
Abstract
Growth hormone (GH) exerts its immune effects on mature lymphocytes through an autocrine/paracrine mechanism. We investigated the prenatal synthesis of GH mRNA in rat lymphoid organs using the sensitive in situ RT-PCR methodology. We show that GH transcripts are detectable in the thymus and liver of the 18-day fetus. At this stage, all thymocytes are immature and express the GH gene. In fetal liver, GH gene expression was localized in circulating lymphocytes and in hematopoietic cells surrounding GH mRNA-negative hepatocytes. In situ GH gene expression in fetal lymphoid organs was confirmed by in vitro RT-PCR showing that the amplified product from fetal lymphoid tissues was similar to the product obtained from the pituitary. Moreover, GH gene expression was detected in the thymus, spleen, and ileum Peyer's patches of adult rat, with a localization restricted to the lymphocytes and endothelial and smooth muscle cells of blood vessels. The autocrine/paracrine expression of the GH gene by lymphoid and hematopoietic cells during fetal growth might influence the generation of regulatory cells involved in immunity and hematopoiesis.
P
It was assumed for decades that embryonic and fetal growth were independent of maternal or fetal GH. However, although pituitary GH synthesis starts on Day 19 of gestation (Rieutort 1974) and GH mRNA expression on Day 18 (Binder et al. 1994), the demonstration of GH receptor expression in rat embryos from Days 1–18 has suggested that the hormone might have a role during early stages of development (Garcia-Aragon et al. 1992). Moreover, the detection of GH mRNA at the morula stage (Day 3) as well as a GH-like immunoreactivity in blastocysts (Pantaleon et al. 1997) strongly indicated that the role of the hormone during fetal development must now be reconsidered. In rat fetus, liver hematopoiesis and thymus development occur at Day 12, whereas spleen differentiation begins to take place at Day 20 of gestation. The precise mechanisms regulating the immune system during fetal growth are still largely unknown. We therefore sought to determine whether fetal immune system cells could express the GH gene and then to visualize and identify immune GH-producing cells in rat fetus.
Materials and Methods
Tissue Preparation
Rat tissues were obtained from male Wistar rats (2 months) housed at 22C on a 12-hr light/dark cycle and 18-day-old (E18) fetuses (Iffa Credo; l'Arbresle, France). Food and water were available ad libitum. Whole fetuses were obtained after cesarean section and fetal tissues by microsurgery. For RNA isolation, adult and fetal rat tissues were stored at −80C until used. For in situ RT-PCR, adult male rat tissues and whole fetuses were fixed in 4% paraformaldehyde and paraffin-embedded. Sections 7 μm thick were mounted on in situ RT-PCR glass slides.
Oligonucleotide Primers and Probes
Two 25-mer oligonucleotide primers, HPLC-purified, were synthesized (Eurobio; Les Ulis, France) and used for the RT-PCR. The antisense primer (nt 135–160: 5′-CGCAAAGCGGCGACACTTCATGACC-3′) is located at the end of the fifth exon and the sense primer (nt 643–668: 5′-CCAGTCTGTTTGCCAATGCTGTGCT-3′) is in the second exon (Binder et al. 1994). For in situ hybridization, two 30-mer oligonucleotide probes located in the middle of the amplified fragment were synthesized. The sequence of the sense probe (nt 407–437) was 5′-AACAGCCTGATGTTTGGTACCTCGGACCGC-3′ and of the antisense probe (nt 248–278) was 5′-TGAGAAGCAGAACGCAGCCTGGGCATTCTG-3′. Probes were 3′ end-labeled with digoxigenin-11-dUTP (Roche Diagnostics; Meylan, France).
RT-PCR
Total RNA was extracted by the guanidium isothiocyanatephenol–chloroform method. One μg of total RNA was primed using polyT and M-MLV reverse transcriptase (Gibco BRL; Cergy Pontoise, France). RT reaction products were amplified by 30 cycles of PCR on a thermal cycler (Perkin) using 1 U/μl Taq DNA polymerase (Eurobio). The hot start method was employed and each cycle consisted of denaturating at 94C for 45 sec, annealing at 69C for 45 sec, and extension at 72C for 1 min, followed by 7 min at 72C. Amplified products were analyzed on a 1.5% agarose gel.
