
Abstract
We present an up-to-date study on the nature, at the protein level, of various members of the dystrophin complex at the muscle cell membrane by comparing red and white caudal muscles from Torpedo marmorata. Our investigations involved immunodetection approaches and Western blotting analysis. We determined the presence or absence of different molecules belonging to the dystrophin family complex by analyzing their localization and molecular weight. Specific antibodies directed against dystrophin, i.e., DRP2 α-dystrobrevin, β-dystroglycan, α-syntrophin, α-, β-, γ-, and δ-sarcoglycan, and sarcospan, were used. The immunofluorescence study (confocal microscopy) showed differences in positive immunoreactions at the sarcolemmal membrane in these slow-type and fast-type skeletal muscle fibers. Protein extracts from T. marmorata red and white muscles were analyzed by Western blotting and confirmed the presence of dystrophin and associated proteins at the expected molecular weights. Differences were confirmed by comparative immunoprecipitation analysis of enriched membrane preparations with anti-β-dystroglycan polyclonal antibody. These experiments revealed clear complex or non-complex formation between members of the dystrophin system, depending on the muscle type analyzed. Differences in the potential function of these various dystrophin complexes in fast or slow muscle fibers are discussed in relation to previous data obtained in corresponding mammalian tissues.
M
Since 1987, dystrophin has been known (Hoffman et al. 1987) to be the protein that is deficient in Duchenne muscular dystrophy (DMD). This is an elongated protein present at the cytoplasmic surface of the vertebrate muscle cell membrane (Hoffman et al. 1987). Three other dystrophin-related proteins, i.e., DRP1 (dystrophin-related protein Type 1, or utrophin), DRP2 (dystrophin-related protein Type 2), and dystrobrevins, have also been identified as products of different genes (Wang et al. 1998). Dystrophins and utrophin have been detected in muscle of other mammalian species (Pons et al. 1994; Wang et al. 1998; Rafael et al. 2000) and also in other tissues, such as the electric organ, that are derived from skeletal muscle (Chang et al. 1989), and in nerves (Rivier et al. 1999a,b) of T. marmorata. Dystrophin in mammal skeletal muscle interacts with an associated protein complex (DAPC) to form a link between the cytoskeleton and the extracellular matrix (Ibraghimov-Beskrovnaya et al. 1992). This complex consists of two dystroglycans (α- and β-) (Deys et al. 1995), sarcoglycans (α-, β-, γ-, δ-, and є) that are complexed with sarcospan (Nigro et al. 1996; Crosbie et al. 1997; McNally et al. 1998), and three syntrophins (α-, β1-, β2-) in muscle tissues (Ahn et al. 1996). However, new isoforms of syntrophins (γ1- and γ2-) have also been reported and were found expressed as brain-specific protein (Piluso et al. 2000). Deficiency or variations in some associated proteins generate a different muscle pathology, but the pathogenesis of all of these related muscular dystrophies is still unclear. This may be due to the heterogeneity of the data recorded, e.g., due to the existence of four dystrophin-related proteins that all share homology with dystrophin's cysteine-rich and C-terminal domains and also because of the muscle type analyzed.
In muscles, dystrophin expression is believed to provide resistance of muscle membrane during contraction processes (Pasternak et al. 1995). Thin and thick myofilament sliding leads to muscle sarcomere contraction while forces are laterally transmitted to the extracellular matrix via the dystrophin–glycoprotein complex (DGC). In dystrophin–deficient muscle, the absence of this complex causes sarcolemmal instability, which renders muscle fibers susceptible to necrosis (Menke and Jockusch 1995). Then the DGC is thought to play a mechanical and structural function in muscle, but recent findings also suggest a role in cell communication due to associations with a variety of signaling molecules (Cohn and Campbell 2000).
This work investigated the possible presence and distribution of dystrophin family products and some dystrophin-associated-like proteins in red and white muscles from T. marmorata. This approach was possible using a large battery of specific monoclonal and polyclonal antibodies produced by us that were authenticated in previous studies as specific for dystrophin, utrophin, or associated proteins (Fabbrizio et al. 1993, 1994; Rivier et al. 1996, 1999a). After a first screening to select antibodies that would give clear immunofluorescent labeling in the muscle cell membranes studied, we determined the presence and size of the molecules detected using Western blotting technique. Both confocal analyses and immunoprecipitation approaches helped us to determine whether these proteins were differently distributed in fast- and slow-type skeletal muscles. Possible functional implications are discussed.
Materials and Methods
Tissues
Transversely striated muscles (red or white) from live T. marmorata fish were dissected and quickly frozen for indirect cytochemical fluorescence labeling or were treated for crude protein extraction.
