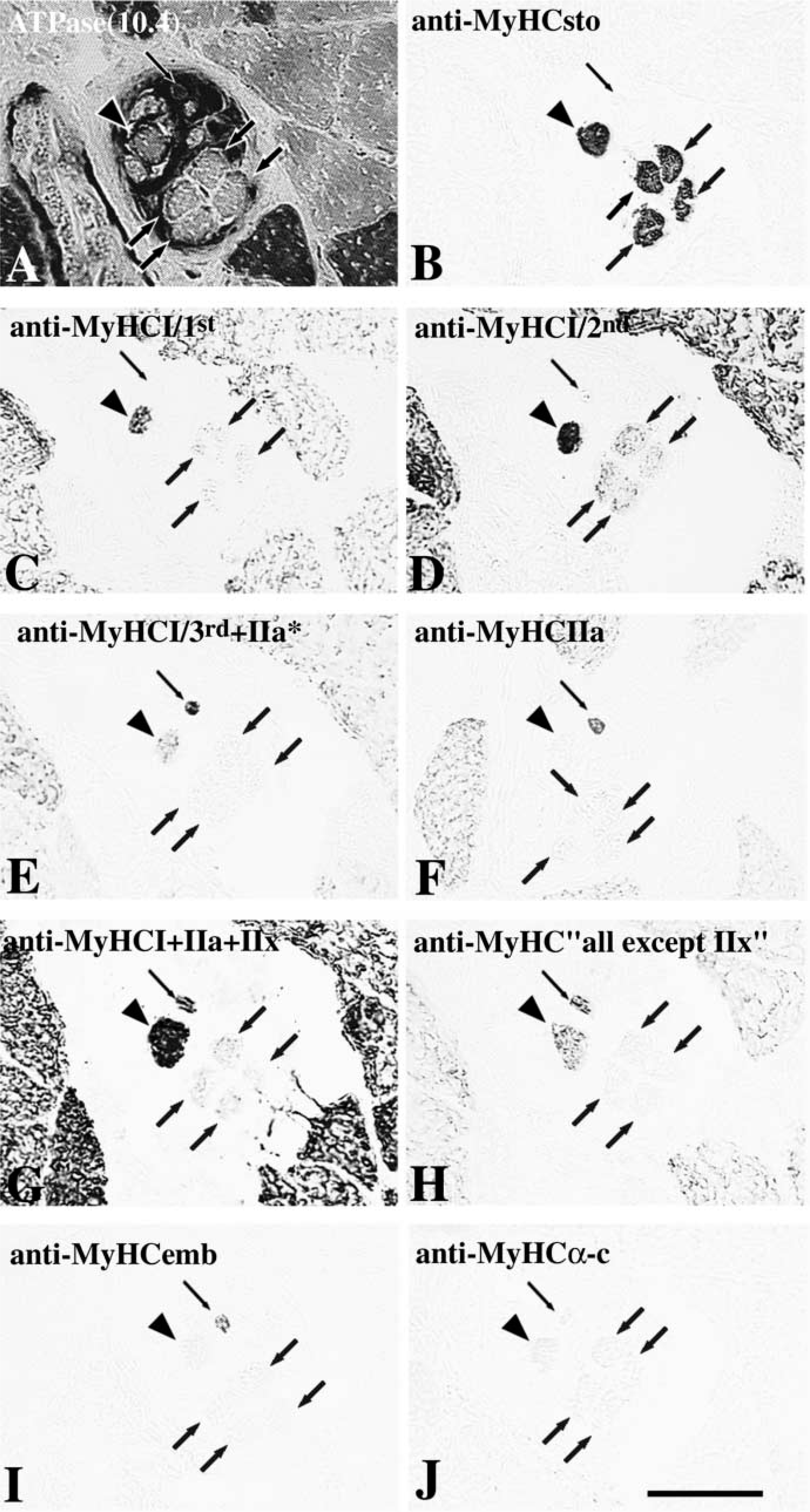
Transverse serial sections of two muscle spindles throughout the A region (
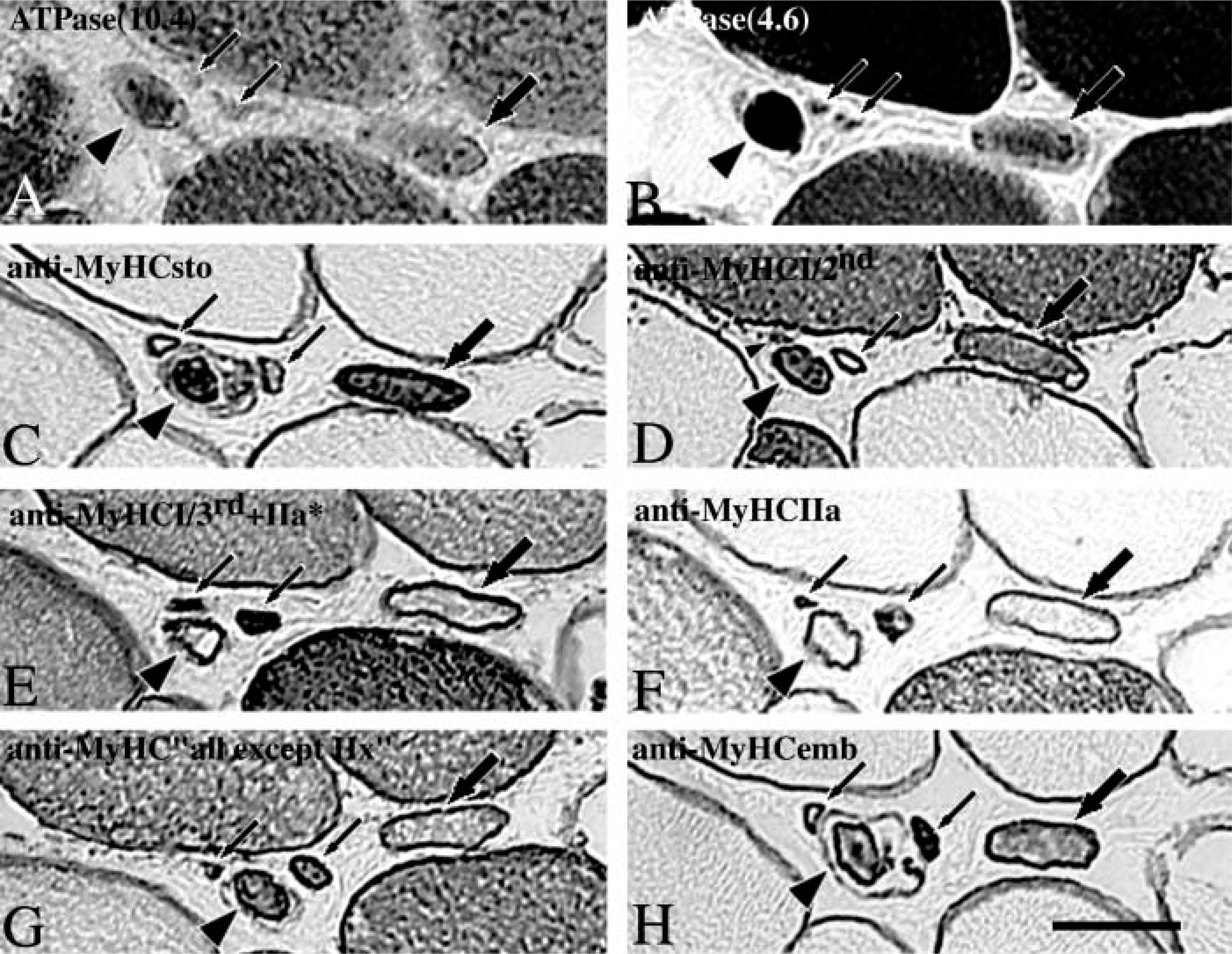
Serial cross-sections of a muscle spindle in the C region stained for mATPase at pH 10.4 (
It is obvious from the current results that nuclear chain fibers lacked MyHCsto, MyHCα-c, and MyHCI but contained MyHCIIa. Although immunocytochemistry is not a quantitative method, the binding patterns of MAb A4.74 indicate that the amount of MyHCIIa varied along the fiber length, being considerably less in the juxtaequatorial region. Therefore, in the juxtaequatorial region, chain fibers likely contained the MyHCIIx isoform according to their strong and even binding with MAb VII1H3, absent staining with the pure anti-MyHCI, and their weaker staining/unstaining with MAb A4.74. Confirmation of whether MyHCIIx was present only in the juxtaequatorial region or in other regions as well was not possible in this study because of the unavailability of a pure anti-MyHCIIx antibody. The presence of MyHCIIa and MyHCIIx in chain fibers in the present study is in contrast to reports that neither of these two MyHCs were found in rat chain fibers (Kucera et al. 1992; Walro et al. 1997; Wang et al. 1997), although rat chain fibers do contain a specific form of fast-twitch myosin (Kucera et al. 1992).
