
Abstract
Culture of articular chondrocytes in alginate beads offers several advantages over culture in monolayer; cells retain their phenotype for 8 months or longer. Earlier studies of chondrocytes cultured in alginate concentrated on collagen and proteoglycan synthesis. However, gene expression by in situ hybridization (ISH) has not been investigated. The purposes of the present study on human chondrocytes were (a) to modify the ISH procedure for the alginate beads to examine the mRNA expression of α1 (II) procollagen, aggrecan, and two matrix metalloproteinases (MMP-3 and MMP-8) thought to be involved in cartilage matrix degradation, and (b) to compare expression in cultured chondrocytes with that in chondrocytes of intact human cartilage. The modifications made for ISH include the presence of CaCl2 and BaCl2 in the fixation and washing steps and exclusion of cetyl pyridinium chloride. By ISH we show that aggrecan, MMP-3, and MMP-8 are continuously expressed during 8 months of culture. The α1 (II) procollagen gene is expressed only during the first 2 months of culture and after 3 months its expression is undetectable, which is consistent with its absence in adult articular cartilage. By Western blotting, Type II collagen protein had been synthesized and deposited in both the cell-associated and further-removed matrix compartments at 7 and 14 days of culture. These data indicate that chondrocytes cultured in alginate beads could be preserved for immunohistochemistry and ISH and that culture of human chondrocytes in alginate beads may serve as a good model for studying cartilage-specific phenotype as well as factors that influence cartilage matrix turnover.
Keywords
W
When bovine and human articular chondrocytes are suspended in alginate beads, these cells maintain their phenotype over long periods of time (Häuselmann et al. 1992, 1994, 1996; Mok et al. 1994; Petit et al. 1996; Binette et al. 1998). The matrix that is formed during culture is compartmentalized in a manner that may in some respects resemble that in the native cartilage (Häuselmann et al. 1994; Petit et al. 1996). As chondrocytes form the matrix of articular cartilage in vivo, they deposit a heterogeneous matrix into pericellular, territorial, and interterritorial compartments. When human articular chondrocytes are enzymatically released from that matrix and cultured in alginate beads, they reform a heterogeneous matrix with a number of similarities to those in the native tissue, including a compartmentalized matrix, the relative and absolute volumes occupied by both cells and matrix, and matrix volume per cell (Häuselmann et al. 1996). In addition, the concentration of aggrecan in the matrix associated with the chondrocyte is similar to that of adult human articular cartilage.
The purpose of this study was to extend the parameters of the alginate bead culture system to include analyses of mRNA expression by in situ hybridization (ISH) in human chondrocytes. A modification of the procedure as previously reported (Chubinskaya et al. 1996; Cole et al. 1996; Büttner et al. 1997) was required because the alginate beads dissociate in fixative and rinsing solutions unless divalent cations are present. The procedure of ISH was altered to include barium chloride, as previously reported for immunohistochemical analyses of matrix in the beads (Petit et al. 1996). The alginate was thus irreversibly crosslinked in polymer form and could then be processed by the same technique used for cartilage tissues. Expression of four genes was included in this study. The two major components of cartilage matrix, Type II collagen and the large aggregating proteoglycan, were included, as well as two members of the family of matrix metalloproteinases (MMPs) that are believed to be involved in the destruction of cartilage matrix observed in osteoarthritis. Messenger RNAs for α1 (II) procollagen IIB, aggrecan core protein, and for MMP-3 and −8 (prostromelysin and procollagenase 2) were compared in chondrocytes of newborn and adult human cartilage both in freshly fixed tissue and in separated cells cultured in alginate beads. This study demonstrates that the genes for aggrecan core protein, MMP-3, and MMP-8 are continuously expressed in human chondrocytes cultured in alginate beads for 8 months. The expression of the Type II collagen (COL2A1) gene is detectable for the first 2 months of culture but by 3 months mRNA is no longer detectable, which is consistent with its absence in adult knee articular cartilage (Aigner et al. 1997a,b; Aigner and Dudhia 1997). The Type II collagen protein is deposited in the cell-associated and further-removed matrix, as was previously reported for bovine chondrocytes. These data indicate that culture of human chondrocytes in alginate beads may serve as a good model for studying cartilage-specific phenotype and factors that influence cartilage matrix turnover and stability.
Materials and Methods
Reagents
Chemicals, either reagent or molecular biology grade, were purchased from Sigma Chemical (St Louis, MO) unless otherwise noted. Pronase was purchased from Calbiochem (La Jolla, CA), collagenase P (Clostridium histolyticum) from Boehringer Mannheim (Indianapolis, IN), and fetal bovine serum (FBS) from Hyclone (Logan, UT). Low-viscosity alginate (Keltone, LV) was a gift from Kelco (Chicago, IL). Dulbecco's modified Eagle's medium/Ham's F12 medium (DMEM/F12) was obtained in a 1/1 powder from Gibco (Grand Island, NY). All electrophoresis-grade chemicals were from Bio-Rad (Hercules, CA).