In Situ Hybridization
Dewaxed sections were treated with 5 μg/ml proteinase K (37C, 15 min) and hybridized overnight at 40C with 30 pmoles of antisense-labeled probe/ml of hybridization buffer as described (Mertani and Morel 1995). Sections were washed in 2 × and 1 × SSC at room temperature and digoxigenin-labeled probe was detected with alkaline phosphatase-conjugated anti-digoxigenin and visualized with NBT/BCIP chromogen.
In Situ RT-PCR
Dewaxed sections treated with proteinase K were dehydrated and air-dried. In situ RT reaction buffer (100 μl) contained 50 mM Tris-HCl, 75 mM KCl, 10 mM dithiothreitol, 3 mM MgCl2, 1 mM of each deoxynucleotide triphosphate, 1 μM antisense primer, 10 U RNase inhibitor, and 200 U M-MLV. The sections were covered with 40 μl of the reaction buffer, sealed with amplicover discs and amplicover clips, and incubated at 42C for 1 hr. Coverclips were removed and sections were washed in 0.1 M phosphate buffer, dehydrated, and air-dried. In situ PCR reaction buffer (100 μl) contained 10 mM Tris-HCl, 50 mM KCl, 3 mM MgCl2, 0.5 mM of each deoxynucleotide triphosphate, 1 μM of each primer, and 20 U Taq DNA polymerase. The sections were covered with 40 μl of this reaction buffer, sealed, and placed on a Perkin-Elmer thermal cycler. A total of 25 PCR cycles were performed under the same conditions as for liquid PCR. Sections were then fixed with 4% paraformaldehyde (15 min) and washed as described in the ISH procedure. The sections were then hybridized overnight at 40C with 30 pmoles of sense- and antisense-labeled probes/ml after 3-min denaturation at 94C. Washing steps were done as described for the ISH procedure. Several controls were performed: (a) ISH only, (b) omission of the reverse transcriptase, and (c) omission of the Taq DNA polymerase.
Results
Expression of GH mRNA in Rat Lymphoid Tissues by Liquid RT-PCR Analysis
The high expression of GH gene in pituitary was used as a positive control (Figure 1, Lane 2). The predicted band of 533 bp was found in adult spleen (Figure 1, Lane 3), thymus (Figure 1, Lane 4), ileum Peyer's patches (Figure 1, Lane 5), fetal thymus (Figure 1, Lane 6), and fetal liver (Figure 1, Lane 7). No band was detected in adult liver tissue (Figure 1, Lane 8) or when the Taq polymerase was omitted (Figure 1, Lane 9). We confirmed the identity of the PCR product after Rsa I digestion, giving two predicted bands of 290 and 243 bp (data not shown).
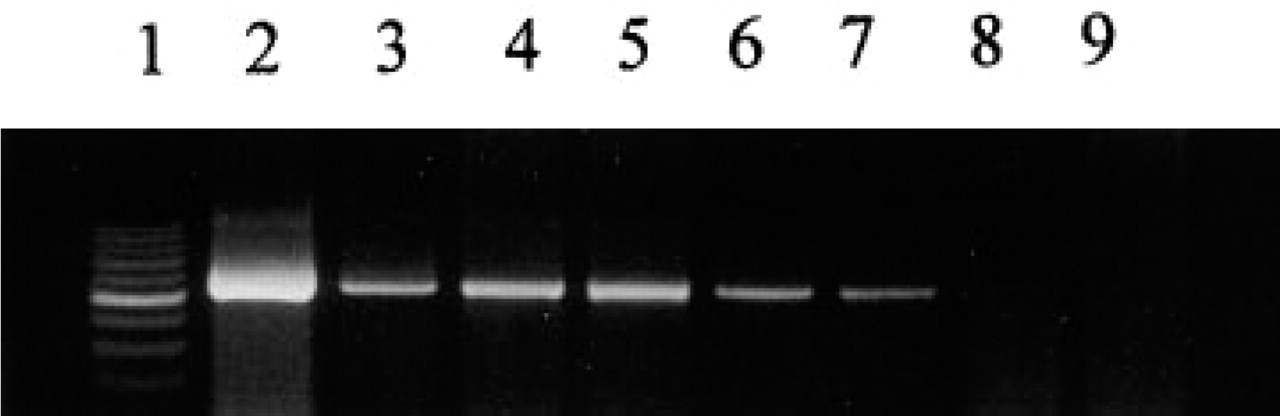
RT-PCR amplification of GH mRNA derived from adult pituitary (Lane 2), spleen (Lane 3), thymus (Lane 4), Peyer's patches (Lane 5), fetal thymus (Lane 6), and fetal liver (Lane 7). Negative controls included RT-PCR amplification of adult liver RNA (Lane 8) and RT-PCR performed without Taq polymerase on RNA extracted from adult spleen (Lane 9).
