
Abstract
Immunogold labeling on samples of isolated perfused rat hearts embedded by an innovative low-temperature LR White procedure provided detailed insight into the interaction of caveolin-1 and endothelial NOS in myocardial capillary endothelium at the subcellular level. Separately, the localization of caveolin-1 and eNOS at caveolae under steady state conditions was visualized. A double-labeling experiment supported their close co-localization. Short-term bradykinin stimulation caused a detectable dissociation of eNOS from caveolin and its redistribution to different cell compartments, whereas caveolin itself remained stationary at caveolae. Morphometric analysis revealed that more than 80% of detectable eNOS was co-localized with caveolin-1 at caveolae under control conditions. After brief stimulation for 2 min with 10-7 M bradykinin, only 26% of the eNOS signals were associated with caveolin-1 and randomly distributed over the endothelial cells. After stimulation, eNOS was found at the plasmalemmal and intracellular membranes, freely in the cytoplasm, and at outer mitochondrial membranes.
Keywords
P
We recently explained alterations in the immunohistochemical detection of eNOS in ischemic and bradykinin-stimulated rat hearts by conformational changes of eNOS epitopes while interacting with other proteins, especially caveolin (Bloch et al. 2001). To date, no experimental approaches have been undertaken to investigate the caveolin-eNOS interaction at the subcellular level; hence, light microscopic work has been done (Segal et al. 1999).
In a variation on the post-embedding method of Berryman and Rodewald (1990), we performed a low-temperature LR White embedding procedure that delivers tissue preservation close to that of standard electron microscopy. We were able to make observations regarding the subcellular distribution of eNOS and caveolin-1 using structural caveolae in cells of intact tissue as a reference landmark, providing an independent method of protein interaction in addition to commonly applied biochemical techniques.
Rat hearts were harvested from male Wistar rats after CO2 anesthesia and cervical dislocation. After 25 min of retrograde perfusion via the aorta according to the Langendorff technique, further perfusion for 2 min with 10-7 M/liter bradykinin (Sigma; St Louis, MO) or normal solution for control was performed.
Hearts were perfusion-fixed with 4% formaldehyde and 0.2% picric acid in 0.1 M HEPES buffer (pH 7.3) plus 0.5 mM CaCl2. After 10 min hearts were stored in the same fixative at 4C for 4 hr.
Next, 25 g tannic acid powder (Merck; Darmstadt, Germany) was added to 25 ml aqua dest., heated, and stirred. The resulting clear dark-brown solution was filtered and stored as a stock solution at 4C.
After postfixation, samples were taken from the heart's left ventricle. The following steps were carried out at 4C. Without washing out the fixative, specimens were directly immersed for 4 hr in 0.1 M HEPES containing 3.5% sucrose (w/v) and 0.75% (v/v) tannic acid from the stock solution followed by buffer rinsing. Remaining free aldehydes were quenched for 1 hr with 50 mM NH4Cl in HEPES/sucrose buffer at 4C. Next, samples were transferred to 0.05 M maleate/sucrose buffer (pH 6.0 plus 0.5 mM CaCl2, 3.5% sucrose w/v). En bloc staining was done at 4C in the same buffer with 1% uranylic acetate for 4 hr.
During graded ethanol dehydration (20-min steps), the temperature was lowered to −20C at the stage of 70% ethanol. After infiltration with a 1:1 mixture of 100% ethanol and LR White and three changes with pure resin (60 min), samples were kept at −20C overnight. Then the specimens were transferred to microtubes placed in a steel block in a heat sink. Pre-cooled LR White was mixed with 15 μl LR White catalyst/ml. The activated resin was placed in the cooled microtubes, which were sealed immediately. After 12 hr, microtubes were left at room temperature (RT) for a further 48 hr. Thin sections with silver interference color were mounted on formvar-coated nickel grids.