Tissue Acquisition
Full-thickness articular cartilage slices were dissected from the load-bearing regions of the tali of talocrural joints and femoral condyles of tibiofemoral joints from 10 adult human donors within 24 hr of death. Epiphyseal cartilage with diaphyseal growth plates was obtained from three newborn human donors less than 1 year old. The adult articular cartilages and epiphyseal cartilages were obtained through the Regional Organ Bank of Illinois. For the adult cartilage, only normal joints with no detectable fibrillations or osteophytes were selected.
Preparation of Chondrocytes and Alginate Beads
Chondrocytes were prepared for culture in alginate beads as previously described (Mok et al. 1994). Briefly, the cells were released from their native matrix by sequential digestion in pronase (0.2%) for 1 hr and collagenase P (0.025%) overnight in DMEM/F12 with 5% FBS at 37C. The isolated chondrocytes were suspended at a density of 4 × 106 per ml in a 1.2% solution of sterile alginate in 0.15 M NaCl. The cell suspension was slowly expressed through a 22-gauge needle and dropped into a 102-mM CaCl2 solution. The beads with approximately 40,000 cells/bead (diameter 3 mm) were allowed to polymerize for 10 min and washed twice with 0.15 M NaCl, followed by two washes in DMEM/F12. The beads were then transferred to medium (500 beads/25 ml/dish) containing DMEM/F12, 50 μg/ml gentamycin, 10% FBS, and 25 μg/ml ascorbic acid. The beads were cultured at 37C in a humidified atmosphere of 5% CO2 for up to 8 months, with medium replaced daily. The beads were transferred to new dishes every other week to avoid the formation of a monolayer on the bottom of the dish by the few chondrocytes that migrated out of the beads. There was no disruption of the beads during the culture period.
Histological Processing of Cartilage Tissue
Epiphyseal cartilage from three newborns and full-thickness articular cartilage from eight adult donors (3 × 3 mm2 pieces) were removed from the joints without subchondral bone or calcified cartilage and processed for ISH or immunohistochemistry (IHC) using RNase-free solutions as previously described (Chubinskaya et al. 1996). Sections (6 μm) were stained with Safranin O as an indicator of sulfated proteoglycans and counterstained with fast green (Rosenberg 1971). Based on the scale of Mankin et al. (1971) with a grade range of 0–13, the adult cartilage samples received histopathological grades of 0–3 and were therefore considered normal.
Processing of Alginate Beads for ISH
The beads were processed as previously described (Petit et al. 1996) except that RNase-free solutions were used. The beads were fixed in 4% paraformaldehyde, 0.1 M cacodylate buffer, pH 7.4, containing 10 mM CaCl2 for 4 hr at 20C and then washed overnight at 4C in 0.1 M cacodylate buffer (pH 7.4) containing 50 mM BaCl2. The beads were dehydrated through alcohols and xylene and embedded in paraffin. Sections (6 μm) were processed for histology (Safranin O and fast green staining), IHC, and ISH.
Oligonucleotide Probes
Probes for human aggrecan core protein [25-mer antisense oligonucleotide probe complementary to bp 736–760 (5′-ACAGAGGGACACGTCATATCGCTGT-3′)] and collagen Type IIB [23-mer antisense oligonucleotide probe complementary to bp 74–96 (5′-GCCAGGATGTCCGGCAACCAGGA-3′)] (Sandell et al. 1991) were received as a gift from Dr. L. Sandell (Washington University School of Medicine; St Louis, MO). Probes for stromelysin (MMP-3; 24-mer antisense oligonucleotide probe complementary to bp 862–885 (5′-GCCAACTGTGATCCTGCTTTGTCC-3′)] and collagenase 2 [MMP-8; 23-mer antisense oligonucleotide probe complementary to bp 1588–1610 (5′-CCCATCACTGTATCCATTCTACC-3′)] were prepared as previously described (Chubinskaya et al. 1996). All four oligonucleotide probes were 3′-end-labeled with 5′-[α-thiol-35S]-dCTP using terminal deoxynucleotidyl transferase (Gibco) and hybridized under conditions of highest stringency corrected for each probe.
