
Abstract
The LIM domain is a protein-protein interaction motif critically involved in a variety of fundamental biological processes, including cytoskeletal organization, cell lineage specification, and organ development. In this study we examined the expression of the LIM proteins paxillin and Hic-5 in adult human tissues by immunohistochemistry and immunoblotting. Paxillin expression was widespread and observed both in non-muscle and muscle tissues. Of the latter, paxillin was mainly expressed in multinuclear striated muscle. In contrast, Hic-5 showed restricted expression and was expressed in muscle tissues, mainly in mononuclear smooth muscle. Taken together with previous findings, it appears likely that the counterbalance between paxillin and Hic-5 may be deeply involved in muscle differentiation.
Keywords
P
Hic-5 was originally identified as a transforming growth factor (TGF)β1- and hydrogen peroxide-inducible gene by differential hybridization (Shibanuma et al. 1994). Although Hic-5 was suggested to be involved in the negative regulation of cell growth, including the senescence process and TGFβ signal transduction (Shibanuma et al. 1994; Zhang et al. 2000), recent studies have shown that Hic-5 is a cytoskeletal protein localized to focal adhesions (Fujita et al. 1998; Matsuya et al. 1998). In addition, Hic-5 contains four LIM domains in the C-terminal half and four LD motifs in the N-terminal half, indicating that Hic-5 and paxillin share structural homology and hence functional similarities (Shibanuma et al. 1997; Thomas et al. 1999). In fact, several paxillin-interacting proteins that play important roles in integrin-mediated signal transduction and remodeling of the actin cytoskeleton, including FAK, vinculin, and syndesmos, are shared with Hic-5 (Thomas et al. 1999; Nishiya et al. 2001; Denhez et al. 2002).
Although it is now established that Hic-5 is a paxillin homologue localized to focal adhesion complexes, recent studies have shown that these LIM proteins have some distinct functional features (see Discussion). Furthermore, recent reports, including ours, have clearly shown that paxillin and Hic-5 may play distinct and exclusive roles in megakaryocytes and platelets. Paxillin, but not Hic-5, is expressed in megakaryocytes, and may play an important role in signaling pathways within the focal adhesions through interaction with Pyk2 (Hiregowdara et al. 1997; Hagmann et al. 1998). In contrast, Hic-5, but not paxillin, is expressed in platelets, is incorporated into the cytoskeleton on stimulation, and plays an important role in platelet aggregation and adhesion, in a manner dependent on the integrin αIIbβ3-mediated outside-in signaling (Hagmann et al. 1998; Oda et al. 2001; Osada et al. 2001). Accordingly, it is likely that a dramatic expression switch within the LIM proteins, i.e., from paxillin to Hic-5, occurs during megakaryocytic differentiation (into platelets). However, in other tissues little is known about the detailed expression patterns of paxillin and Hic-5. In this study we examined the expression of paxillin and Hic-5 in adult human tissues by immunohistochemistry and immunoblotting to provide an insight into the distinct functional roles of these LIM proteins.
Materials and Methods
Tissues
Cardiac muscle and iliopsoas muscle were obtained from adult autopsy cases. All other tissue samples were obtained from the surgical margins of various lesions. The sample collection was in accordance with the Yamanashi Medical University Guidelines. The tissues were immediately fixed in methanol for 24–28 hr and then embedded in paraffin.
Antibodies
In this study, two anti-LIM protein antibodies (Transduction Laboratories; Lexington, KY) were used. Anti-paxillin MAb (clone 349) recognizes not only the 68-kD paxillin protein but also the 50-kD Hic-5 protein, while anti-Hic-5 MAb (clone 34) recognizes only Hic-5 (Osada et al. 2001). We found that the anti-paxillin MAb (clone 165) specifically recognized paxillin in immunoblotting, but did not in immunohistochemistry. This antibody was likely to detect only denatured ones. Accordingly, clone 349 was used for detection of paxillin in this study.
Immunohistochemistry
Methanol-fixed, paraffin-embedded specimens were incubated with anti-paxillin MAb or anti-Hic-5 MAb (as the primary antibody) overnight at 4C. Binding of the primary antibody was detected using biotinylated anti-mouse IgG (ICN Pharmaceuticals; Aurora, OH) and streptavidin-peroxidase conjugate (Dako LSAB Kit; Dako Japan, Kyoto, Japan), and visualized with diaminobenzidine. The specimens were counterstained with Mayer's hematoxylin.