Antibodies
We used various antibodies specific for various proteins of the dystrophin family. In detail, monoclonal (5G5, 3E7, 5A3, 4A1,) and polyclonal (C) antibodies were directed against dystrophin and/or utrophin molecules. The specificities of all monoclonal and polyclonal antibodies were previously confirmed in muscle and nerve tissues of rabbit (Fabbrizio et al. 1993), Torpedo (Rivier et al. 1996), and adult bovine heart (Rivier et al. 1999a, 1999b). The antigenic sequences, corresponding to the recombinant C-protein that was prepared from a dystrophin fragment derived from the chicken skeletal muscle gene (residues 1173–1728), gave rise to polyclonal antibodies (C as referred to in Pons et al. 1990). We used polyclonal antibodies directed against DRP2, α-dystrobrevin, and sarcospan that were obtained by injecting their specific C-terminal synthetic peptides (ANTLLAS), (GVSYVPYCRS), and (SLTAESEGPQQKI) as antigens, respectively, according to a previously described protocol (Pons et al. 1994). In this study, we also used other polyclonal antibodies directed specifically against sarcoglycans (α, residues PLILDQH; β, residues AGYIPIDEDRL; γ, residues VREQYTTAT; δ, recombinant protein residues 84–290) and β-dystroglycan (residues PPPYVPP). All antibodies were previously characterized in adult bovine heart (Rivier et al. 1999a,b), in a Becker dystrophy patient (Rivier et al. 2000), and in other works related to annelid muscles (Royuela et al. 1999). The polyclonal antibody against α-syntrophin (antigenic residues 191–206) was recently detected in Torpedo (Nigro et al. 1996). Finally, we used the 13E2 monoclonal antibody that we previously obtained from a crude extract of T. marmorata electric organ (as reported in Rivier et al. 1996) directed against a 90-kD protein rather than against a 400-kD protein. This 13E2 antibody was of interest mainly because of the apparent molecular weight of the protein detected in crude extract obtained from Torpedo electric organ, which corresponded to the Ao protein reported by Japanese authors as belonging to the dystrophin complex (Yoshida and Ozawa 1990). Monoclonal antibodies against α-, β-, γ-, and δ-sarcoglycans were obtained from Novocastra (Sigma; St Louis, MO).
The specificity of the immunochemical procedures was checked by incubation of sections with non-immune serum instead of primary antibody. As positive controls for this study, skeletal muscle sections of mouse (for polyclonal antibodies) or rabbit (for monoclonal antibodies) were treated simultaneously with the Torpedo muscles.
Western Blotting Analysis
Tissues were homogenized in the extraction buffer (0.05 M Tris-HCl, pH 8) with the addition of a cocktail of protease inhibitors (100 mM iodoacetamide, 0.1 mM phenylmethyl sulfonic fluoride, 0.01 mg/ml soybean trypsin inhibitor, 1 μl/ml leupeptin) in the presence of 1% Triton X-100. Homogenates were centrifuged for 10 min at 10,000 rpm. Supernatants and pellet (resuspended in 50 mM Tris-HCl, pH 8) were mixed with an equivalent volume of SDS buffer (10% SDS in Tris-HCl, pH 8, containing 50% glycerol, 0.1 mM 2-β-mercaptoetanol, and 0.1% bromophenol blue). The mixture was denatured for 5 min at 100C and 10-μl aliquots of homogenate were separated in SDS-polyacrylamide slab minigels (3–12% gradient gels). To carefully compare protein extraction, we equilibrated each sample by equilibrating the muscle actin content. Separated proteins were transferred overnight (30 V, 100 mA) in the transfer buffer (25 mM Tris-HCl, 192 mM glycine, 0.1% SDS, and 20% methanol). Nitrocellulose membranes (0.2 μm) were blocked with 3% BSA dissolved in TBST buffer (10 mM Tris-HCl, 150 mM NaCl, 0.05% Tween-20, pH 8) for 1 hr with specific antibodies and then were incubated with a phosphatase-labeled second antibody (1:3000 dilution; Jackson ImmunoResearch Laboratory, West Chester, PA). The protein band was visualized with p-nitroblue tetrazolium, and 5-bromo-4-chloro-3-indoylphosphate substrate, as previously described (Fabbrizio et al. 1994).
Scanning Densitometry
Western blots were digitized with a 256 gray scale and images were quantitatively analyzed using the NIH 1.62 image program. Each lane was treated independently and corrected by estimation of the relative optical density found in the Coomassie blue gel corresponding to the amount of actin present in the W or R lane. Each blot was obtained in triplicate to avoid errors due to differences in signal intensities. Values were normalized and averages for the three assays were compared between white and red muscles. Their ratio was expressed as a percentage of each component found in red muscle vs white muscle.
Immunoprecipitation
Anti-β-dystroglycan polyclonal antibody was used as in Araishi et al. (1999) for immunoprecipitation. White or red muscle (0.5 g) from Torpedo was homogenized and solubilized in 2 vol. (w/v) of extraction buffer (50 mM Tris-HCl, pH 9, 100 mM iodoacetamide, 0.1 mM phenylmethyl sulfonic fluoride, 0.01 mg/ml soybean trypsin inhibitor, 1 μl/ml leupeptin, 1% Triton X-100, and 1% saponin). After centrifugation at 140,000 × g for 45 min at 4C, the resulting supernatant, corresponding to the enriched muscle membrane preparation, was incubated overnight at 4C with anti-β-dystroglycan polyclonal antibody. The mixture was applied on a prepacked protein A–Sepharose column 1 × 4 cm (Amersham Pharmacia Biotech; Arlington Hts, IL). Resin was recycled with this antigen–antibody solution for 2 hr at 4C and then washed extensively with extraction buffer containing 0.5 M NaCl, until no protein was detected at 280 nm OD. Total elution was obtained after adding 10% SDS solution and aliquots were immediately treated for protein transfer. The resulting nitrocellulose sheets were analyzed using the first specific antibody relative to the component that we intended to detect.