Localization of GH gene expression on adult rat pituitary (
Localization of GH mRNA by ISH and In Situ RT-PCR
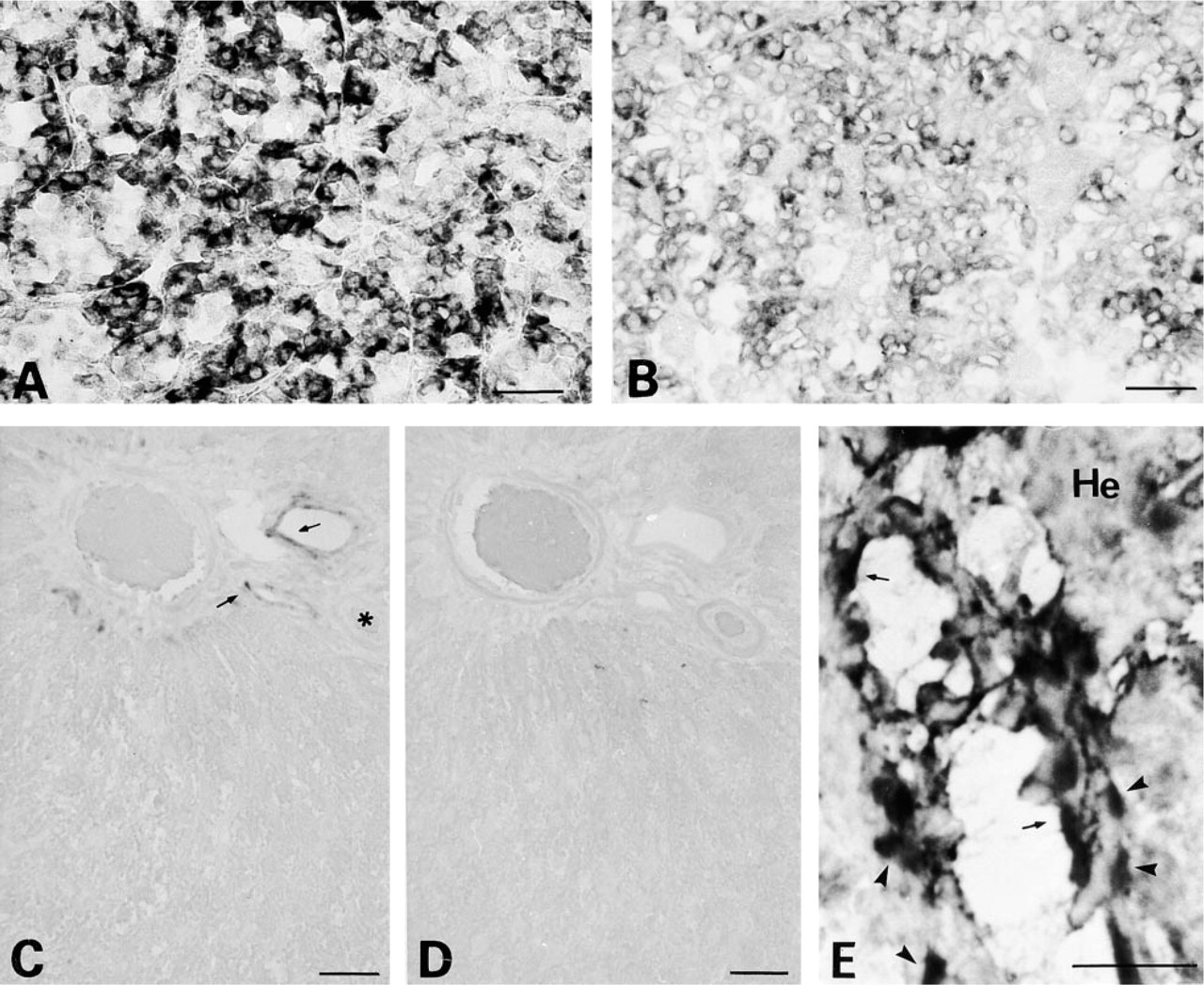
The positive signal indicating the expression of GH gene appears as a dark purple precipitate (Figures 2–4). The in situ RT-PCR methodology was validated on pituitary sections. The signal obtained (Figure 2A) was stronger compared to the ISH signal (Figure 2B) and was restricted to the somatotrophs, representing about 40% of the pituitary cells (Dada et al. 1984). In the rat liver, a non-GH-secreting organ, no signal could be evidenced in hepatocytes either by in situ RT-PCR (Figure 2C) or ISH (Figure 2D). However, we found that both endothelial and smooth muscle cells of blood vessel wall of portal spaces synthesize GH mRNA (Figures 2C and 2E) In the thymus of adult rats, using in situ RT-PCR, GH mRNA-positive cells were mainly localized in the thymic cortex, the site of T-cell differentiation (Figure 3A). A strong signal was also associated with the endothelial and smooth muscle cells of the vessel wall (arteries and veins) (Figure 3A). In the medulla, no signal could be detected in reticular or epithelial cells (Figures 3A and 3B). GH gene expression could be efficiently detected by ISH in the adult thymus (Figure 3C) although the signal appeared weaker compared to that obtained after in situ RT-PCR (Figure 3B). In the spleen of adult rat, GH mRNA synthesis was localized in the lymphoïd white pulp that surrounds and follows the arteries, and in the lymphocytes of the red pulp (Figure 3D). A GH-positive signal was also associated with the vessel walls, particularly those of the vascular sinuses of the red pulp (Figure 3D). These results were validated by the absence of any signal when the Taq polymerase was omitted during the PCR reaction (Figure 3E). In the Peyer's patches of the terminal ileum of the adult rat, all lymphocytes showed a positive signal for GH mRNA, whereas epithelial intestinal cells remained negative (Figure 3F). No positive signal could be detected when the reverse transcriptase was omitted in the reaction (Figure 3G), thus demonstrating the validity of the in situ RT-PCR methodology used.
Localization of GH gene expression by in situ RT-PCR on adult rat sections of thymus (
At Day 18 of gestation, the majority of GH mRNA-positive cells in the thymus were localized at the periphery of thymic lobes in the fetal lymphocytes, whereas cells in the center exhibited a weak in situ RT-PCR signal (Figure 4A). No signal was detected when the Taq polymerase was omitted in the PCR reaction (Figure 4B). In the fetal liver, strong expression of the GH gene was detected in hematopoietic cells that surrounded large GH mRNA-negative hepatocytes (Figures 4C and 4E). When direct ISH was used for detection of GH gene expression, the positive signal could be detected only within the lymphocytes circulating in the fetal liver sinusoidal spaces (Figure 4D). Finally, we also found that lymphocytes from the intracardiac cavity express the GH gene, in contrast to red blood cells (Figure 4F).
Discussion