Immunoblotting
Fresh samples were incubated with ice-cold lysis buffer [final concentrations: 1% Triton X-100, 0.5% Nonidet P-40, 50 mM Tris-HCl (pH 7.2), 1 mM EGTA, 1 mM Na3VO4, 0.5 mM PMSF, and 50 μg/ml of leupeptin] and homogenized. The lysates were centrifuged at 15,000 X g for 10 minutes (at 4C). The resultant supernatants were resolved on 8% SDS-PAGE and then electrophoretically transferred to PVDF membranes. The membranes were blocked with 1% BSA in PBS. After extensive washing with PBS containing 0.1% Tween-80, the immunoblots were incubated with 0.05 μg/ml of anti-paxillin MAb or anti-Hic-5 MAb for 2 hr at room temperature. Antibody binding was detected using peroxidase-conjugated goat anti-mouse IgG and visualized with ECL chemiluminescence reaction reagents (Amersham Pharmacia Biotech; Poole, UK).
Results
The results are summarized in Table 1. For each result described in this table, sections from at least three tissue blocks were studied. The immunohistochemistry studies on different samples were highly consistent, with no major variations in the tissue distribution of anti-paxillin and anti-Hic-5 reactivities.
Reactivities of anti-paxillin or anti-Hic-5 antibody in normal human tissues
Widespread Expression of Paxillin and Restricted Expression of Hic-5
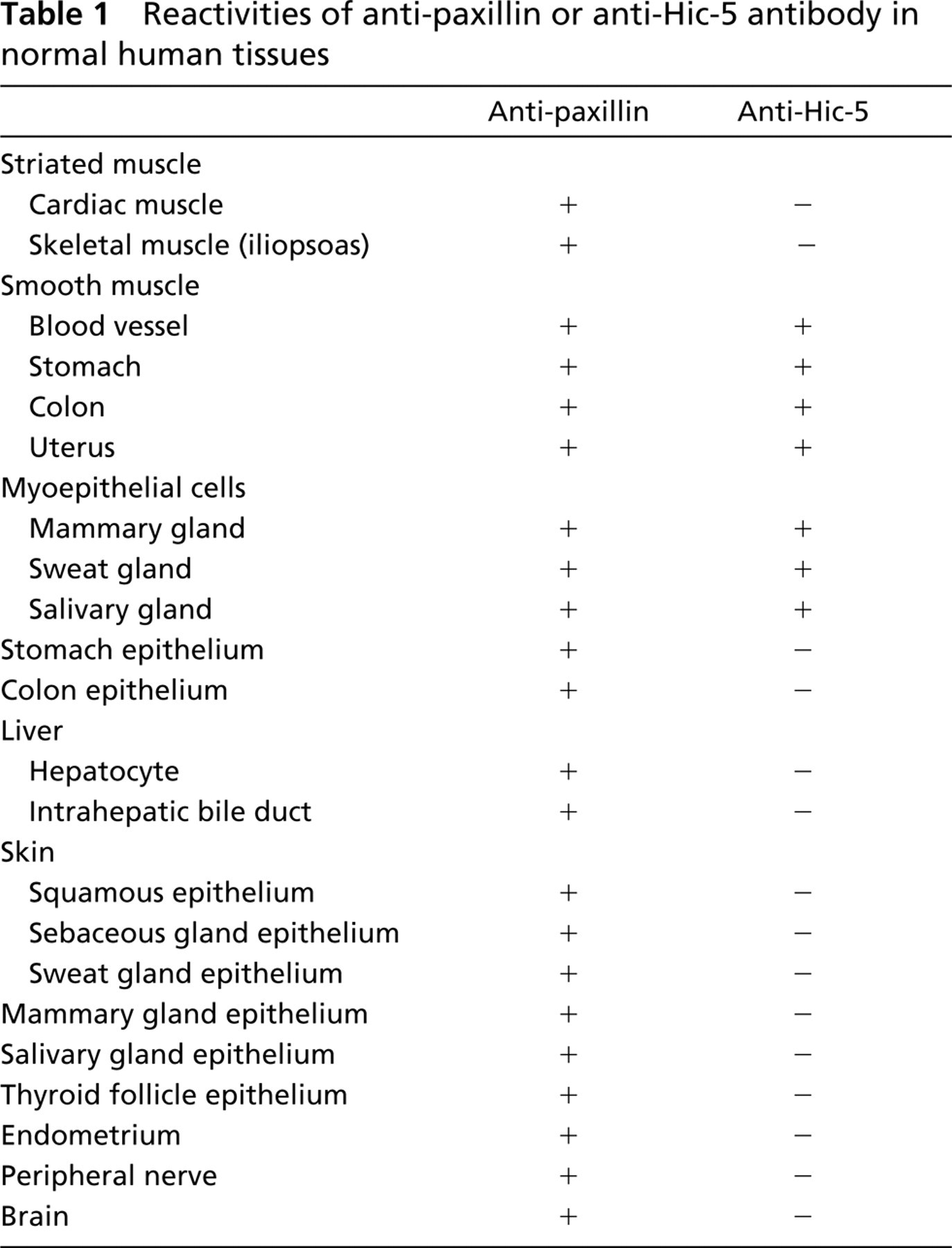
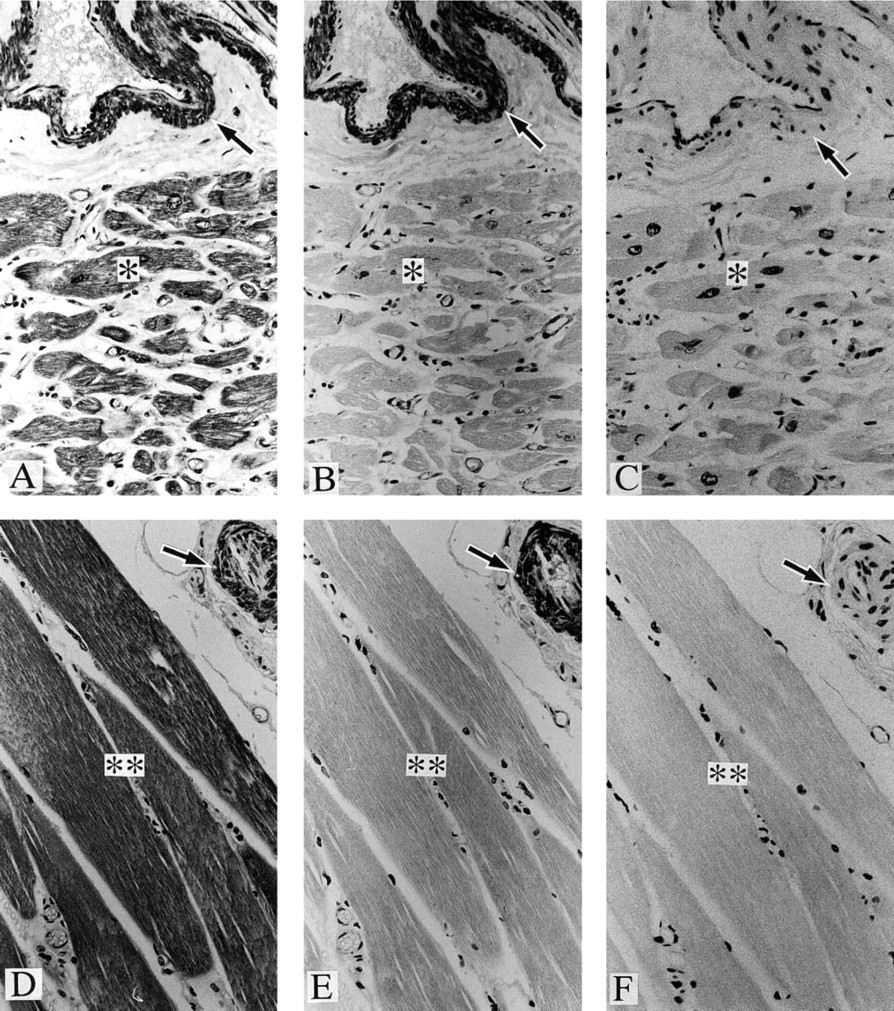
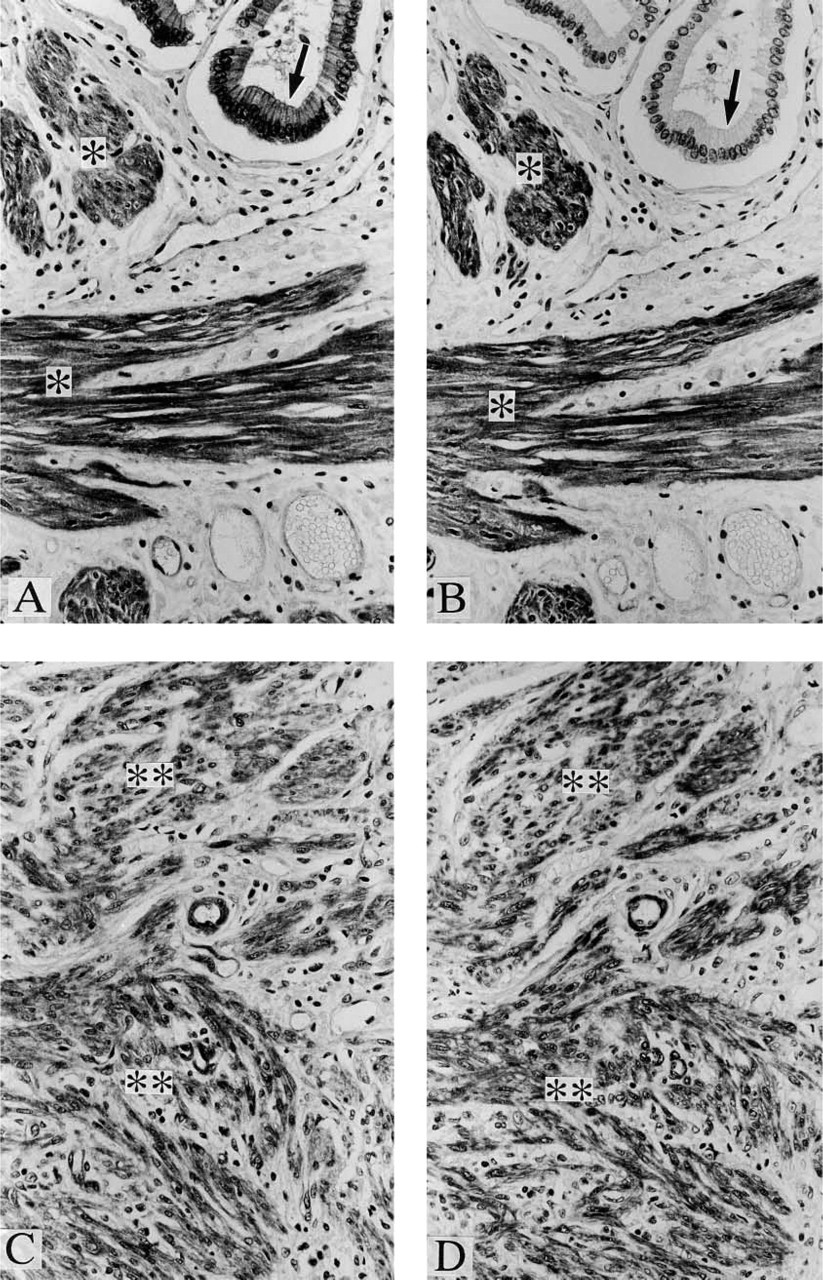