Immunolabeling was carried out free-floating. After 15-min equilibration in 0.05 M TBS (pH 7.6), a 30-min blocking step using 3% defatted milk was performed. Polyclonal antibodies against caveolin(-1) (Transduction Labs, Lexington, KY; 1:300, cat. no. C13630) or eNOS (Biomol, Hamburg, Germany; 1:250) were applied in TBS plus 1% defatted milk for 2 hr at RT. After rinsing in TBS plus 0.05% Tween-20, grids were incubated with secondary goat anti-rabbit antibodies conjugated to 10-nm gold particles (Sigma) for 1 hr in TBS/Tween-20.
After repetitive rinsing with TBS and aqua dest., grids were transferred to 1% aqueous uranylic acetate and stained for 20 min.
For sequential double labeling, the immunoprocedure described above was done for eNOS. After a further blocking step with TBS plus 3% defatted milk to minimize artifactual cross-labeling, sections were similarly incubated with caveolin-1 antibodies, which were then coupled to 5-nm gold-conjugated secondary antibodies.
Localization of eNOS and caveolin-1
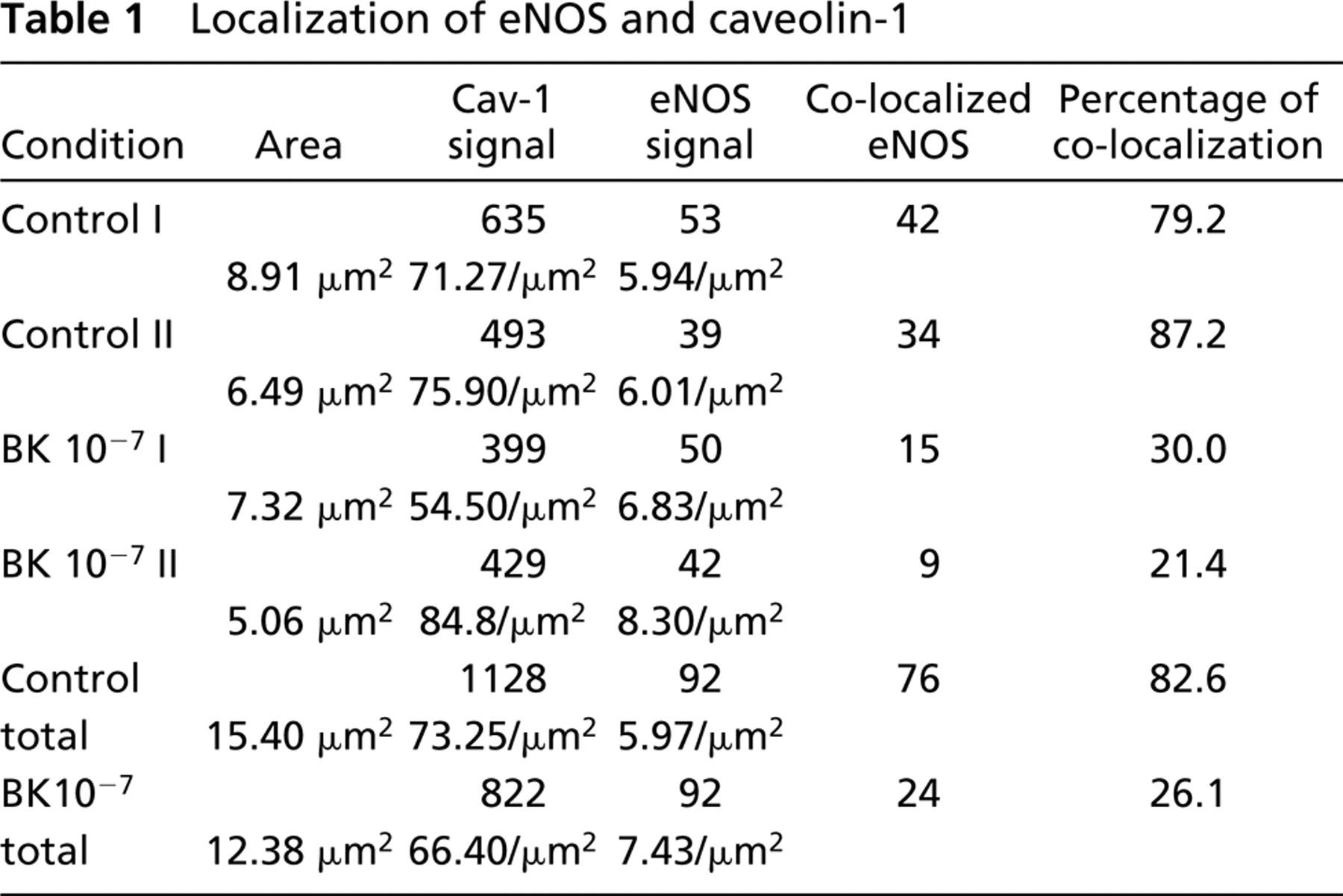
Sections were observed with a Zeiss EM 902A electron microscope at 80 kV. Images of capillary endothelium were taken at ×85,000 magnification and analyzed with a morphometric PC software (Analysis; SIS, Münster, Germany). Endothelial cells from five randomly chosen capillaries of two control and two bradykinin-treated rat heart samples were evaluated. eNOS signals (10 nm) either co-localizing with caveolin-1 (5 nm) or isolated were counted and related to the cell area (Table 1). Furthermore, co-localized signals were investigated independently by measuring the distance between two gold particles.