Immunofluorescence Light Microscopy
Cryostat sections (10 μm) of unfixed muscle were labeled with different specific antibodies. Immunoreactions were detected with Cy3-conjugated sheep anti-mouse Ig (for monoclonal antibodies) or Cy3-conjugated sheep anti-rabbit IgG (for polyclonal antibodies). For double detection, sections were first labeled with the monoclonal antibody concomitantly with a polyclonal antibody (against dystrophin-related protein or DAP). In a second step, the monoclonal antibody was revealed with Cy3-conjugated sheep anti-mouse IgG (1:500) and fluorescein-conjugated goat anti-rabbit IgG (1:100) for all other polyclonal antibodies (Sigma Bio-Sciences Laboratory; St Louis, MO).
Confocal Laser Microscopy
Confocal laser microscopy analyses were performed as described in previous studies (Rivier et al. 1999b, 2000), using a Bio-Rad MRC-1024 apparatus mounted on an Optiphot II Nikon fluorescence microscope equipped with a ×60 lens (Plan apo; NA 1.4). The two signals were processed using the COMOS software program (BioRad, Hercules, CA). Images were obtained after processing through the Adobe Photoshop (version 3.0) software program and were printed on an Epson Stylus Color 800 printer.
Results
Immunofluorescence Analyses
Immunofluorescence reactions were studied through two concomitant investigations and are presented comparatively in two figures, i.e., Figures 1A-1J (white muscle) and Figures 2A-2J (red muscle). It was always clear that the large fibers corresponded to white muscle (fast-type), whereas small fibers were found only in red muscle (slow-type) (see Figure 1 and 2). For these experiments, as later in Western blotting assays, components such as dystrophin and 90-kD protein were viewed with monoclonal antibodies 5A3 and 13E2, respectively, while all other proteins were identified with polyclonal antibodies that we produced. Torpedo dystrophin detection was found (as expected) at the muscle cell membrane with