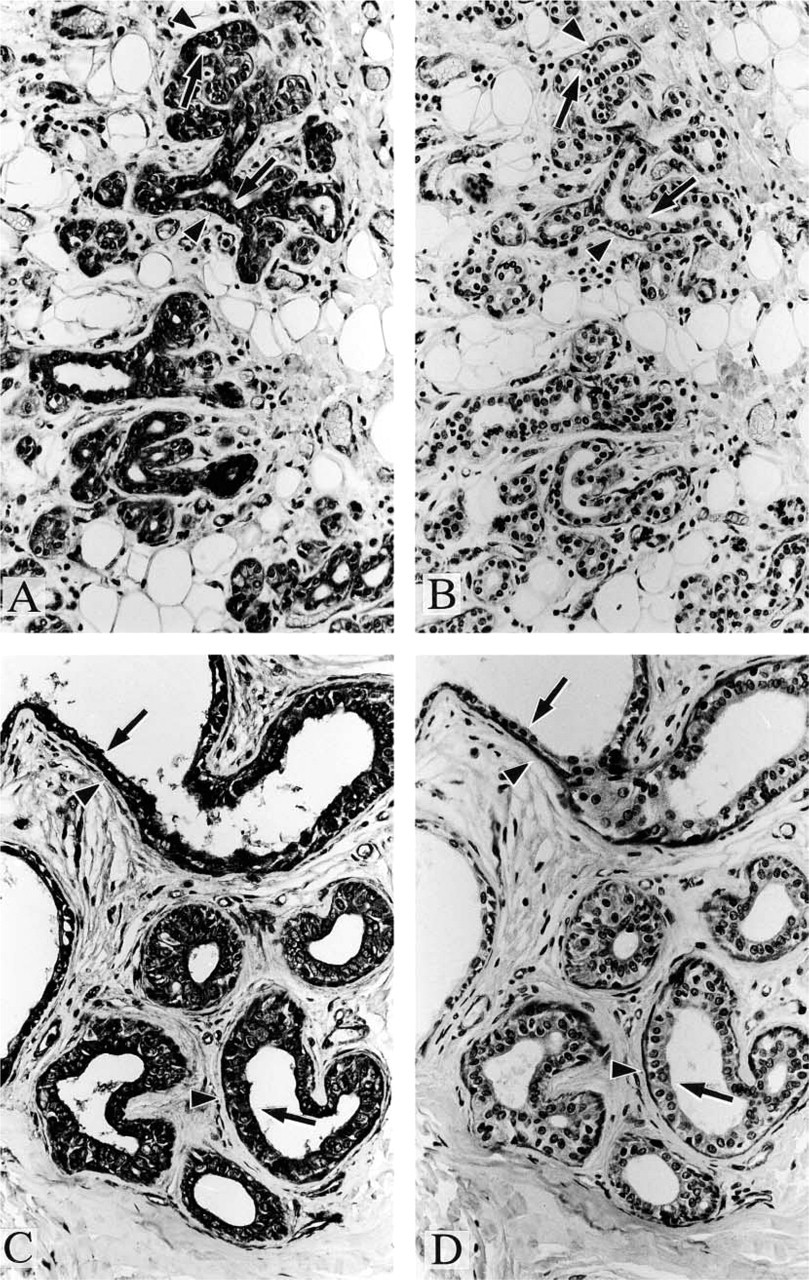
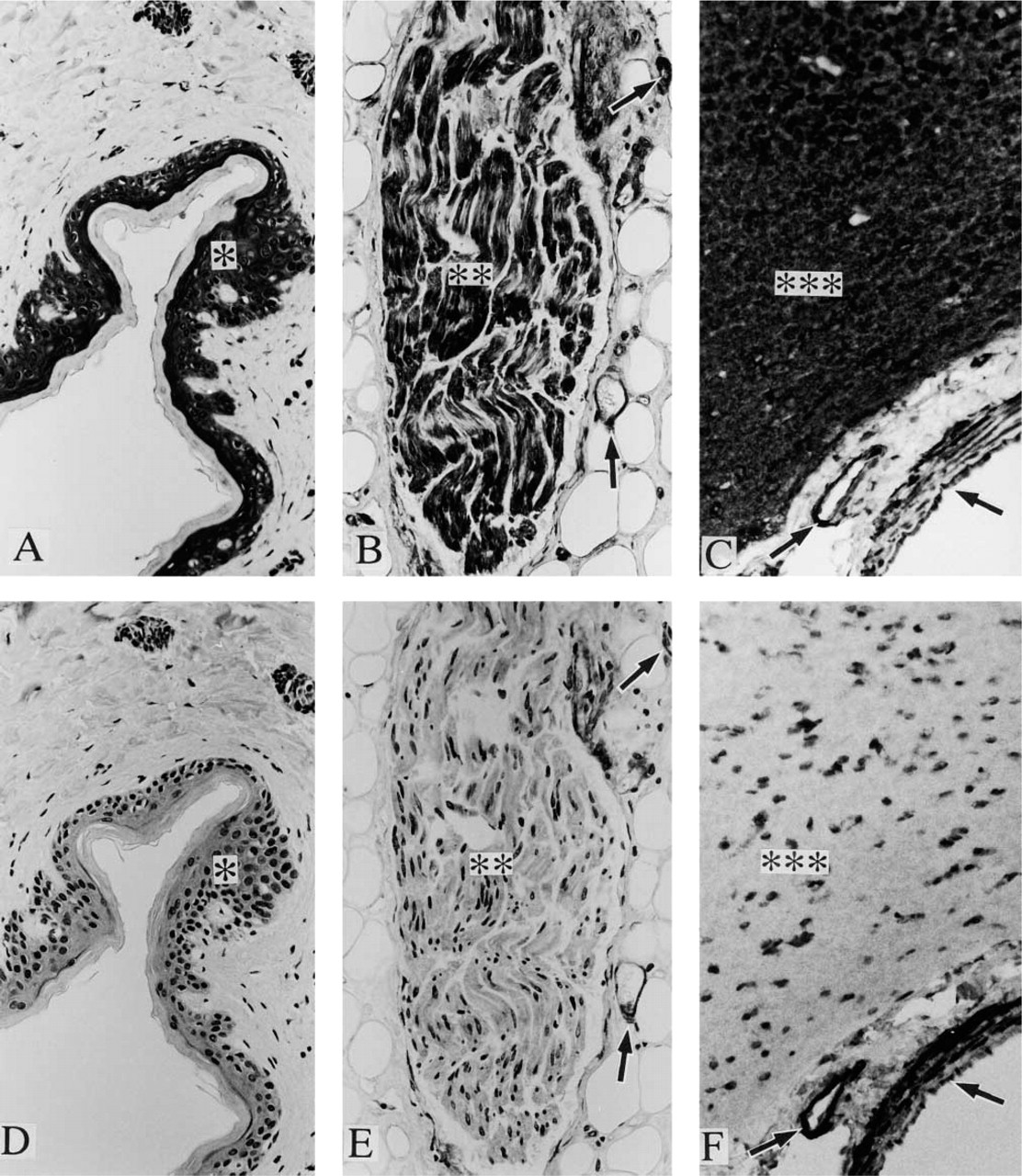
Positive staining was observed with anti-paxillin antibody in a variety of samples, including cardiac muscle (Figure 1A), skeletal muscle (Figure 1D), vascular smooth muscle (Figures 1A and 1D), organ smooth muscle (Figures 2A and 2C), gland epithelium (Figures 2A, 3A, and 3C), skin epithelium (Figure 4A), peripheral nerve (Figure 4B), and brain (Figure 4C); see Table 1 for summary.