In Situ Hybridization
Cartilage processing under RNAase-free conditions followed the procedure of Sandell et al. (1991) as published by our laboratory (Chubinskaya et al. 1996). The major modifications for ISH with chondrocytes in the alginate beads was the presence of divalent cations Ca+2 or Ba+2 in the fixation or buffer washing solutions, respectively, and the omission of cetyl pyridinium chloride (CPC) from the fixative. The beads were embedded in paraffin and sections were deparaffinized, rehydrated, and fixed for 5 min in 4% paraformaldehyde to crosslink the sections onto gelatin-coated slides. The sections were then treated sequentially with 0.25% acetic anhydride in 1.5% triethanolamine, 2 × SSC (sodium chloride/sodium citrate), and 100% chloroform to block nonspecific binding and remove lipid. Each oligonucleotide probe (1.5 pmol/μl) was labeled with [35S]-dCTP in the presence of terminal deoxynucleotidyl transferase (30 U) for 1.5 hr at 37C. All four radiolabeled probes were separately purified on a NENsorb column (NEN; Boston, MA) and eluted with 50% isopropanol. The radiolabeled probes were then hybridized to tissue sections in a mixture containing 25% formamide, 10% dextran, 0.3 M NaCl, 10 mM Tris, pH 8.0, 10 mM dithiothreitol, 1 mM ethylenediaminetetraacetic acid (EDTA), and 1 × Denhardt's solution. The final probe mixes each contained 4 parts hybridization mix and 1 part probe mix [2 pmol/ml labeled oligonucleotide probe, 10 mg/ml yeast tRNA, and TED (Tris–EDTA–dithiothreitol)]. The final probe mixes (60 μl/slide) were separately added to the slides and the sections were hybridized overnight at an incubation temperature calculated for each probe. The sections were extensively washed in 1 × SSC (four changes at 30 min each) to remove unbound radiolabeled probe at a washing temperature calculated separately for each probe. The sections were then rinsed through final two changes of 1 × SSC at ambient temperature. The sections were dehydrated, dried, and then dipped in liquid emulsion (NTB2; Kodak, Rochester, NY) warmed to 42C. The emulsion was allowed to dry and exposed for 3 days in total darkness. The emulsion was developed in Kodak D-19 developer and fixed in Kodak Fixer. The sections were then stained with 0.2% cresyl violet, and coverslips were applied with Acrytol (Surgipath; Richmond, IL). The sections were viewed with bright-field and darkfield microscopy to visualize reduced silver grains (Microphot-FXA; Nikon, Melville, NY). Identical camera settings and exposures were used for all photography.
Immunohistochemistry
Cartilage samples from two additional adult donors were obtained and processed for immunohistochemistry and immunoblotting after culture of the chondrocytes in alginate beads under conditions that were identical to those described for the ISH technique. Mouse anti-human Type II collagen antibody (a generous gift from Dr. T. Glant; Rush Medical College, Chicago, IL) was used as the primary antibody for indirect immunostaining. Deparaffinized sections of beads were washed for 5 min in PBS–0.2% Tween-20, followed by incubation with chondroitinase ABC (5 U/ml) in 0.1 M sodium acetate, 0.1 M Tris, pH 7.3, for 30 min at 37C. The sections were washed in PBS–0.2% Tween-20 before incubation with 80% methanol:20% of a 3% H2O2 for 20 min to block endogenous peroxidase activity. Nonspecific antibody binding sites were blocked by incubation with normal mouse sera [1:2000 dilution in PBS–0.2% Tween-20 with 3% bovine serum albumin (BSA)]. The sections were then incubated for 1 hr in primary antibody (anti-human Type II collagen) diluted 1:1500 in PBS–0.2% Tween-20–3% BSA. Control sections did not receive primary antibody; otherwise, they were treated identically. The sections were washed and incubated for 1 hr with horseradish peroxidase-conjugated goat anti-mouse IgG (Pierce Chemicals; Rock-ford, IL) diluted 1:500 in PBS–0.2% Tween-20–3% BSA. At the end of the incubation, the sections were rinsed in PBS–0.2% Tween-20 three times for 5 min each and antibody binding was visualized using one tablet of 3,3′-diaminobenzidine tetrahydrochloride and one tablet of urea hydrogen peroxide. The sections were dehydrated through alcohols and xylene and coverslips were mounted with Acrytol.
Western Blotting
The identity of Type II collagen chains in the conditioned media, cell-associated matrix, and further-removed matrix was performed using immunoblotting. The cultures were stopped at 7 and 14 days. The media were harvested every 2 days and pooled for each 7-day period. The beads were dissolved in 55 mM Na-citrate, 0.15 M NaCl, 30 mM Na2EDTA, pH 6.8, following the procedure of Mok et al. (1994). For each time point the suspension was centrifuged at 100 × g for 10 min at 4C, resulting in two fractions: the supernatant, containing macromolecules from the further-removed compartment, and a pellet, containing cells with their cell-associated matrix. The media, the cell-associated matrix, and the further-removed fractions were digested overnight at 4C with pepsin (100 μg/ml in 0.2 M NaCl, 0.5 M acetic acid). The digests were centrifuged at 12,000 × g for 30 min at 4C and the supernatants were dialyzed for 48 hr at 4C against 10 mM Tris, 1 μg/ml CHAPS, and 25 mM Na2EDTA and lyophilized overnight. The collagen in the media, cell-associated matrix, and further-removed matrix fractions was separated by SDS-PAGE (Mini-gel; Bio-Rad) in a 7% acrylamide gel (reducing conditions) according to the method of Laemmli (1970). The collagen separated by SDS-PAGE was blotted (48 mM Tris, 39 mM glycine, 20% methanol, pH 9.2) at 20 V overnight at 4C onto a nitrocellulose membrane. Nonspecific binding was blocked by incubation in PBS–5% Carnation non-fat milk and the membrane was incubated with the mouse anti-human Type II collagen antibody diluted 1:2000. After rinsing, the membrane was incubated for 1 hr at room temperature in horseradish peroxidase-conjugated goat anti-mouse IgG at a dilution of 1:500 in PBS–0.02% Tween-20–1% BSA. After rinsing, the membrane was immersed in the peroxidase substrate solution (2 ml of 4-chloro-1-naphthol, 9 ml methanol, Tris-buffered saline, and 5 μl 30% H2O2).