In this study, GH gene expression was characterized in fetal and adult rat lymphoid tissues, using RT-PCR and the highly sensitive in situ RT-PCR. The primers used guaranteed the amplification of almost the entire GH transcript. Both sense and antisense primers of GH span two exons, and intronic regions between the primers are approximately 1100 bp. Therefore, amplification of genomic DNA was impossible. The PCR product of 533 bp identified in immune organs is of GH gene origin and is similar to the pituitary product. We identified lymphocytes as the major GH mRNA-positive cells in lymphoid organs of the fetal and adult rats. In the adult thymus, the site of T-lymphocyte differentiation, GH mRNA synthesis was localized mainly in cells of the cortex, although some cells of septa and medulla were GH mRNA-positive. Lymphocytes expressing the GH gene were identified according to their localization in the cortex, where they are in the majority compared to the medulla. In our study, rat GH gene expression could not be detected in reticular or epithelial cells, in contrast to studies on human thymus (Maggiano et al. 1994; Wu et al. 1996). In the spleen, GH mRNA-positive cells were localized in the germinal centers containing a majority of B-lymphocytes. A similar signal localized in lymphocytes was also detected in rat Peyer's patches. This result contrasted with the in situ hybridization study by Delhase et al. (1993) showing that GH gene expression occurs in the red pulp and marginal zone of rat spleen. This difference could be due to the very high sensitivity of the in situ RT-PCR we have used. Since the studies by Weigent's group, it is known that murine lymphocytes produce GH in vitro, as well as in vivo (Weigent et al. 1988; Baxter et al. 1991; Weigent and Blalock 1991), and interactions between GH and the immune system are well documented (Clark 1997). Expression of the GH receptor gene was detected in vivo in the rat spleen and thymus (Mertani and Morel 1995) and in vitro in B- and T-lymphocytes, thymocytes, and circulating immune cells (Badolato et al. 1994; Rapaport et al. 1995), strongly suggesting that the hormone modulates the immune functions through an autocrine/paracrine mechanism. The paracrine effects of GH may not support body growth but they would affect cytokine production from the immune cells, as demonstrated by Weigent and Blalock (1994) using GH-secreting spleen cells transferred to dwarf mice. It was indeed demonstrated that lymphocyte-produced GH, structurally identical to pituitary GH, functions as an autocrine growth factor in lymphoproliferation (Weigent et al. 1991). Moreover, proliferation of human thymocytes in primary culture is dependent on their own GH secretion and resultant production of IGF-I (Sabharwal and Varma 1996). The existence of an autocrine regulatory circuit has also been confirmed in the human thymus with complete phenotypic characterization of GH and GH receptor-positive cells (de Mello–Coelho et al. 1998). Wu et al. (1996) also suggested that GH expression in thymoma and in T- and B-cell lymphoma might be involved in tumor progression. Of keen