
Abstract
As techniques evolve that allow molecular characterization of disease processes such as cancer, definition of “normal” at a molecular level becomes increasingly important. Increasingly large numbers of mutations are found at the genomic level, but whether all of those mutations contribute to the malignant state of a carcinoma cell is not clear. Without knowledge of what constitutes normality on the proteomic level in an organ or cell, we cannot determine what genomic changes are physiologically important. Traditionally, colon cancer is identified and classified by histological criteria. Margins of the colon are defined as “grossly uninvolved” when the histology is indistinguishable from that of normal (free from disease) colon. By using molecular pathology techniques and working backward from colon adenocarcinoma to hypoplastic polyps to presumably normal mucosa, we defined some of those protein differences. Our results may provide a molecular basis for identifying tumor formation and progression in situ.
(
T
Materials and Methods
Grossly uninvolved colon samples resected for reasons other than cancer, colon polyps, and matched samples of grossly uninvolved/adenocarcinoma from the same patient were screened in the pilot study (n = 5 for each group of samples). Tissue samples were collected from patients who had given informed consent before surgery at the University of Utah Hospital. The tissues were embedded in paraffin by standard techniques and sections were cut at 6-μm thickness for immunohistochemistry. The human gastric mucin (HGM), mucin-1, CA242, and heat shock protein 70 (HSP70) antibodies were from Novacastra/Vector Labs (Burlingame, CA). γ-Catenin/plakoglobin and zonula occludins-1 (ZO-1) antibodies were from Santa Cruz Biotechnology (Santa Cruz, CA). β-Catenin antibody was from Transduction Laboratories (San Diego, CA), and the topoisomerase IIα antibody was from DAKO (Carpinteria, CA). Antibodies conjugated to biotin (Jackson ImmunoResearch; West Grove, PA) were used to detect the primary antibodies, followed by use of the Vector Elite ABC–HRP kit according to the manufacturer's instructions. The reactions were visualized with Vector Lab's DAB substrate kit and the sections counterstained with Gill's hematoxylin (Vector Labs) before dehydration and mounting. Relative staining intensity and distribution (pattern change) were assessed by two independent observers (KM and KN).
Results
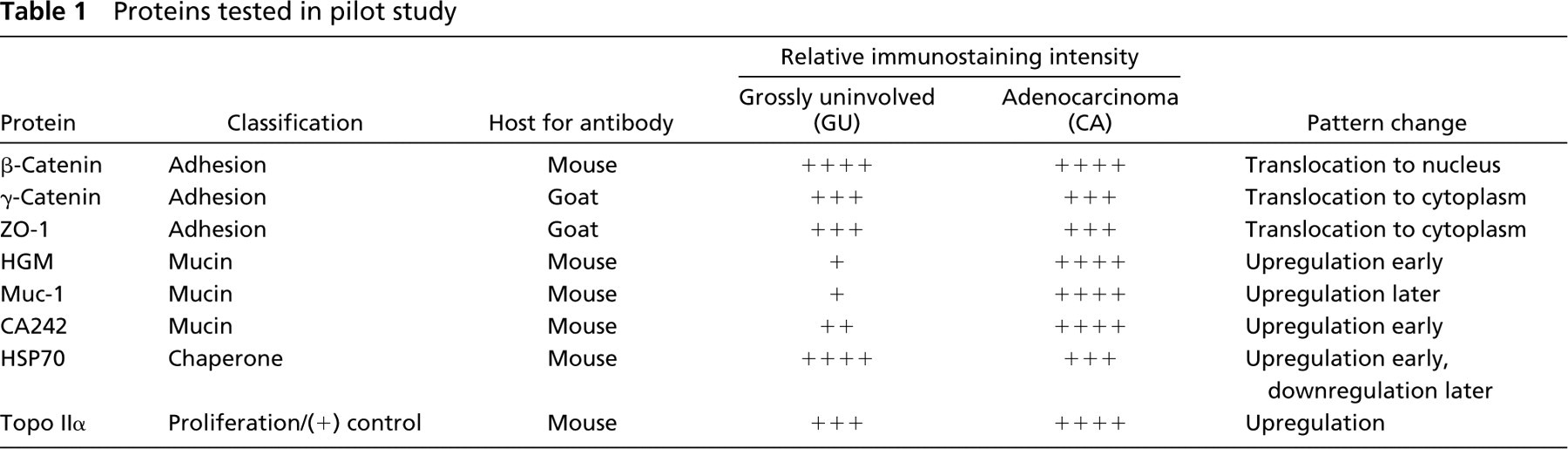
Baseline immunostaining intensity and distribution for each protein were determined by examining the grossly uninvolved (GU) samples. The proteins associated with adhesion tended to have the same relative immunostaining intensity between GU and the polyps or adenocarcinoma (CA) (Table 1), while their expression translocated from the cell membrane in the GU samples to either the cytoplasm of the cell or the nucleus in the polyp and CA samples. The mucins showed variable immunostaining patterns. None of them was highly expressed by either signal intensity or distribution in the GU samples. Muc-1 was expressed less in the polyps than in the CA, whereas HGM or CA242 proteins were expressed more in the polyps than in the CA. The HSP70 signal in GU colon epithelium was in the apical half of the crypt, whereas it was seen throughout the neoplastic epithelium. HSP70 was upregulated in the polyps and in the least dysplastic of the CAs, and was downregulated (as compared to the grossly uninvolved samples) in the most dysplastic cancers. Topoisomerase IIα was upregulated and generally more widespread in CA, as we expected.
Proteins tested in pilot study
Conclusions
Protein expression (relative intensity and distribution) differs between grossly uninvolved colon, polyps, and adenocarcinoma. Recently, a group found approximately 11,000 genomic mutations per colon carcinoma cell (Stoler 1999), although whether or not all of those mutations contributed to the malignant state of the cell remains to be determined. In what has been called the new biology (Quirke 1999), molecular techniques are available that distinguish abnormal clones of cells in morphologically normal mucosa. The role of β-catenin protein in the progression of colon cancer is increasingly well defined (Mann et al. 1999; Brabletz et al. 2000), and led us to investigate its family member, γ-catenin, and ZO-1, another cell-cell adhesion molecule, to determine if their protein expression changes during that progression. Although their expression patterns did not change as dramatically as that of β-catenin, other groups have found that they do play roles in pathways that may contribute to the development of the disease (Gottardi et al. 1996; Hakimelahi et al. 2000), and further study of these proteins may identify more subtle changes indicating that a given cell is moving from normal signaling and adherence to an abnormal state. Similarly, a group recently studied the distribution of four apomucins (Biemer–Huttmann et al. 1999) to determine their expression in normal epithelium and colon polyps. We compared the expression of Muc-1 with HGM and CA242 to see if we could identify differences in their expression patterns and found that they, along with HSP70, may be useful markers to determine progression of the cancer. A recent report demonstrated that inhibition of the synthesis of HSP70 in breast cancer cell lines activated a tumor cell-specific apoptosis that was independent of known caspases and p53 tumor suppressor protein and could not be rescued by Bcl-2 (Nylandsted et al. 2000). This, along with reports linking topo IIα immunostaining to either metastases (Holden 1999) or altered chemosensitivity (Jarvinen et al. 2000) indicate that these may be useful prognostic markers in colon cancer.
Footnotes
Acknowledgements
Acknowledgments
Supported by grants 5 P3 CA2014 and PO1 CA73992–03 from the National Institutes of Health and by the Huntsman Cancer Foundation.