each of the specific antibodies used in this study (Figures 1A and 2A, left panels) and this presentation is repeated in each following image (left panel), excluding Figures 1E and 2E, left panels. The two other dystrophin family proteins studied, i.e., DRP2 and α-dystrobrevin, are presented in Figures 1A, 1B, 2A, and 2B, middle panels, and were not found at the muscle cell periphery of fast-type fibers (Figure 1) but rather at the membrane of slow-type fibers (Figure 2). β-Dystroglycan, which is presented in Figures 1C, 1E, 2C, and 2E, middle panels, was detected in both fast- and slow-type fibers. α-syntrophin Figures 1D and 2D, central portion, was located in fast-type fibers (Figure 1) but was absent in cell membranes of slow-type fibers (Figure 2). Similarly, in Figures 1E and 2E, left panels, a specific monoclonal antibody against a 90-kD protein (as described in Materials and Methods) was clearly detected in the membrane of fast-type fibers (Figure 1) co-localized with β-dystroglycan but was not present in cell membrane structures of slow-type fibers (Figure 2). In the following set of experiments, we analyzed polyclonal antibodies that we had developed and characterized as specific for sarcoglycan family products. We also obtained a good correlation between results with polyclonal antibodies and similar commercially available specific monoclonal antibodies, in their capacity to detect, at the membrane or not, one of these sarcoglycans. However, only a commercial monoclonal antibody against α-sarcoglycan was undetectable in both fiber types, in contrast to our polyclonal antibody produced against this protein, which was efficient. In fast fibers (Figure 1), the following differences appeared in the middle panel: α-sarcoglycan (Figure 1F), γ-sarcoglycan (Figure 1H), and sarcospan (Figure 1J) were all present at the white muscle membrane. In contrast, β-sarcoglycan (Figure 1G) and δ-sarcoglycan (Figure 1I) were not detected at the membrane of these fast-type fibers (Figure 1). In slow fibers (Figure 2), all proteins detected Figures 2F-2J (middle panels, corresponding respectively to α-, β-, γ-, δ-sarcoglycan, and sarcospan) were found at the cell membrane.