Immunoreactivities with anti-paxillin MAb (
Immunoreactivities with anti-paxillin MAb (
Immunoreactivities with anti-paxillin MAb (
Immunoreactivities with anti-paxillin MAb (
In contrast, positive staining with anti-Hic-5 antibody was observed specifically in smooth muscle samples (Figures 1B, 1E, 2B, and 2D); see Table 1 for summary. As described in Materials and Methods, anti-paxillin MAb (clone 349) recognizes both paxillin and Hic-5, whereas anti-Hic-5 MAb recognizes only the latter. Accordingly, a combination of positive staining with anti-paxillin antibody and negative staining with anti-Hic-5 antibody indicates the expression of paxillin (but not Hic-5), which is true of striated muscle, epithelium (from various origins), and nerves (Table 1). On the other hand, whether paxillin or Hic-5 is predominantly expressed cannot be determined in the case of positive staining with both antipaxillin antibody and anti-Hic-5 antibody, which is true of smooth muscle (and myoepithelial cell) samples (Table 1).
Predominant Expression of Paxillin and Hic-5 in Striated Muscle and Smooth Muscle, Respectively
As described above, the relative expression of paxillin and Hic-5 in smooth muscle (and myoepithelial cell) samples cannot be determined solely by immunohistochemistry studies. Accordingly, immunoblotting studies were performed; paxillin and Hic-5 can be discriminated by the difference between their molecular weights. As shown in Figure 5, not only the 68-kD paxillin protein but also the 50-kD Hic-5 protein was confirmed to be recognized by immunoblotting with anti-paxillin MAb. Only the 50-kD Hic-5 protein was recognized by immunoblotting with anti-Hic-5 MAb. Muscle samples were analyzed by this procedure.
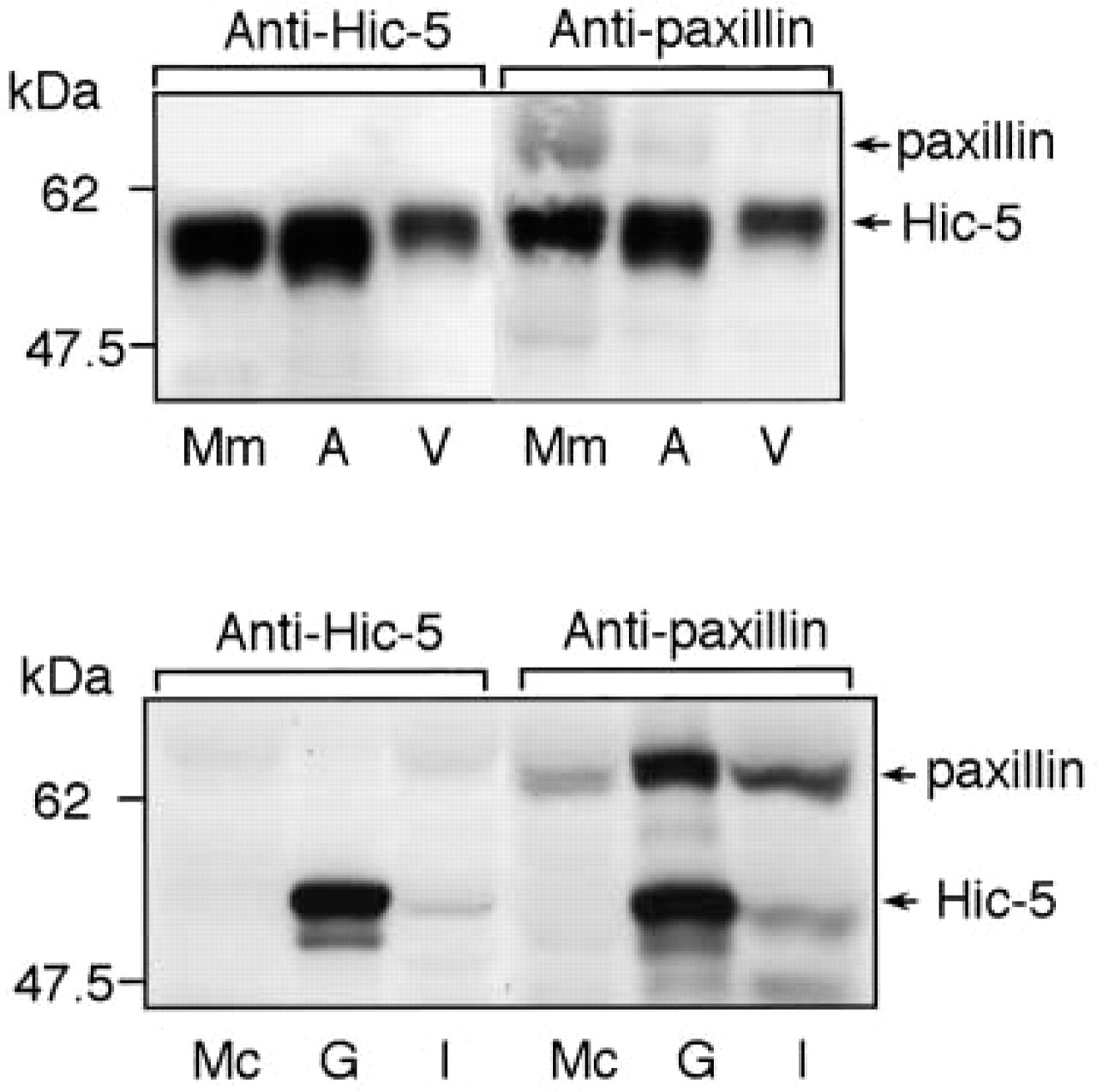
Abundant expression of paxillin and Hic-5 in the striated muscle (cardiac muscle and iliopsoas) and the smooth muscle (uterus, blood vessel, and stomach), respectively. The lysates obtained from the myometrium (Mm), gastric greater omentum artery (A), gastric greater omentum vein (V), myocardium (Mc), gastric smooth muscle (G), and iliopsoas (I) were resolved on SDS-PAGE and then immunoblotted with anti-paxillin or anti-Hic-5 MAb. Protein amounts loaded were not adjusted. Arrows indicate the locations of paxillin and Hic-5. The locations of molecular mass markers are indicated at left.
As expected from the immunohistochemistry studies, striated muscle samples, i.e., cardiac muscle and skeletal muscle (iliopsoas), were confirmed to express mainly paxillin (Figure 5). Hic-5 was not detected in cardiac muscle (Figure 5). Although this LIM protein was detected in iliopsoas, its expression was very weak (Figure 5). This weak Hic-5 detection may be due to cross-contamination from the vasculature, which abundantly expresses Hic-5. In contrast, smooth muscle samples, including myometrium, arterial and venous vascular smooth muscle tissues, and gastric smooth muscle, were found to strongly express Hic-5 (Figure 5). Paxillin expression was much weaker than Hic-5 expression in gastric and uterine smooth muscle cell (Figure 5). Furthermore, paxillin detection was almost negligible in vascular smooth muscle tissues. Paxillin was detected only very weakly in the artery and not at all in the vein (Figure 5).