interest, we localized GH gene expression in endothelial and smooth muscle cells of veins and arteries of the liver, spleen, and thymus. Because the GH receptor is expressed by the rat endothelial vasculature (Lobie et al. 1992), it is likely that GH production by endothelial cells plays a direct role in angiogenesis. Indeed, hGH has been shown to stimulate endothelial cell proliferation in vitro (Rymaszewski et al. 1991), and work by Smith et al. (1997) clearly demonstrated that GH is a very potent stimulator of transgenic mice retinal neovascularization. In addition, a recent study has shown that endothelial vasodilatation mechanisms of rat aortic rings are directly stimulated by GH (Rossoni et al. 1999). Interestingly, Struman et al. (1999) have shown that one possible mechanism explaining the in vivo effects of hGH on formation of capillaries involved the activation of the MAPK signaling pathway. The physiological importance of such an autocrine loop might also be relevant in human development, because GH receptors expressed by vascular endothelial and smooth muscle cells are detectable by 8.5 weeks of fetal age (Simard et al. 1996). Finally, one could expect that the angiogenic potential of GH, which is secreted by diverse neoplasms, would be directly involved in tumor growth and metastasis.
Localization of GH gene expression by in situ RT-PCR on E18 fetal sections of thymus (
Our study has demonstrated for the first time that the GH gene is expressed by fetal lymphoid tissues. The fetal GH gene was expressed principally in thymic and blood lymphocytes. The demonstration of GH synthesis by fetal lymphocytes is of importance and suggests that the hormone could directly regulate immune and hematopoietic functions during fetal growth. Experiments that strongly suggest no role for GH in fetal development are disputed by the demonstration that GH receptors are expressed in major rat organs at fetal stages E18 (Garcia–Aragon et al. 1992), E12 (Edmonson et al. 1995), in mouse embryonic stem cells (Ohlsson et al. 1993), splenocytes from 16- and 18-day mouse fetuses (Gagnerault et al., 1996), and mouse preimplantation embryos (Pantaleon et al. 1997). Futhermore, GH gene expression at the morula stage E3 and in blastocysts (Pantaleon et al. 1997) clearly suggests that the hormone has a pleiotropic autocrine/paracrine action on embryonic growth. In this study, we also evidenced GH gene expression in hematopoietic progenitor cells of the fetal liver. In this environment, local production of GH might regulate growth and differentiation of hematopoietic cells as do other cytokines sharing an identical signal transduction pathway. Tian et al. (1998) demonstrated that hGH is a potent stimulator of hematopoeisis after bone marrow transplantation, in doses that do not promote anabolic growth effects. Indeed, if endocrine GH has been shown to exert hematopoietic growth-promoting effects (Kelley et al. 1996), the impact of autocrine/paracrine GH on the common progenitors shared by hematopoietic and endothelial cell lineages is still unknown and represents a challenge for future studies.