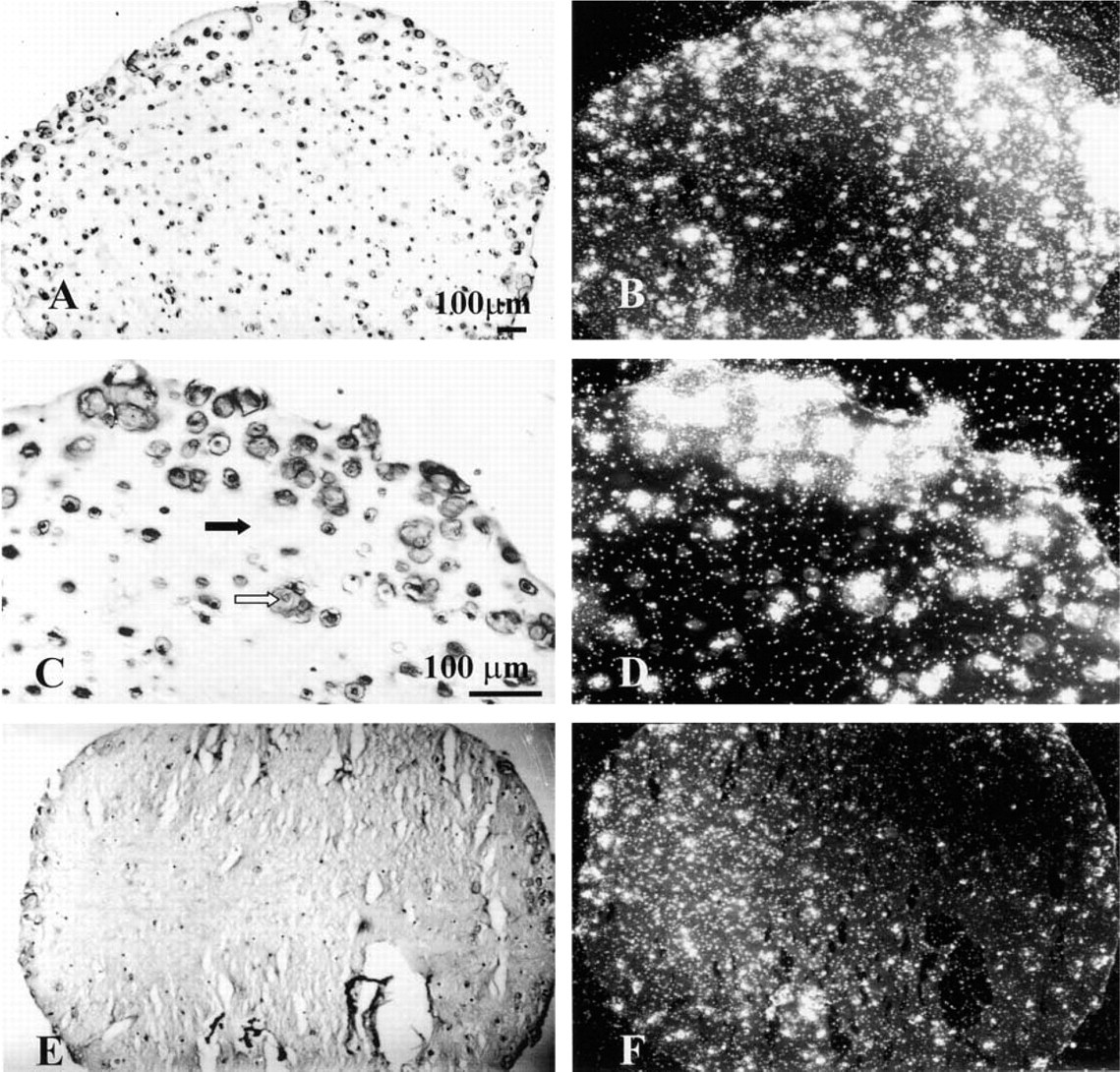
Chondrocytes from an 11-week male infant cultured for 7 days in an alginate bead. The beads were processed for histology, stained with Safranin O, and photographed with brightfield microscopy. The alginate within the bead stains more intensely than does the matrix surrounding the chondrocytes. White arrow, newly formed matrix; black arrow, alginate. Field width = 800 μm.
