Combining fluorescence in situ hybridization (FISH) and indirect immunofluorescence staining of protein markers provides a highly specific method for identifying chromosomes in phenotypically defined cells and tissues. We developed a technique enabling dual chromosome painting and immunofluorescence staining of archival formalin-fixed, paraffin-embedded material, and used this to phenotype chimeric cells in female-to-male human liver transplants.
A chronological sequence of biopsies from five female-to-male liver transplants was analyzed. Tissue sections of 5 μm were routinely prepared. Chromosome paints were prepared by degenerate oligonucleotide primer–polymerase chain reaction amplification of microdissected whole human Y chromosomes, followed by biotin–labeling by nick translation. Indirect immunofluorescence was performed using either a 1:4 dilution of a mouse monoclonal antibody against cytokeratin 7, which recognizes biliary epithelium (provided by Professor E. B. Lane, Dundee) or a 1:250 dilution of rabbit polyclonal antibody recognizing human albumin, a marker of normal liver function (Dako; Cambridgeshire, UK). Secondary antibodies were either anti-mouse or anti-rabbit Texas Red conjugates (Vector Laboratories; Orton Southgate, Peterborough, UK) used at 1:75 dilution. Sections were microwaved for 20 min in 0.1 M citrate buffer, cooled, washed in 2 × sodium saline citrate (SSC), denatured for 3 min at 75C in 70%formamide/2 × SSC, plunged into ice-cold 70% ethanol for 3 min, dehydrated through an alcohol series, and air-dried. For each slide, 150 ng labeled chromosome paint, 12 μg Cot1 DNA, and 5 μg salmon sperm DNA were denatured together for 5 min at 75C and then re-annealed at 37C for 15 min. Slides were incubated overnight at 37C in a humidified chamber. Hybridized probes were detected using avidin–fluorescein isothyocyanate (FITC) followed by biotinylated anti-avidin and a final layer of avidin–FITC (all from Vector Laboratories), washing thoroughly in 4 × SSC/0.1% Tween-20 between each step. Slides were then washed in PBS and successively incubated for 30 min with species-specific blocking serum, followed by primary and secondary antibodies, with thorough washing in PBS between incubations. Slides were then mounted in Vectashield containing 1 μg/ml 4,6-diaminidino-2-phenylidole (DAPI) counterstain (Vector) and visualised using a Zeiss Axioplan 2 fluorescence microscope equipped with a triple bandpass filter (Chroma #83000). Grayscale images were collected with a cooled charge-coupled device (CCD) camera and analyzed using custom IPLab scripts.
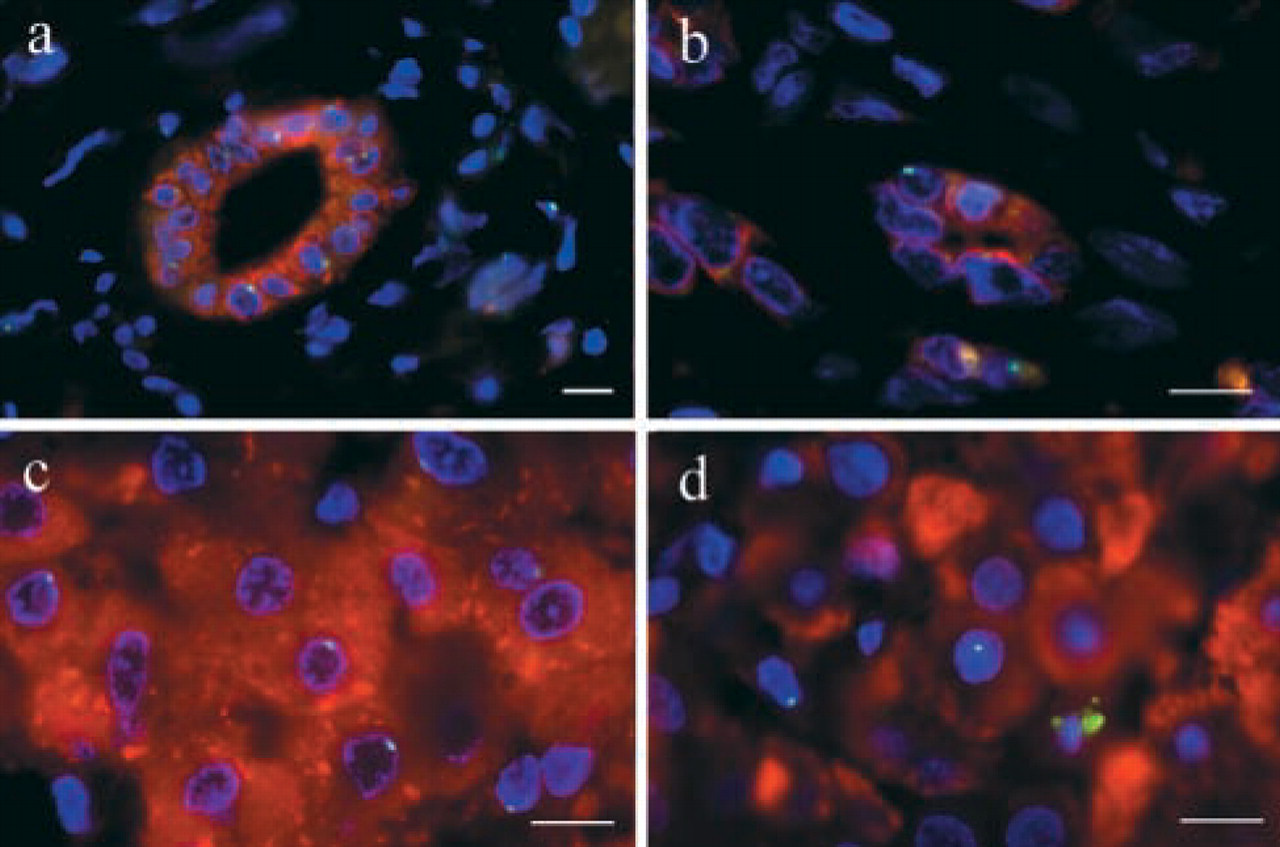