Two developmental MyHC isoforms, MyHCemb and MyHCfet, were found in all three intrafusal fiber types, although reactivity in bag fibers was lower than that in chain fibers, or might even be absent for a short distance. Maier et al. (1988) reported that chain fibers of cat spindles reacted strongly with anti-MyHCemb but that those of rat and rabbit did not react at all. In contrast, Wang et al. (1997), using a distinct anti-MyHCemb antibody, found that rat chain fibers contained MyHCemb in their intracapsular region. MyHCfet is prevalent in chain and bag2 fibers but lacking in bag1 fibers of rat muscle spindles (Kucera and Walro 1989; Pedrosa et al. 1989; Soukup et al. 1995; Wang et al. 1997). This is in contrast to our finding of MyHCfet in bag1 fibers. The present study showed that bag2 fibers contained the same types of MyHCs as bag1 fibers did but that bag2 fibers reacted to some antibodies with higher intensity and/or for a greater distance along their fiber length. In rat muscle spindles, however, bag2 fibers contained neonatal and fast-twitch MyHCs, which were absent from bag1 fibers (Kucera and Walro 1989; Pedrosa-Domellöf et al. 1993; Soukup et al. 1995; Walro et al. 1997; Wang et al. 1997), indicating that the distinctive immunocytochemical differences between bag1 and bag2 fibers are less apparent in human than in rat muscle spindles.
MyHCα-c was present in both bag1 and bag2 fibers but lacking in chain fibers, as previously reported for both human and rat muscle spindles (Pedrosa et al. 1990; Pedrosa-Domellöf and Thornell 1994; Wang et al. 1997). However, the distribution of MyHCα-c in intrafusal fibers of human biceps was distinct from that of rat, and it remains to be determined whether motor innervation regulates the expression of this isoform (Pedrosa et al. 1990) in human nuclear bag fibers as well.
During development, immunoreactivity to different anti-MyHCI antibodies (MAbs A4.840, A4.951 and N2.261) appears sequentially, suggesting that the three antibodies recognize different MyHCI epitopes in developing human and rat muscles (Hughes et al. 1993). With four distinct MAbs to MyHCI, including MAbs A4.840 and A4.951, several different phenotypes among MyHCI-containing fibers have been reported in adult rabbit masseter muscle (English et al. 1998). However, to date only one MyHCI gene has been identified, and it has generally been accepted that no alternative splicing occurs and therefore that each MyHC isoform is the product of a single gene (Weiss and Leinwand 1996). Irrespective of the possible mechanisms involved, our results further support the existence of epitope diversity for MyHCI in human muscle (Hughes et al. 1993). Significant differences in reactivity were observed with MAbs A4.951 and A4.840 in nuclear bag1 fibers. MAb A4.951 reacted with bag1 fibers in the B and C regions, whereas MAb A4.840 bound only in the C region. This is somewhat similar to the finding that MAb A4.840 reacts with a subset of the A4.951-positive Type 1 fibers in rabbit masseter muscle (English et al. 1998). The functional implications of epitope diversity have not been determined, but it can be interpreted as an additional level of complexity likely to allow very fine tuning of the contractile properties of the fibers.