Results
The histological appearance of human chondrocytes cultured in alginate beads and stained with Safranin O and fast green is shown in Figure 1. In the absence of Ca+2, the alginate beads depolymerize. To maintain the structural integrity of the beads and the entrapped chondrocytes, CaCl2 was included during fixation, followed by inclusion of BaCl2 for washing to permanently crosslink the alginate. After these modified fixation steps, beads were processed for histology and the sections were stained with Safranin O and fast green (Figure 1). Both the newly formed cartilage matrix and the alginate stained with Safranin O, indicating the presence of sulfate groups. However, the alginate stained more intensely than the matrix so that the chondrocytes with newly formed matrix could be distinguished from the alginate (Figure 1: white arrow indicates the newly formed matrix; black arrow indicates the alginate). The intensity of staining in the alginate was not due to deposition of sulfated glycosaminoglycans in the further-removed matrix alone because beads formed in the absence of cells and processed for histological staining with Safranin O were comparatively intensely stained (data not shown). The histological appearance of alginate beads could be also visualized with cresyl violet (Figures 3A, 3C, 3E, 5A, and 5C). This stain is used for ISH to counterstain the matrix of the tissue or the beads. With cresyl violet the alginate was less intensely stained and the newly formed matrix was more intensely stained (Figure 3C: open arrow indicates intensely stained newly formed matrix; closed arrows indicate less intensely stained alginate matrix).

Darkfield photomicrographs of uncultured (
A second major modification made in the fixation step for ISH was the exclusion of CPC from the fixative. The presence of CPC prevented the identification of mRNA in chondrocytes cultured in alginate beads (data not shown).
Brightfield (
The comparison between mRNA expression of aggrecan core protein, α1 (II) procollagen IIB, MMP-3, and MMP-8 in the chondrocytes cultured for 7 days in alginate beads and that in fresh tissue is summarized in Table 1. In the cartilage processed for ISH without culture, mRNA expression for aggrecan, MMP-3, and MMP-8 was detectable in the chondrocytes from both adult and young donors. Type II collagen message was detectable in the chondrocytes of the young donors but not in those of the adult in the intact knee tissue (Figure 2). Messages for all four genes were detectable in both newborn and adult chondrocytes cultured in alginate beads for 7 days (Table 1) and up to 2 months (Figures 3 and 4 for Type II collagen expression). Whereas mRNA expression for aggrecan, MMP-3, and MMP-8 was detectable for an additional 6 months of chondrocyte culture in alginate (as an example, Figure 5 illustrates MMP-8 expression), message for Type II collagen became undetectable by 3 months in culture.
Relative levels of mRNA expression for aggrecan, MMP-3, MMP-8, and type II collagen in chondrocytes of freshly isolated cartilage and cartilage cultured in alginate beads for 7 daysa

Evaluation of the ISH was based on the number of grains covering the majority of the positive cells: –, negative message: no silver grains over the cells above background; [+], weak positive: less than 20 grains per cell; +, positive: more than 20 grains per cell; + +, strongly positive: individual grains too numerous to be counted.
To determine whether Type II collagen mRNA was translated into protein and deposited in the cell-associated and further-removed matrix compartments, after 7 and 14 days of culture the beads were dissolved and separated into two fractions as described in Materials and Methods. Western blotting (Figure 6) confirmed that after 7 days in culture the chondrocytes were synthesizing Type II collagen. At that time point, Type II collagen was present in the cell-associated matrix and further-removed matrix. No Type II collagen was detectable in the media. By 14 days in culture, Type II collagen was present in the media as well. Because the antibody recognizes both Types I and II collagen, Type I collagen standard was used as a negative control on the Western blots. The appearance of α2 (I) chain specific for collagen Type I would be indicative of phenotypic changes during chondrocyte cultures. As shown on the Western blot (Figure 6, α2 (I) chain marked by arrow), the α2 (I) chain that is present in the Type I collagen standard was absent from the matrix formed by the chondrocytes cultured in alginate beads.
Immunohistochemical staining of the beads with the same Type II collagen antibody used for Western blotting showed that chondrocytes with their cell-associated matrix were intensely stained at 14 days of culture (Figure 7). The further-removed matrix was stained but not as heavily as the cell-associated matrix.
Discussion
A method described for immunohistochemical evaluation of chondrocytes cultured in alginate gel (Petit et al. 1996) has been adapted for detection of mRNA by ISH. Cartilage processing under RNAase-free conditions for ISH according to the procedure of Sandell et al. (1991) has been previously published by our laboratory (Chubinskaya et al. 1996). The modifications in this method for the alginate beads primarily included the addition of Ca+2 to the fixative and Ba+2 to the subsequent rinse solutions. The Ca+2 stabilized the alginate during fixation with paraformaldehyde and the Ba+2 irreversibly crosslinked the alginate so that further processing could be done without the addition of the divalent cations. A second modification to the immunohistochemistry technique published by Petit et al. (1996) included the omission of cetyl pyridinium chloride (CPC) from the fixative. The CPC had been added to prevent loss of highly cationic proteoglycans from the matrix during processing. When CPC was included during fixation, no mRNA was detectable in the chondrocytes after ISH.