Possible Expression of Hic-5 in Myoepithelial Cells
When mammary (Figures 3A and 3B), sweat, and salivary (Figures 3C and 3D) glands were studied by immunohistochemistry, positive staining with both anti-paxillin antibody and anti-Hic-5 antibody was observed in myoepithelial cells, which surround ductal and acinar epithelium of these glandular organs. It appears likely that myoepithelial cells express mainly Hic-5 because these cells are strongly stained with anti-Hic-5 antibody and because myoepithelial cells are of a smooth muscle phenotype, which is related to the predominant expression of Hic-5 (not paxillin). However, further studies are needed to clearly determine the relative expression of paxillin and Hic-5 in these cells. It was impossible to perform immunoblotting studies with the use of myoepithelial cells due to the difficulty in isolating them purely.
Discussion
Proteins containing LIM domains play important roles in a variety of fundamental biological processes, including cytoskeletal organization, cell lineage specification, and organ development. The LIM domain is a protein-protein interaction motif that is critically involved in these processes (Bach 2000). In this study we showed that the expression patterns of the LIM proteins paxillin and Hic-5 were very different. Paxillin expression was observed both in muscle and non-muscle tissues and was widespread, which is consistent with a previous finding that paxillin is present in a number of tissues in the chicken (including gizzard smooth muscle, cardiac muscle, skeletal muscle, liver, and brain) (Turner 1991). In contrast, Hic-5 shows restricted expression, at least when adult human tissues were examined by immunohistochemistry and immunoblotting. Hic-5 was expressed only in muscle tissues. Moreover, this paxillin homologue was expressed mainly in smooth muscle. The predominant expression of Hic-5 in myoepithelial cells may be explained by the fact that the myoepithelial cell is of a smooth muscle phenotype. Furthermore, Hic-5 is abundantly expressed in highly differentiated platelets, which may be related to the similarity in cytoskeletal components between smooth muscle cells and platelets (Nosaka et al. 1984; Takeuchi 1985). The ubiquitous expression of paxillin (compared with Hic-5) may be consistent with a report that paxillin seemingly plays an important role as an adapter and/or signaling protein in integrin-mediated adhesion and the resultant cytoskeletal responses, even in Drosophila, whereas Hic-5 was not found in the Drosophila genome (Wheeler and Hynes 2001). It seems likely that, from an evolutionary viewpoint, paxillin occurred first and Hic-5 appeared later to supplement or regulate the function of paxillin. Leupaxin is a novel LIM protein that is most homologous to paxillin (Lipsky et al. 1998). In fact, anti-paxillin MAb (clone 349) was found to recognize leupaxin when blood mononuclear cells were analyzed by immunoblotting. However, leupaxin was not detected in normal human tissues listed in Table 1 (data not shown), which is consistent with the previous report that this new paxillin homologue is preferentially expressed in hematopoietic cells (Lipsky et al. 1998).