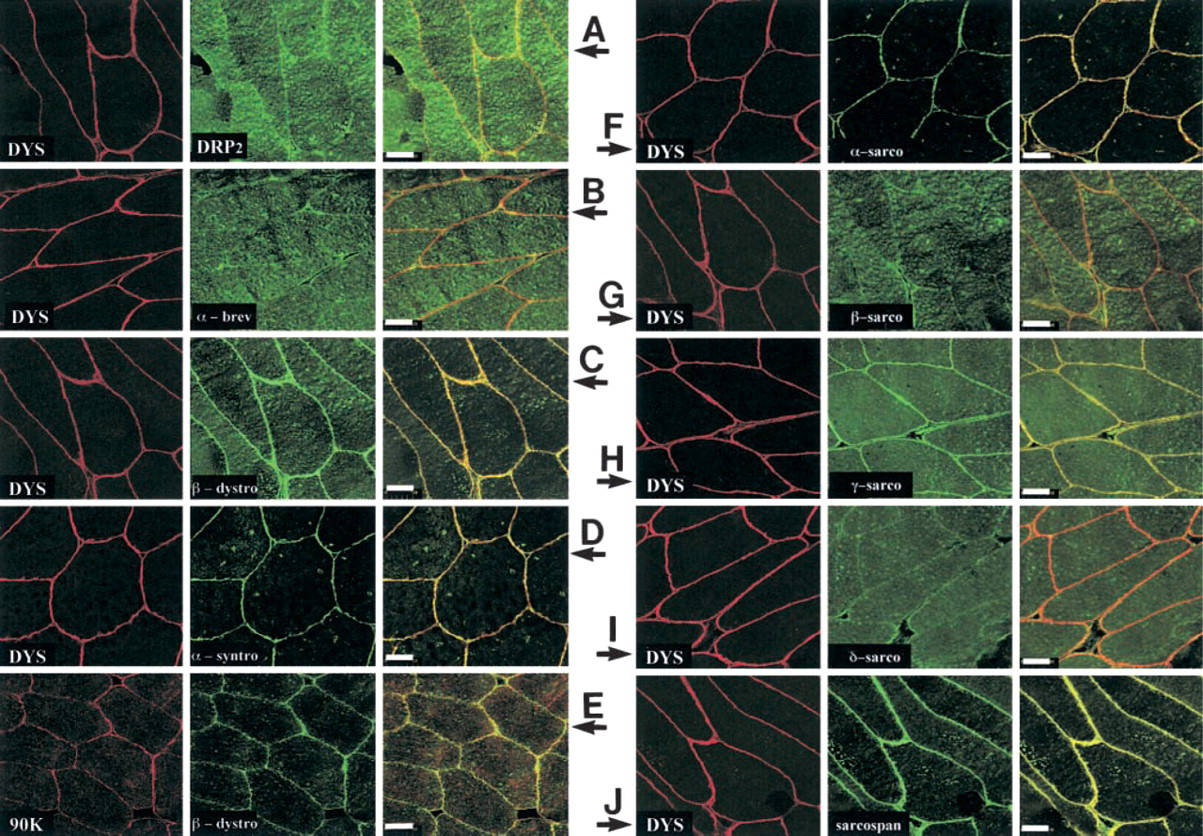
Confocal analyses of fast-type muscle fibers from T. marmorata with different antibodies related to dystrophin family products or dystrophin-associated proteins. The antibodies used in these experiments were monoclonal for dystrophin (5A3) and the 90-kD protein (13E2) and polyclonal for all other components studied. White muscle cross-sections (10 μm) from Torpedo were viewed using antibodies directed against different molecules belonging to dystrophin family products or associated proteins in reference to vertebrate muscle. (
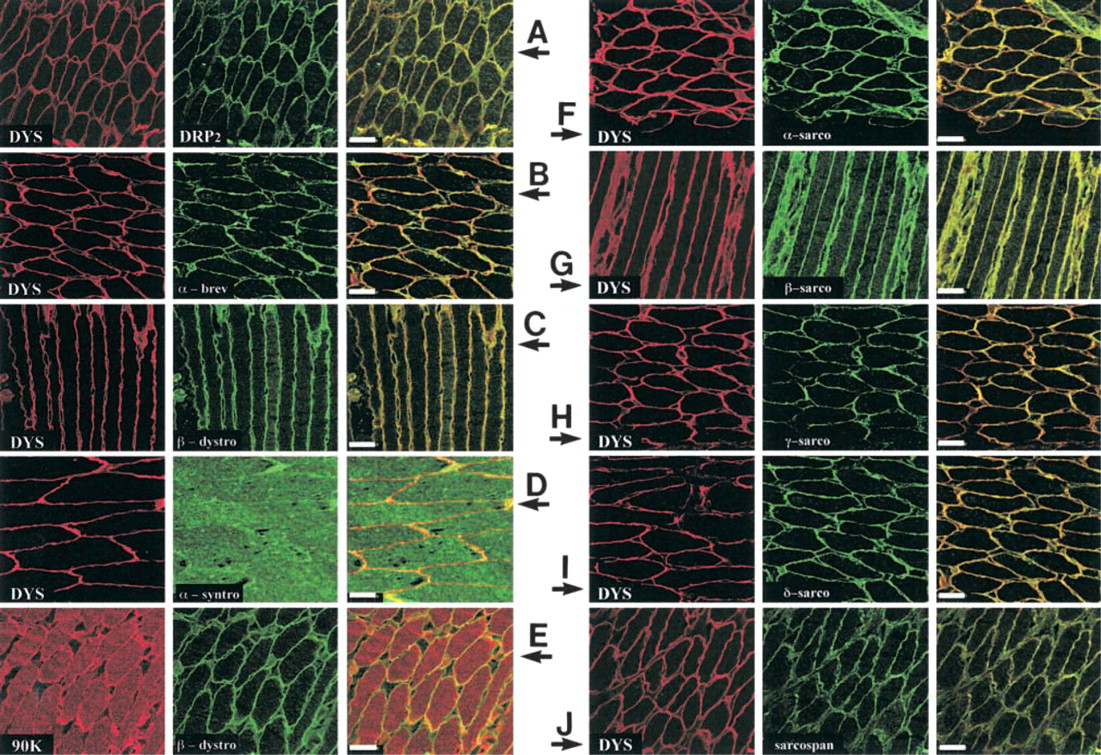
Confocal analyses of slow-type muscle fibers from T. marmorata with different antibodies related to dystrophin family products or dystrophin-associated proteins. As in Figure 1, comparative analyses were undertaken on red muscle fibers. Cross-sections (10 μm) were viewed using antibodies against different molecules belonging to dystrophin family products or associated proteins in reference to vertebrate muscle. (
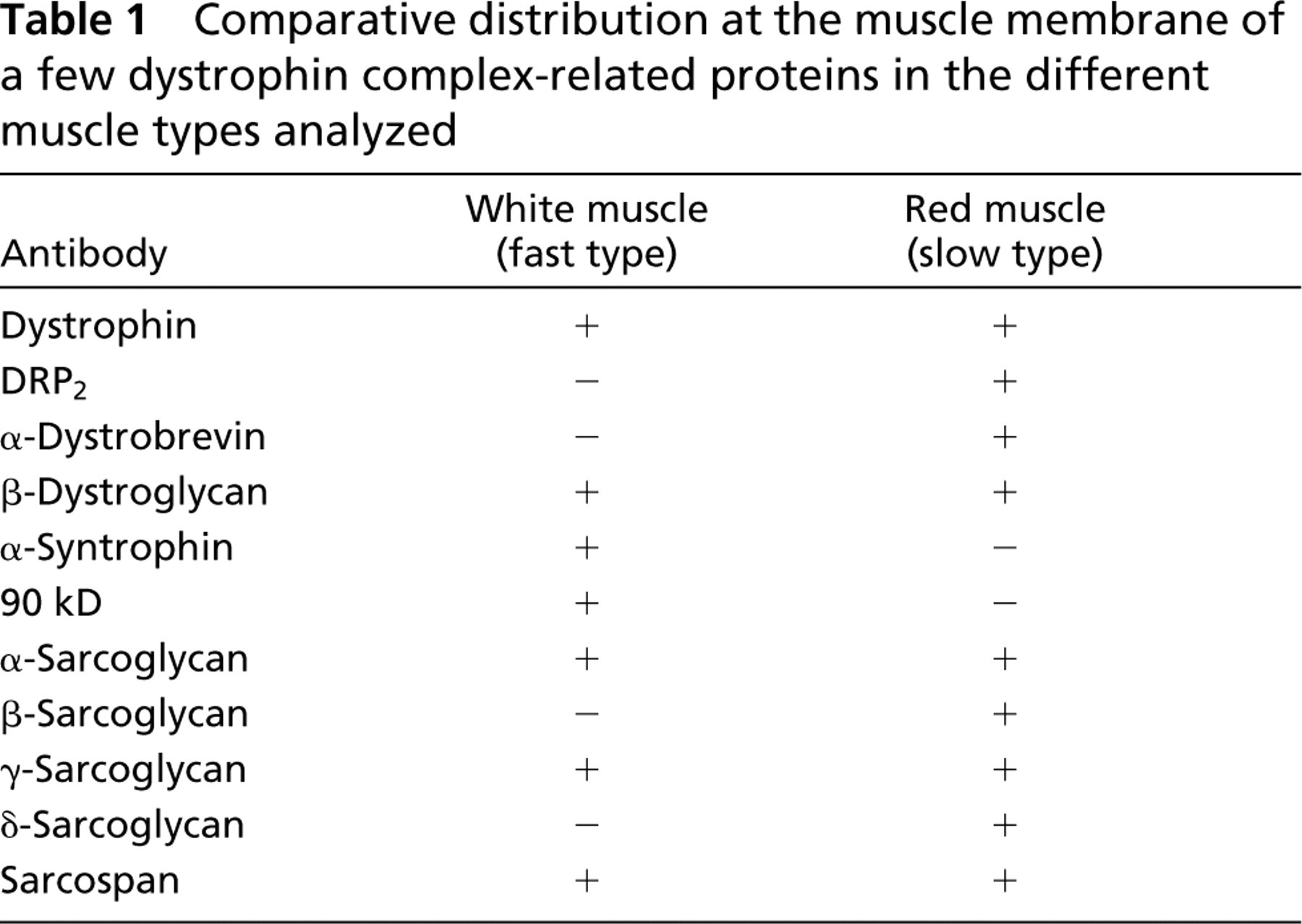
These confocal microscopic observations are illustrated in the right panel of each set, corresponding to the superimposed images, which gave a clear yellow color when there was co-localization but only a pale orange color when the associated protein was not found at the muscle cell membrane. These results are summarized in Table 1, in which we comparatively present the membrane distribution in white muscle (fast type) and red muscle (slow type) for each member of the dystrophin complex.
To obtain more information in these comparative immunodetections, we confirmed the presence of the molecule analyzed in crude white (W) and red (R) muscle extracts.