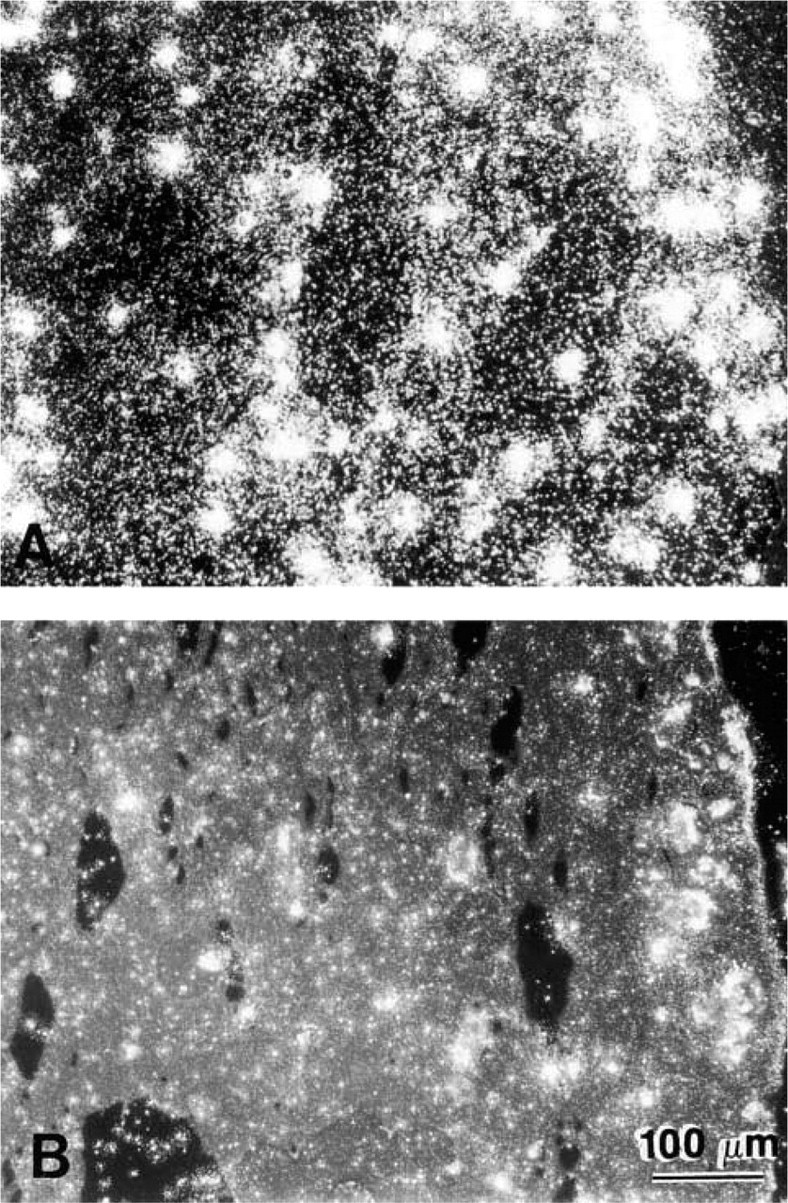
Darkfield photomicrographs of chondrocytes (38-year-old man) in alginate beads after ISH for mRNA for Type II collagen. (

Brightfield (
Applying the modifications described above, we were able to detect messenger RNA in chondrocytes cultured in alginate beads through 8 months of culture. The presence of mRNA for MMP-3 and MMP-8 in the chondrocytes of uncultured newborn and adult human cartilage is consistent with earlier studies (Chubinskaya et al. 1996) and confirms that these macro-molecules are continuously expressed within the tissue. The results of the present study demonstrate that the genes for MMP-3 and MMP-8 are also continuously expressed in chondrocytes in alginate bead culture for up to 8 months. This study further extends those observations to include the expression of aggrecan and Type II collagen.
The gene for human aggrecan core protein is continuously expressed in both developing and adult cartilage (Treilleux et al. 1992; Vornehm et al. 1996; Aigner et al. 1997b; Melching et al. 1997). Expression is upregulated in osteoarthritic cartilage compared to normal cartilage (Cs-Szabo et al. 1995, 1997). The aggrecan half-life (t1/2) has been estimated to be 2 years based on the measurement of racemization of aspartic acid in aggrecan isolated from adult human articular cartilage (Maroudas et al. 1992, 1994). In the alginate culture system, human adult chondrocytes synthesize two different pools, one with a t1/2 of 29 days in the cell-associated matrix and another in the further-removed matrix with a longer t1/2 of 100 days (Mok et al. 1994). A second pool may also exist in human cartilage in the pericellular and territorial matrices that has a t1/2 shorter than 2 years. A more rapid turnover in this pool would explain the continued expression of the aggrecan core protein gene in the adult cartilage.