Light microscopic immunohistochemistry for caveolin-1 showed a strong continuous staining of the heart's microvascular endothelium. Cardiomyocytes were not labeled (not shown). This was a prerequisite to confirm the endothelial specificity of the polyclonal antibody used.
Immunoelectron microscopy revealed specific gold labeling of caveolin-1 associated with endothelial caveolae (Figure 1). Background signal was low and control incubations without the use of primary antibodies did not show any relevant signals. Myocardial caveolae were not labeled by the antibody used.
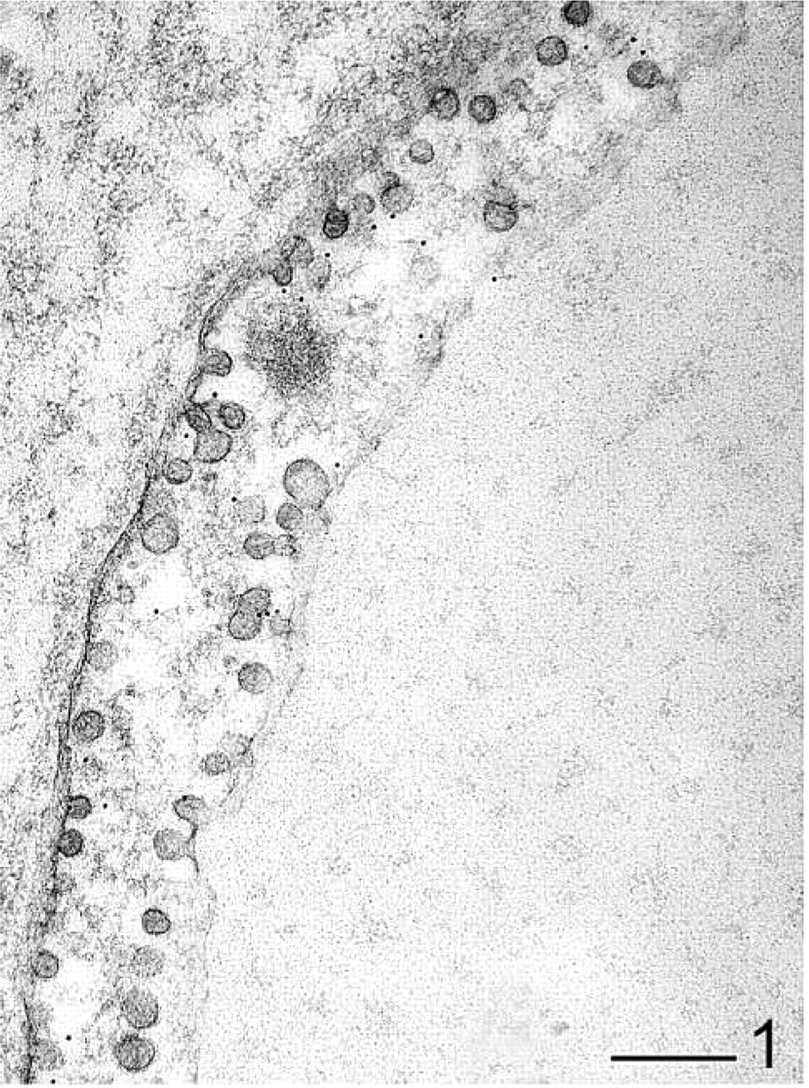
An endothelial cell from isolated perfused heart tissue. Immunolabeling for caveolin-1 is localized to caveolae. Bar = 200 nm.