Although it is established that Hic-5 is a paxillin homologue localized to focal adhesion complexes (see Introduction), recent studies have shown that these LIM proteins have some distinct functional features. Hic-5 expression is increased during cell senescence of normal human fibroblasts and is decreased during immortalization of mouse embryo fibroblasts (Shibanuma et al. 1994; Ishino et al. 2000). The forced expression of Hic-5 in immortalized fibroblasts induced growth retardation and senescence cell-like morphology (Shibanuma et al. 1997). Accordingly, Hic-5 is involved in the negative regulation of cell growth, including the senescence process and TGFβ signal transduction, which is not the case with paxillin. Paxillin tended to increase when the Hic-5 expression level was specifically decreased during immortalization (Ishino et al. 2000). Furthermore, it was reported that Hic-5 inhibits cell spreading via competition with paxillin for FAK and subsequent prevention of downstream signal transduction. The expression of anti-sense Hic-5 was shown to increase spreading (Nishiya et al. 2001). It was hypothesized that Hic-5 could compete for common interaction factors with paxillin and antagonize the signaling pathways that involve paxillin. This competitive effect between paxillin and Hic-5 through interaction with FAK suggests that the counterbalance of paxillin and Hic-5 expression may be a novel mechanism regulating integrin-mediated signal transduction and the resultant cytoskeletal reorganization (Nishiya et al. 2001). These opposing functions of paxillin and Hic-5 may be related to the distinct expression patterns of these LIM proteins.
The contrasting expression of the LIM proteins in muscle tissues may be worthy of notice. Paxillin is mainly expressed in multinuclear striated muscle, whereas Hic-5 is in mononuclear smooth muscle, at least in adult human tissues. Recently, these LIM proteins have been shown to be involved in muscle differentiation. During muscle development, myoblasts function as muscle stem cells and, in response to the loss of trophic support, exit the cell cycle, fuse, and up-regulate a variety of genes that commit them to form striated skeletal muscle. Those cells that fail to activate appropriate survival and differentiation programs instead rapidly die by apoptosis (Hu et al. 1999). It was shown that the expression of Hic-5 blocks differentiation and induces apoptosis in mammalian myoblasts and that Hic-5 expression is specifically and dramatically induced in normal myoblasts that die after removal of trophic support (Hu et al. 1999). When myoblasts were placed in a differentiation medium and the Hic-5 levels were increased dramatically, immunocytochemical analysis of such cells revealed that the high levels of Hic-5 protein were restricted to mononucleated and apoptotic myoblasts relative to multinucleated myotubes (Hu et al. 1999). Therefore, Hic-5 appears to function as a regulator to block muscle differentiation and induce death (Hu et al. 1999), and differentiation into multinucleated striated cells may be related to reduction of Hic-5 levels. Furthermore, Hic-5 was recently shown to be strongly and transiently expressed first in the early developing heart and then in the smooth muscle layer of the developing tissues, including the intestinal tract and bronchial airways, in mice (Brunskill et al. 2001). These results suggest that Hic-5 expression changes in a dynamic pattern in embryonic development and is involved in muscle differentiation. However, it was also shown that paxillin overexpression prevents myoblast differentiation (Sastry et al. 1999). Although it is likely that the LIM proteins Hic-5 and paxillin are deeply involved in muscle differentiation, available knowledge on these LIM proteins in muscle development is still limited and fragmentary. Further studies on the expression of and mutual interactions among paxillin, Hic-5, integrins, and other cytoskeletal proteins are awaited.
Footnotes
Acknowledgements
Supported in part by a grant-in-aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology, Japan.
We are indebted to Dr T. Takafuta (Yamanashi Medical University) for helpful discussions.