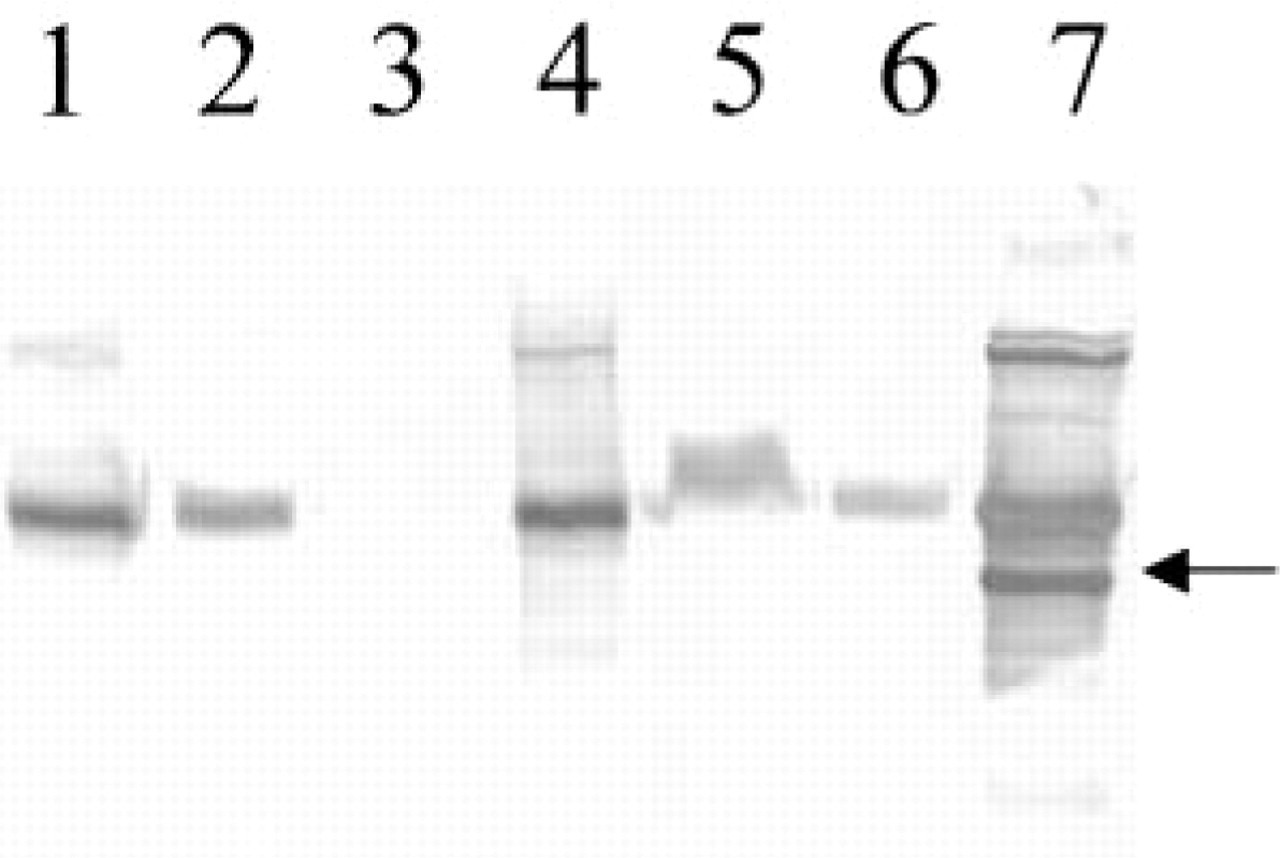
Western blotting analysis of pepsin-treated Type II collagen from adult donors in the cell-associated matrix (Lanes 1 and 4), the further-removed matrix (Lanes 2 and 5), and the media (Lanes 3 and 6) on Day 7 (Lanes 1, 2, and 3) or Day 14 of culture (Lanes 4, 5, and 6). The pepsin-treated collagen was electrophoresed in a 7% polyacrylamide reducing SDS-PAGE gel, transferred to nitrocellulose membrane, and subjected to immunoblotting with a mouse anti-human Type II collagen antibody. Lane 7 contains Type I collagen standard. Arrow points to the α2 (I) chain in collagen I standard.