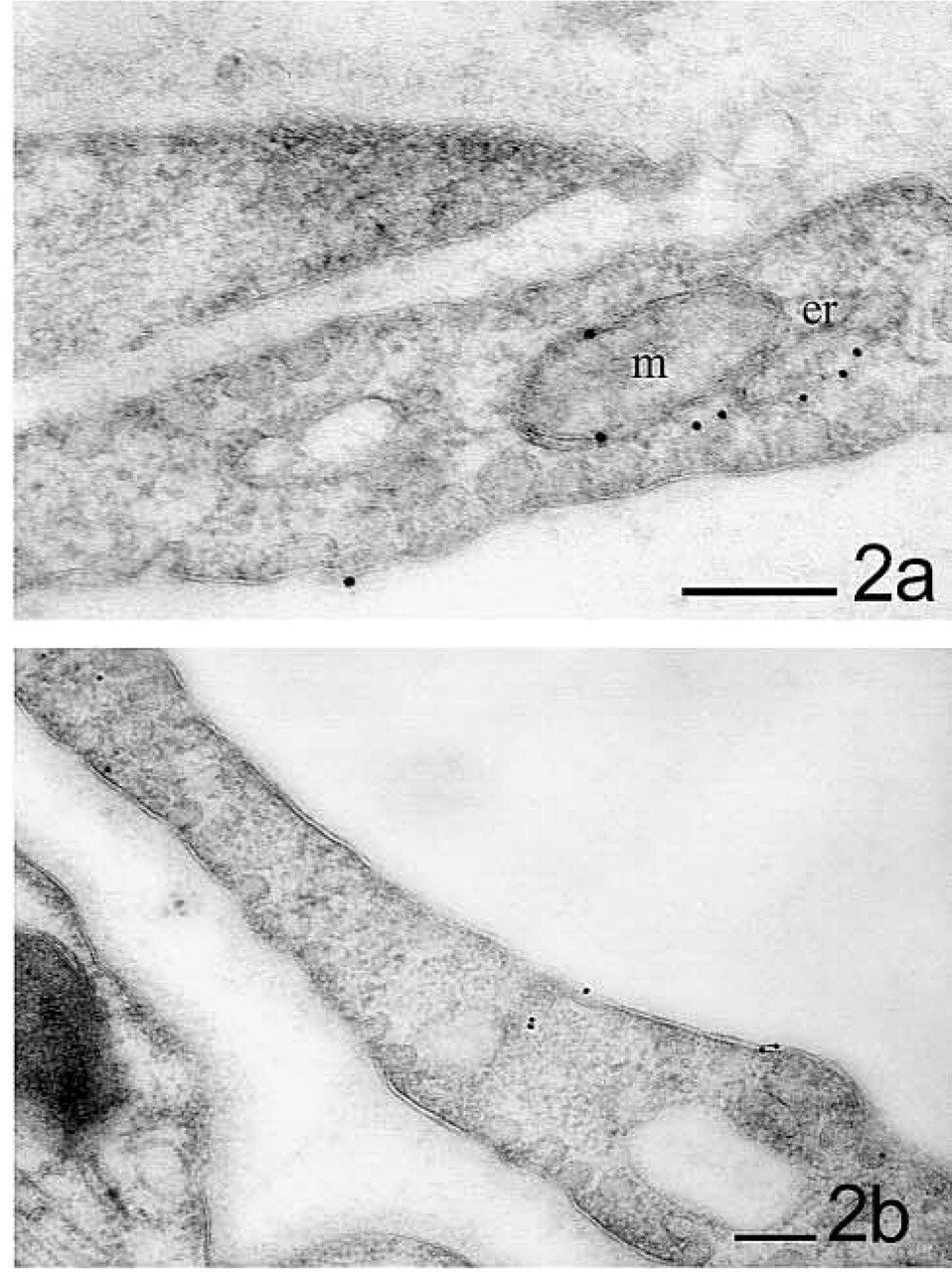
(
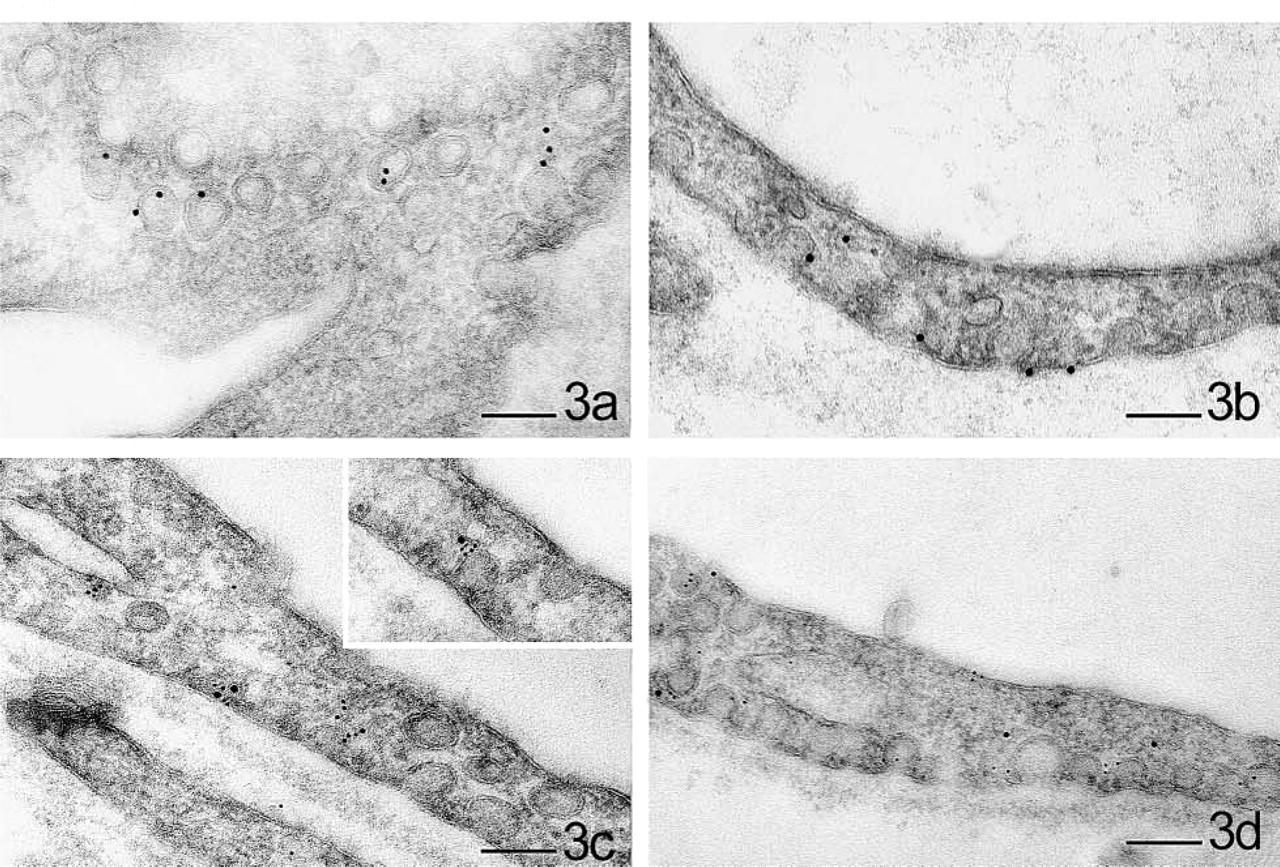
Detection of eNOS and co-localization with caveolin-1 in myocardial capillary endothelium under control and stimulated conditions. (
In unstimulated control tissue, the immunosignal for eNOS was less intense than that for caveolin-1. In the vascular endothelium it was obviously associated with caveolae (Figure 3a). After short-term stimulation with bradykinin (10-7 M/liter), the immunogoldstaining for eNOS was slightly more frequent (~24%; Table 1). It was still observable at caveolae but was mostly randomly distributed over the whole endothelium. Intracellular and outer mitochondrial membranes were also labeled, both in non-caveolar membrane sections and a solitary cytosolic signals (Figures 2a, 2b, and 3b). A preferential site for the redistribution of eNOS was not definitely determinable with the antibody used.