Western Blotting Analyses
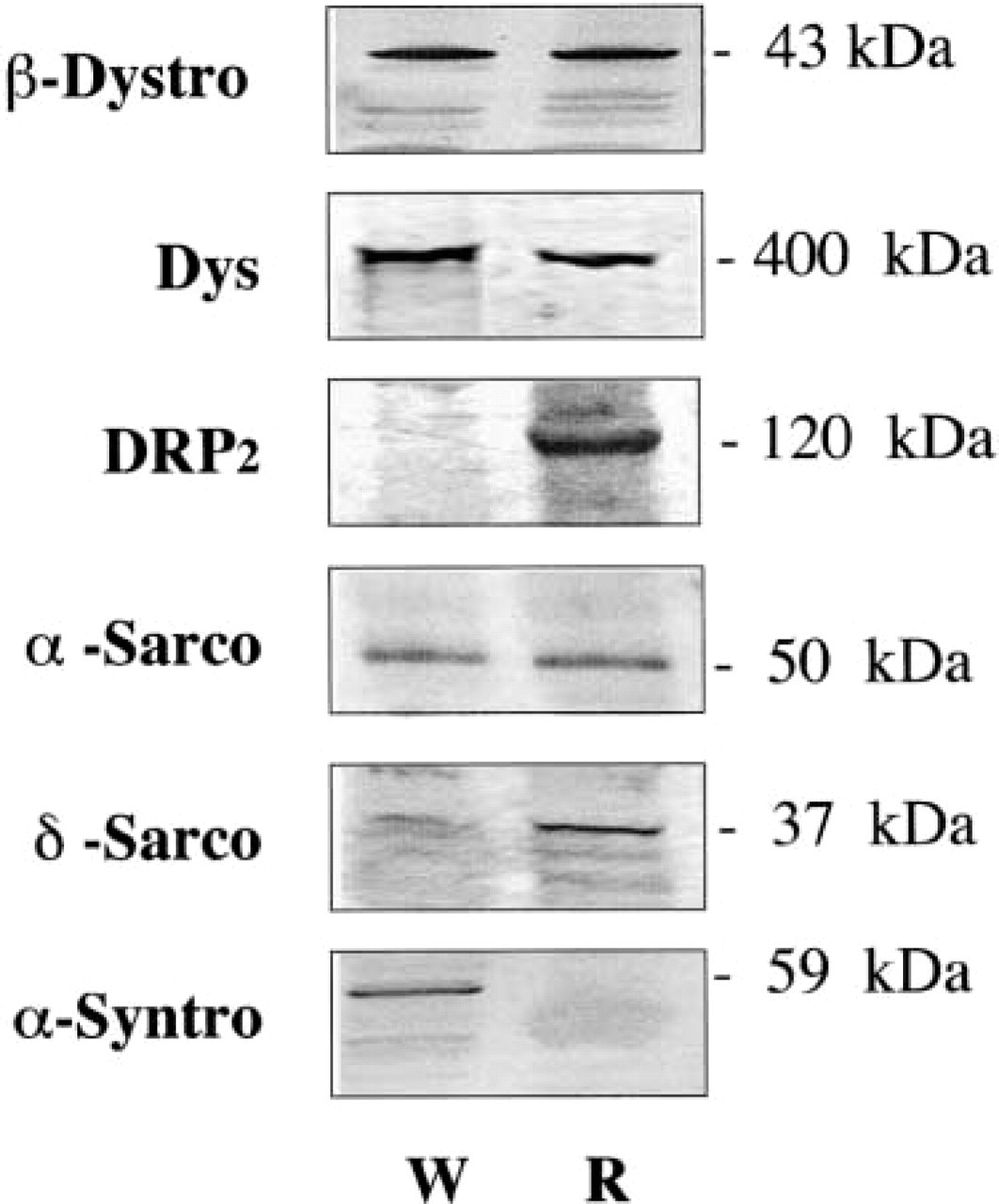
Crude protein extracts from T. marmorata slow-type and fast-type fibers were transferred to nitrocellulose sheets and incubated respectively with each antibody reported above. Whereas no clear result was obtained with α-dystrobrevin, γ-sarcoglycan, and sarcospan, the specific antibodies presented in Figure 3 showed that (as expected) dystrophin (400-kD protein band), β-dystroglycan (43-kD protein band), and α-sarcoglycan (50-kD protein band) were detectable in both fast (W) and slow (R) muscles.
Fast muscle fibers from Torpedo contained α-syntrophin (59-kD) but appeared to be depleted of DRP2 and both β- and δ-sarcoglycan. There was also a 90-kD protein (called 90k) in crude protein extracts from white muscles, i.e., detected with monoclonal antibody 13E2 that we produced and which is assumed to correspond to the Ao protein reported by Yoshida and Ozawa (1990).
Comparative distribution at the muscle membrane of a few dystrophin complex-related proteins in the different muscle types analyzed
Slow muscle fibers from Torpedo were found to contain DRP2 (120-kD) and β-sarcoglycan (43 kD). δ-sarcoglycan appeared to be present but with an apparent migration of about 37 kD, as compared to 35–31 kD in mammalian muscles. No α-syntrophin or 90-kD protein was detected in Western blots.
In addition to these positive or negative detections, it was also possible to estimate the relative amount of proteins present in both muscle types. We have presented here a preliminary attempt at Western blotting quantification. The densitometric measurements enabled us to estimate that the relative OD levels were different in red (slow) and white (fast) Torpedo muscle. This corresponded to the presence of dystrophin (125%), β-dystroglycan (110%), and α-sarcoglycan (160%) at higher levels in Torpedo slow-type fibers.