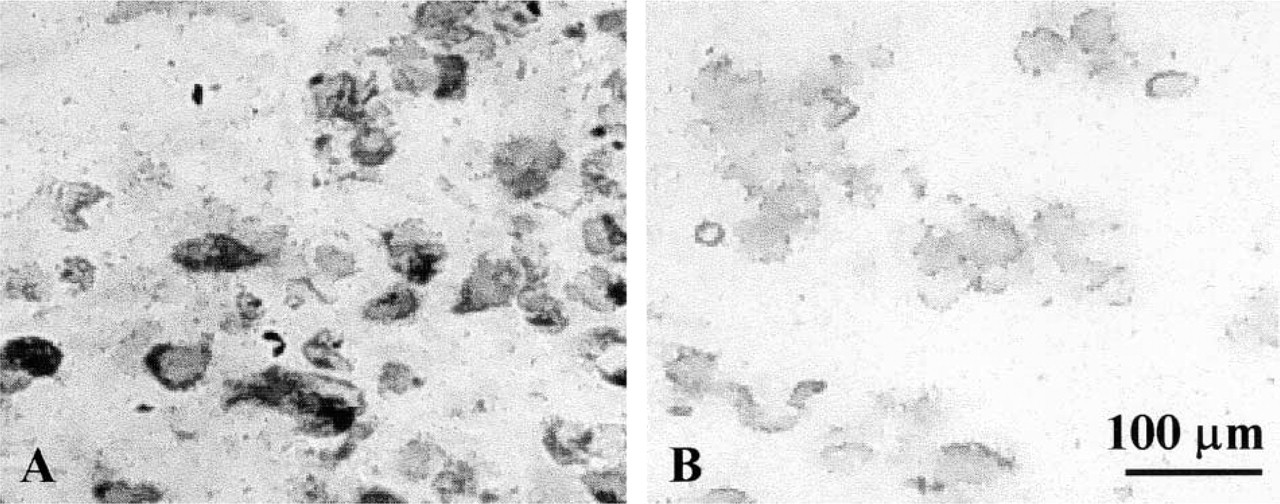
Immunohistochemical staining to detect Type II collagen protein in sections of beads after 2 weeks of culture. (
The gene for Type II collagen (COL2A1) is expressed in young, growing cartilage (Sandberg and Vuorio 1987; Reichenberger et al. 1991; Treilleux et al. 1992; Vornehm et al. 1996), with an alternatively spliced transcript Type IIB mRNA that excludes exon 2 encoding the major portion of the amino (NH2)-propeptide (Ryan and Sandell 1990; Sandell et al. 1991; Sandell 1994; Lui et al. 1995). A nonspliced variant, Type IIA, was first identified in chondroprogenitor cells in developing skeletal tissues (Sandell et al. 1991). Articular chondrocytes under normal conditions do not express Type IIA mRNA but express Type IIB message. However, during degeneration of osteoarthritic cartilage the re-expression of Type IIA collagen could occur (Aigner et al. 1999). In the adult cartilage, no expression of Type IIB mRNA is detectable unless the matrix becomes damaged, as in osteoarthritis (Aigner et al. 1992, 1997a,b; Aigner and Dudhia 1997). Maroudas et al. (1992) have estimated that the collagen network in mature cartilage turns over extremely slowly in vivo with a t1/2 >100 years. When the matrix becomes damaged in osteoarthritis, the chondrocytes again begin to synthesize Type II collagen in an attempted repair (Aigner and Dudhia 1997). The articular chondrocytes also begin to express Type X collagen (Aigner et al. 1993; Girkontaite et al. 1996) as well as Types I and III (Aigner et al. 1992).
Chondrocytes within adult cartilage have formed a network of surrounding collagen fibers in the territorial matrix. In the alginate beads, the chondrocytes, which have been stripped of their native matrix during processing, re-establish the matrix in the cell-associated and further-removed matrix compartments, similar to those previously reported for bovine chondrocytes (Petit et al. 1996). The fact that the chondrocytes no longer express the COL2A1 gene after 2 months suggests that these collagen fibers turn over slowly in the alginate beads as they do in adult articular cartilage. The alginate bead culture system appears to be suitable for studies of factors that are able to modulate not only the COL2A1 gene but also those that regulate collagen turnover, including the MMPs, especially the collagenases. Previous studies (Goldring et al. 1988, 1994; Häuselmann et al. 1993; Flechtenmacher et al. 1996) have shown that synthesis of aggrecan and Type II collagen can be either suppressed by IL-1 and INFγ or stimulated by transforming growth factor-β or osteogenic protein-1. Moreover, our recent work (unpublished data) indicates that not only normal articular chondrocytes but also osteoarthritic chondrocytes can be studied in an alginate bead system.
In this study we have shown for the first time that the alginate bead system can be analyzed for gene expression by ISH and that chondrocytes over prolonged periods of culture (up to 8 months) continue to maintain their phenotype by expressing the cartilage-specific genes.
Footnotes
Acknowledgements
Supported by NIH grant 2-P50-AR-39239, by a DAAD Fellowship (KH, Germany) and by the Deutsche Forschungsgemeinschaft (DFG Schu 1267/1–1; MS, Germany).