Monoclonal antibody BF35 reacted strongly with MyHCsto from chicken Ald muscle in immunoblots (Figure 2), indicating that this bovine-derived antibody also recognizes MyHCsto epitopes. On immunoblots, however, MAb BF35 reacted only with the muscle spindle bands corresponding to MyHCIIa and MyHCI (Figure 2). With immunocytochemistry, no bag1 fibers were stained with MAb BF35 in the intracapsular region, and the staining seen with this MAb in the extracapsular region corresponded to the presence of MyHCI. Therefore, MyHCsto identified by BF35 MAb on immunoblots was lacking in bag1 fibers based on both immunostaining and Western blotting. Because bag1 fibers were strongly stained along their entire length with MAb ALD19 but were unreactive to MAb BF35, at least in the intracapsular region, ALD19 does not appear to recognize the same epitopes on MyHCsto as BF35 MAb does, perhaps because of their different sources (chicken/bovine). The staining seen with MAb BF35 on bag2 and chain fibers revealed the presence of MyHCemb and MyHCfet in chain fibers and MyHCI, MyHCemb, and MyHCfet in bag2 fibers rather than MyHCsto.
It is noteworthy that human spindles are far less stereotypical than spindles of small animals in terms of numbers of intrafusal fibers and their patterns of MyHC expression. Muscle spindles usually contain four or five fibers in the rat hind limb and six to nine in the cat, with no more than four nuclear bag fibers (Boyd and Smith 1984; Barker and Banks 1994). In our samples, however, we have encountered spindles containing up to a total of 16 fibers and as many as five bag fibers. A maximum of four bag fibers have previously been reported for human limb muscle spindles (Cooper and Daniel 1963). Moreover we report the presence of spindles lacking bag2 fibers. One-bag-fiber spindles have been encountered in cat (Bakker and Richmond 1981; Richmond et al. 1986; Abrahams and Richmond 1988), but they all lacked bag1 fibers except for one spindle in cat tenuissimus muscle (Kucera 1982). Most of one-bag2-fiber spindles could be traced into tandem linkages with larger spindles containing at least two nuclear bag fibers (Bakker and Richmond 1981; Richmond et al. 1986; Abrahams and Richmond 1988). The diversity seen in human muscle spindles probably reflects the higher complexity of their sensory and motor innervation (Kucera 1986), which are well-known factors influencing the expression of MyHC isoforms of intrafusal fibers (Pedrosa et al. 1989; for review see Soukup et al. 1995; Walro and Kucera 1999).
The biceps brachii muscle has a relatively ordered architecture in which the muscle fibers run parallel to the muscle's force-generating axis (Lieber and Friden 2000). This muscle has two heads, each with a different origin, and two insertions, one on the deep fascia of the forearm and the other on the radius. The combined muscle performs a variety of functions, i.e., flexes the elbow, supinates the forearm, and abducts the arm if the latter is laterally rotated. The biceps brachii has been shown to have neuromuscular compartments (Segal 1992) and, within the long head of the biceps brachii, motor units from different locations could perform different tasks (ter Haar Romeny et al. 1982). The data collected in the current study showed that virtually each muscle spindle in our samples had a different allotment of numbers of bag1, bag2, and chain fibers. In addition, the MyHC composition of intrafusal fibers was complex and variable, giving each muscle spindle its own identity.
In a rather large muscle with low muscle spindle density, such as the biceps (Peck et al. 1984), the position and function of each muscle spindle are likely to be of great importance. It is therefore not surprising that each muscle spindle presents unique morphological features. These structural variations fit with physiological data showing that each gamma-motor neuron has a response profile of its own (Johansson and Sojka 1991).
Footnotes
Acknowledgements
Supported by grants from the Medical Faculty, Umeå University, The Swedish Society for Medical Research, The Swedish Medical Research Council (12X-03934), and the Magnus Bergvall Foundation.
We thank Drs A. Kelly, D.A. Fischman, G.S. Butler-Browne, J.J. Leger, and S. Schiaffino for kindly providing antibodies. We also thank Mona Lindström and Inga Johansson for excellent technical assistance.