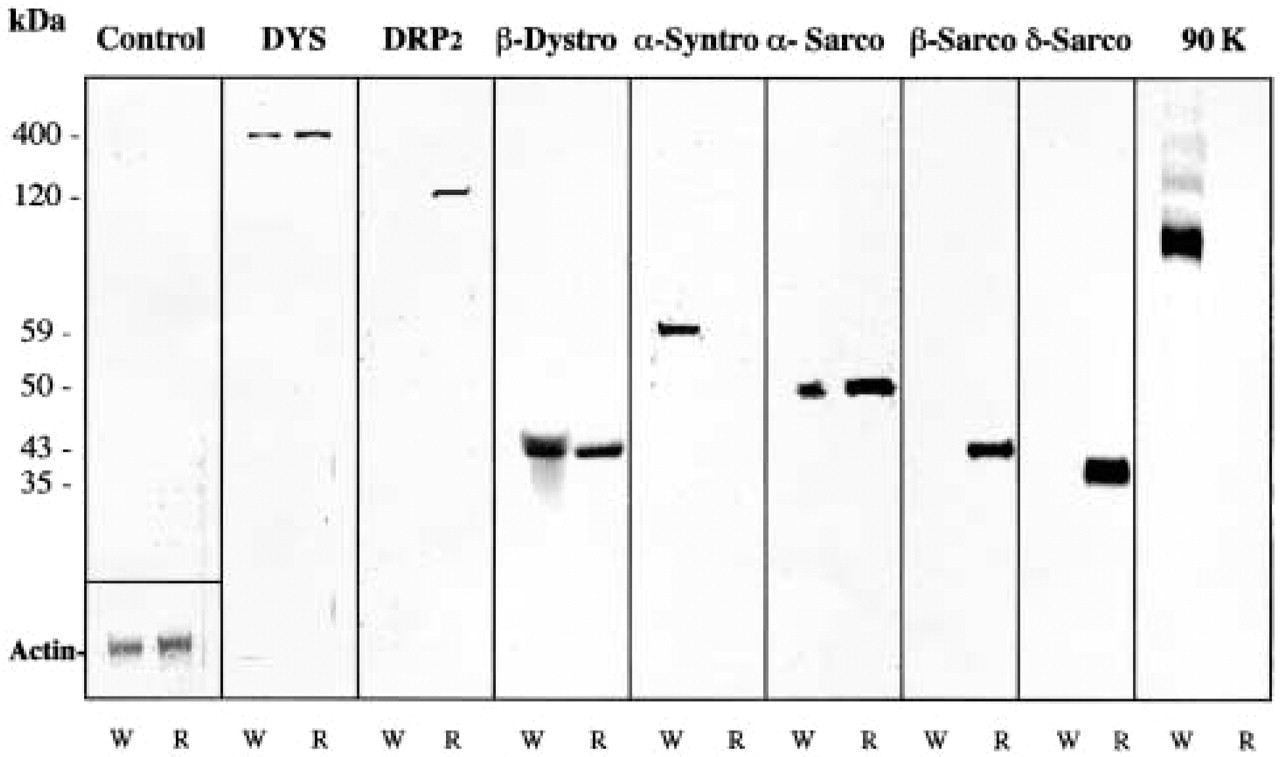
Western blot analysis of crude extracts from different T. marmorata muscles. Only monoclonal antibodies against dystrophin (5A3) and the 90-kD protein (13E2), and polyclonal antibodies that we produced were used in these Western blotting analyses. Crude protein extracts from W (white) fast-type fibers and R (red) slow-type fibers were analyzed by incubation with the primary antibody indicated at the top of the corresponding lanes. DYS, dystrophin; DRP2, dystrophin-related protein Type 2; β-dystro, β-dystrophin; α-syntro, α-syntrophin; α-sarco, α sarcoglycan; β-sarco, β-sarcoglycan; δ-sarco, δ-sarcoglycan; 90K, protein with Mr of 90 kD assume to correspond to Ao; C, control, in which the first specific antibody was omitted. Molecular weights are indicated at left. In the control lane, separation of the same extracts on SDS-PAGE stained with Coomassie brillant blue enabled estimation of the actin content.
Immunoprecipitation
Comparative analyses of proteins specifically immunoprecipitated with anti-β-dystroglycan polyclonal antibody and then retained on a protein A–Sepharose column are presented in Figure 4. The use of anti-β-dystroglycan revealed in both enriched membraneeluted fractions from white and red Torpedo muscles the presence of the expected proteins, such as β-dystroglycan, but also dystrophin. Similarly, α-sarcoglycan was detected in both membrane fractions, whereas DRP2 and δ-sarcoglycan proteins were found only in red muscle extracts. In contrast, only the white muscle-eluted fraction contained α-syntrophin.
Discussion
We comparatively investigated the composition of dystrophin-associated proteins from red and white caudal muscles in Torpedo marmorata. This was undertaken first to visualize the presence of this protein system at the fish muscle membrane and then to compare the expression pattern according to the muscle fiber type analyzed. We intended to establish, in the light of increasing knowledge on the number of molecules constituting dystrophin-associated protein complex, a catalogue of dystrophin family proteins that exist in slow- and fast-type muscle fibers from fish (which are similar in terms of myosin presence as tested with a monoclonal antibody directed specifically against slow fiber type muscles from mammals and which gave cytoplasmic staining only in red Torpedo muscle; not shown).
Western blotting analysis of immunoprecipitation fractions from the enriched membrane muscle preparation. Enriched membrane extracts from W (white) and R (red) Torpedo muscles were incubated with anti-β-dystroglycan. After immunoprecipitation with anti-β-dystroglycan and elution on a protein A–Sepharose column, the resulting fractions (eluted by 10% SDS) were analyzed by Western blotting. We revealed in these aliquots the presence or absence of some component of the DGC by treating the corresponding nitrocellulose sheets with specific antibodies indicated at left: β-dystro, β-dystrophin; Dys, dystrophin; DRP2, dystrophin-related protein Type 2; α-sarco, α-sarcoglycan; δ-sarco, δ-sarcoglycan; α-syntro, α-syntrophin.