Figure 3 gives an overview of the sequential double labeling with caveolin-1 and eNOS antibodies under basal conditions and bradykinin stimulation. The co-localization of eNOS (10 nm) with gold particles showing caveolin-1 (5 nm) under control conditions was confirmed.
After short-term stimulation with bradykinin, the co-localization was examined (Figure 3d) and evaluated by morphometric analysis. In control samples, a mean of 82.6% of eNOS was found in the vicinity of caveolin-1, whereas only 26.1% was co-localized with caveolin after bradykinin stimulation (Table 1). Caveolin-1 was not detected in other locations than at electron microscopic-observable caveolae.
In an independent evaluation, the distance between eNOS-indicating gold particles in both groups that were associated with caveolin-1 was measured. Independently of the experimental conditions, the distance was ~15 nm in both groups (control 15.67 ± 2.90 nm; BK 10-7 14.63 ± 2.45 nm). The single distances measured were normally distributed following a Kolmogorov-Smirnov analysis.
The subcellular distribution of the membrane-associated protein caveolin-1 and its presumable interaction partner, eNOS, was demonstrated under basal and stimulated conditions by a sensitive post-embedding procedure that preserves both antigenicity and ultrastructural detail. In myocardial microvasculature endothelium, caveolin-1 is exclusively localized to caveolae. This provides further evidence for an essential role of caveolin as a “backbone” in the shaping of caveolae (Fra et al. 1995; Fujimoto et al. 2000).
The co-localization of caveolin-1 and eNOS and their functional interaction is presumed mainly on the basis of in vitro experiments (Garcia-Cardena et al. 1997; Feron et al. 1998a, b). Although the molecular interactions of caveolins and eNOS have been carefully investigated, morphological and functional examinations at the subcellular level are still lacking (Segal et al. 1999). Using the caveola of the intact cell as a reference point, we present ultrastructural evidence in an isolated organ assay that caveolin-1 acts as a functional scaffold for eNOS, which can be activated by a short-term response to kinin stimulation, as proposed in previous works (Garcia-Cardena et al. 1997; Ju et al. 1997; Michel et al. 1997). The determined spatial relation of caveolin-1 and inactive eNOS indicates a coupling of the two proteins under basal conditions, as suspected by biochemical findings. The morphometric analysis of the redistribution of eNOS from caveolin after 2 min is striking (82%:26%). This ratio might be even higher regarding the possible crossreaction of secondary antibodies due to sequential double-labeling, which leads to an artifactual co-localization of eNOS and caveolin in both groups.
In agreement with our recent findings at the light microscopic level, a slight increase of detectable eNOS after stimulation was observed at ultrastructural resolution in the present study. The subcellular translocation of active eNOS to the plasmalemma, and to the outer mitochondrial and intracellular membranes, e.g., Golgi apparatus, has been independently proposed by other groups. In the experiments presented here we could not detect a preferential location of “active” eNOS because it was found not only at the plasmalemma but also in other cell compartments, including the cytosol. Many proteins have been claimed to be associated with caveolins. It is likely that the ultrastructural visualization of their functional interaction in intact cells can be enabled by the method presented here to provide a different, more plastic insight into caveolin-related cell regulation and signaling.
Footnotes
Acknowledgments
We appreciate the excellent technical assistance provided by Ms E. Janssen and Mr Ch. Hoffmann.