Microwaving tissue sections maintained surface and cytoplasmic immunogenicity and increased DNA probe access while causing minimal morphological disruption. The chromosome paint, which decorates both p and q arms of the Y chromosome, produced bright nuclear signals. No Y chromosome-positive cells were detected in any female donor liver before transplantation (5/5 normal histology). Y chromosome-positive cells could be detected in needle biopsies sampled 60 min after transplantation. The majority were intravascular, and none stained for cytokeratin or albumin at this time. This demonstrates that these livers were not chimeric for the Y chromosome through carriage of a male fetus (Bianchi et al. 1996; Fanning et al. 2000). Y chromosome-positive cytokeratin 7-positive epithelial cells were observed in proliferating cholangioles in an allograft infected with hepatitis B virus 10 months after transplantation (Figure 1b). These structures, which occur in response to serious liver injury, may be generated by hepatic stem-cell proliferation and differentiation (Thiese et al. 1999). Y chromosome positive albumin-positive hepatocytes were observed in female livers 1 year after transplantation into male recipients (Figure 1d). The number of Y chromosome-positive hepatocytes increased in response to liver injury to a maximum 5% of all hepatocytes. The Y chromosome-positive hepatocytes and cholangiocytes in this series of transplanted female livers must have been recipient-derived, presumably generated by circulating progenitors that infiltrated the grafts. The observation that the albumin-positive hepatocyte population in transplanted female livers may be Y chromosome-positive implies that the functional population becomes chimeric in human liver allografts.
Dual Y chromosome painting and immunofluorescence staining of liver biopsies. (a) Normal male liver displaying Y chromosome (green) and cytokeratin 7 (red) fluorescence. Bile duct epithelial cells are cytokeratin 7-positive, whereas hepatocytes are Y chromosome-positive/cytokeratin 7-negative (b). Female-to-male liver allograft displaying Y chromosome (green) and cytokeratin 7 (red) fluorescence. Note the Y chromosome-positive/cytokeratin 7-positive epithelial cell integrated into a cholangiole (only one Y chromosome signal in this focal plane). (c) Normal male liver displaying Y chromosome (green) and albumin (red) fluorescence. Thirty-three percent of hepatocytes were Y-positive in any one focal plane, increasing to >95% with 3D microscopy. (d) Female-to-male liver allograft displaying Y chromosome (green) and albumin (red) fluorescence. Note the Y chromosome-positive/albumin-positive hepatocyte integrated into the hepatic plate. Bars = 10 μm.
References
1.
BianchiDWZickwolfGKWeilGJSylvesterSDeMariaMA (1996) Male fetal progenitor cells persist in maternal blood for as long as 27 years postpartum.Proc Natl Acad Sci USA93:705–708
2.
FanningPAJohnssonJRCloustonADEdwards–SmithCBaldersonGAMacdonaldGACrawfordDHGKerlinPPowellLWPowellEE (2000) Detection of male DNA in the liver of female patients with primary biliary cirrhosis.J Hepatol33:690–695
3.
ThieseNDSaxenaRPortmanBCThungSNYeeHChiribogaLKumarACrawfordJM (1999) The canals of Hering and hepatic stem cells in humans.Hepatology30:1425–1433