With the monoclonal and polyclonal antibodies against dystrophin and/or utrophin used in this study, we identified two proteins in white and red muscles of Torpedo: a 400-kD protein band corresponding to Torpedo dystrophin and utrophin proteins and a 116-kD protein band that corresponded to a short Torpedo dystrophin/utrophin family product (referred to as DRP2). These proteins have already been described in the T. marmorata electric organ (Cartaud et al. 1993; Fabbrizio et al. 1993; Rivier et al. 1996) confined to the innervated surface of the electroplate (Chang et al. 1989; Yeadon et al. 1991). Different authors' studies of dystrophin-associated proteins in the electric organ of T. marmorata revealed some discrepancies regarding mammalian descriptions. Torpedo syntrophin was identified in the electric organ as a single protein highly homologous to α-syntrophin (Wagner and Huganir 1994). In this study, we described the α-syntrophin only in Torpedo white muscle. Dystroglycan in Torpedo electric organ consists of 190-kD and 43-kD proteins (Cartaud et al. 1993). In white and red muscles, with the polyclonal antibodies that we produced, β-dystroglycan was detected with a molecular weight of 43 kD, i.e., similar to the molecule described in mammalian muscle. In Torpedo electric organ, two sarcoglycans of 45 and 30 kD have been described that may correspond to β- and γ-sarcoglycan (Cartaud et al. 1993; Fabbrizio et al. 1997). Our Western blotting results showed that α-sarcoglycan was detected in both muscles studied, but β- and δ-sarcoglycan appeared only in red muscle. Finally, the 90-kD protein that was present in electric organ, in accordance with Yoshida and Ozawa (1990), was detected only in white muscle cell membranes.
These differences between Torpedo electric organ and two kinds of Torpedo muscles might be explained by the fact that Torpedo electric organ is embryonically derived from immature striated muscle (Fox and Richardson 1978). However, we report marked differences between white (fast-type fibers) and red (slow-type fibers) skeletal muscles. A summary of these comparative distributions is presented in Table 1. These two muscles are located in the caudal part of the fish but have different functions. In the past 10 years, previous studies partially revealed that cell and fiber distributions are interesting to study with respect to dystrophin (Howard et al. 1999). Studies on fast- and slow-twitch mammalian skeletal muscles (Anh et al. 1992) have revealed that dystrophin content is approximately twofold higher in soleus muscle (a slow-type muscle) than in vastus lateralis (a fast-type muscle). Quantitative analysis of some partners of the dystrophin complex, such as dystrophin, β-dystroglycan, and γ-sarcoglycan, increased by about 130%, 110%, and 120% in rat soleus muscle (Chopard et al. 2000). Our preliminary results are in close agreement with the above quantitative data corresponding to dystrophin and β-dystroglycan overexpression in slow-type muscles. γ-Sarcoglycan was not quantified in this work, but we observed significant overexpression (160%) for α-sarcoglycan in slow-type muscles. Recently, Straub et al. (1999) hypothesized that the novel є-sarcoglycan might replace α-sarcoglycan in smooth muscle but participate in the skeletal muscle dystrophin complex, as reported recently by Hack et al. (2000). This point might be interesting to investigate in slow- and fast-type muscle fibers from Torpedo in relationship to data obtained on α-sarcoglycan abundance in slow-type fibers.
Rafael et al. (2000), working with the dystrophin/utrophin-deficient dko mouse, obtained another interesting result. They found that the structural and biochemical properties of skeletal muscle (i.e., fiber type determination) were clearly under the influence of dystrophin and utrophin. In mammalian muscle, dystrophin and associated proteins link the extracellular matrix and the subsarcolemmal actin cytoskeleton, and deficiencies or variations in these proteins can generate different muscular pathologies, such as limb–girdle muscular dystrophies, that correspond mainly to sarcoglycan deficiency (Bushby 1999). The difference in the composition of the dystrophin-associated proteins complex could be related to the different structural and function properties in two muscle types. In Torpedo fish, we found significant differences between the two muscles. Red muscle is made up of short but abundant fibers that confer high strength to this muscle. White muscle has long fibers forming a propulsion organ that, by strong undulatory movements, plays a major role in fish locomotion. In red muscle, the dystrophin, β-dystroglycan, α-sarcoglycan complex, including DRP2, β-sarcoglycan, δ-sarcoglycan, γ-sarcoglycan with sarcospan and α-dystrobrevin, might be related to the high-strength capacity of this muscle and the absent α-syntrophin might be replaced by other proteins of the syntrophin family (β or γ isoform). In white muscle, α-syntrophin in complex with dystrophin/utrophin, α-sarcoglycan, and γ-sarcoglycan might be important for the fish undulatory movement, while the absence of DRP2, β-sarcoglycan, δ-sarcoglycan, and α-dystrobrevin might be compensated for the presence of a 90-kD protein and/or other proteins from the dystrobrevin family. Then, in reference to the work cited above (Rafael et al. 2000), we assumed that the presence of DRP2 in fish muscle might influence the fiber type.
To our knowledge, the present study is a rather broad investigation on dystrophin family products present in two different muscle types from the Torpedo caudal region. Due to the loss of the vasoconstrictor response regulation and eventual muscle damage that now appears to be correlated with dystrophin deficiency (Sanders et al. 2000; Crosbie 2001), it should be noted that a well-known difference between muscles is that red muscles are richer in capillary beds compared to white muscles (Romunal 1965). This study highlighted the interdependence between the dystrophin family system composition and the muscle type contractile performances (in connection with vascular control), which will be of interest for further study.
Footnotes
Acknowledgements
Supported in part by grants from INSERM and the Association Française contre les Myopathies (AFM). RM was a Consejo Social University of Alcalá and INSERM postdoctoral fellow over the course of this work.
We are very grateful to Nicole Lautrédou–Audouy for granting access to the confocal microscope at the Centre de Recherche en Imagerie Cellulaire (CRIC, Montpellier, France). We are also grateful to the fisherman Jacques Lapeyre, who provided us with the live fish.